Microbiome Differences in Preeclampsia Versus Lupus Nephritis
Harshita Nadella, Julia L Armstrong, Rachel Greenwald, Kenneth Johnson, Marc M Kesselman

TL;DR
This paper compares the microbiomes of preeclampsia and lupus nephritis, highlighting similarities and differences that could aid diagnosis and treatment.
Contribution
The study identifies disease-specific microbiome patterns in preeclampsia and lupus nephritis that could be used for differentiation and targeted therapies.
Findings
Both preeclampsia and lupus nephritis show reduced microbiome diversity and pro-inflammatory microbes.
Preeclampsia is linked to microbes causing endothelial dysfunction, while lupus nephritis involves gut dysbiosis promoting renal inflammation.
Shared microbiome features suggest potential for microbiome-based diagnostics and therapies.
Abstract
Preeclampsia (PE) and lupus nephritis (LN) share clinical features of hypertension, proteinuria, and systemic inflammation, reflecting overlapping immune dysregulation. Both conditions involve activation of pro-inflammatory cytokines and endothelial dysfunction, which contribute to organ damage. They also exhibit similarities in their microbiomes, including reduced diversity and loss of beneficial immunoregulatory taxa, which may exacerbate systemic inflammation. Despite similarities, they differ in etiology. PE results from placental dysfunction, whereas LN arises from autoimmune-driven renal injury. PE is associated with enrichment of pro-inflammatory microbes, which contribute to endothelial dysfunction and impaired trophoblast invasion. In contrast, LN exhibits gut dysbiosis involving expansion of pro-inflammatory species and depletion of protective immunoregulatory taxa, promoting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
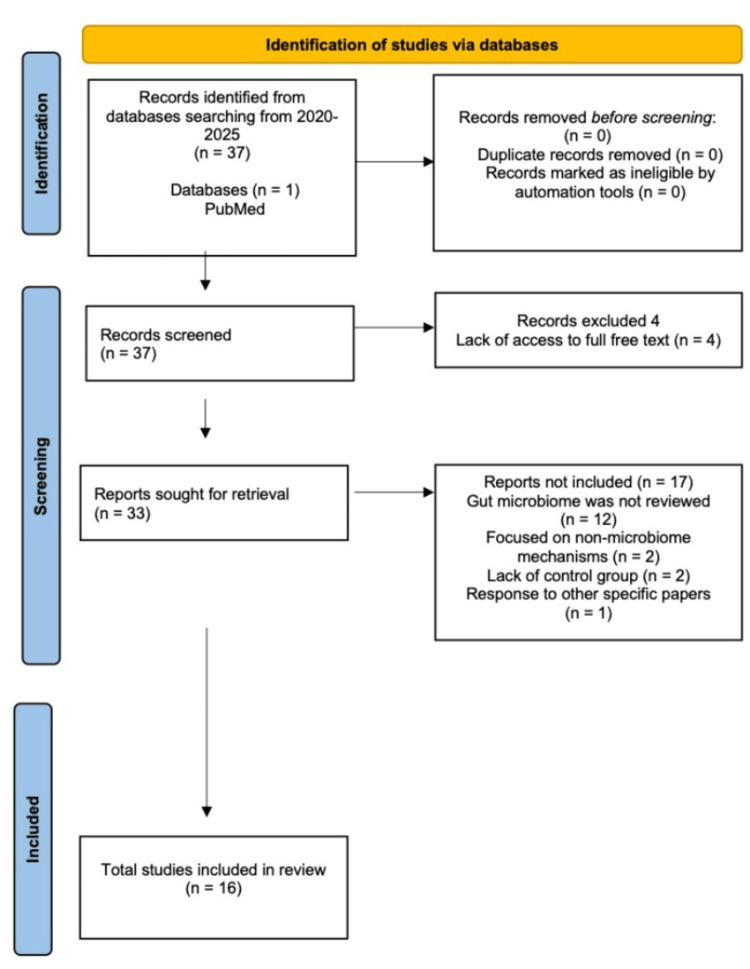 Figure 1
Figure 1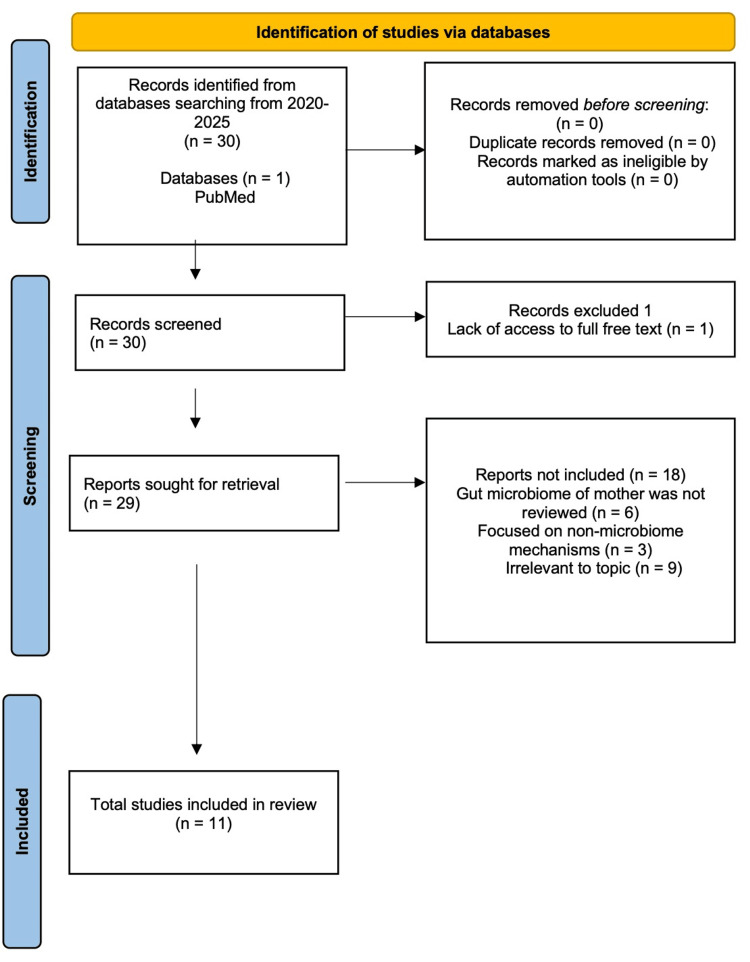 Figure 2
Figure 2| Diagnosis | Diagnostic criteria | Management |
| Chronic hypertension | HTN diagnosed before pregnancy or in the first 20 weeks of pregnancy | Consider delivery at 37 weeks; lifestyle modifications; consider antihypertensives if BP ≥140/90 |
| Gestational hypertension | New-onset HTN ≥140/90 after 20 weeks; no prior HTN; asymptomatic | Consider delivery at 37 weeks; lifestyle modifications; consider antihypertensives if BP is consistently ≥140/90 |
| Preeclampsia (without severe features) | HTN (≥140/90) and proteinuria diagnosed by: 24-hour urine collection >300 mg/24 hours; urine protein/creatinine ratio >0.3; urine dipstick >2+ protein | Consider delivery at 37 weeks; lifestyle modifications; consider antihypertensives if BP is consistently ≥140/90; educate on warning signs and symptoms of preeclampsia |
| Preeclampsia (with severe features) | Gestational HTN plus ≥1: severe HTN (≥160/110); thrombocytopenia (<100,000); impaired renal function (serum Cr >1.1 or 2× baseline); elevated AST/ALT (>2× ULN) or severe RUQ/epigastric pain; pulmonary edema; new onset of headache unresponsive to medication or visual disturbances (blurred vision, scotoma) | Consider delivery at 34 weeks if mother and fetus are stable; earlier if maternal/fetal distress; consider termination if pregnancy is prior to fetal viability; lifestyle modifications; start antihypertensives if BP is consistently ≥140/90; magnesium sulfate for seizure prophylaxis; monitor BP, O2, and urine output; continuous fetal monitoring, US, and appropriate antenatal testing (e.g., biophysical profile) |
| HELLP syndrome | Preeclampsia plus all: Hemolysis (↓ Hb, ↓ haptoglobin, ↑ LDH, ↑ indirect bilirubin); elevated liver enzymes (AST/ALT >2× ULN); low platelets <100,000 | Start antihypertensives if BP is consistently ≥140/90; magnesium sulfate for seizure prophylaxis; monitor BP, O2, and urine output; continuous fetal monitoring; consult hematology; administer blood products if needed; continuous fetal monitoring and deliver immediately |
| Eclampsia | New-onset seizures in a patient with preeclampsia | Start continuous fetal monitoring and immediate delivery; treat seizures; airway/oxygen support; magnesium sulfate (add additional medications if needed); monitor BP, O2, and urine output; start antihypertensives if BP is consistently ≥140/90 |
| Organ | Pathogenesis |
| Kidney | Glomerular endothelial dysfunction and hypertension-induced vasoconstriction lead to proteinuria, impaired renal function, and edema |
| Lung | Increased SVR and volume overload lead to left ventricular dysfunction, increased pulmonary capillary hydrostatic pressure, increased capillary permeability, and decreased albumin; this leads to pulmonary edema and respiratory distress |
| Liver | Vasoconstriction and microthrombotic obstruction lead to liver cell damage and liver swelling |
| CNS | Vasoconstriction and endothelial damage lead to vasospasm in the CNS, which can lead to seizures |
| Blood | Systemic microthrombi and vasoconstriction lead to overactivation of the coagulation system and platelet consumption, which leads to DIC, thrombocytopenia, and anemia |
| Class | Histopathology |
| Class 1 | Minimal mesangial lupus nephritis |
| Class 2 | Mesangial proliferative lupus nephritis |
| Class 3 | Focal lupus nephritis (<50% of glomeruli involved) |
| Class 4 | Diffuse lupus nephritis (≥50% of glomeruli involved) |
| Class 5 | Membranous lupus nephritis |
| Class 6 | Advanced sclerosing lupus nephritis |
| Author (year) | Experimental group | Control group/Baseline | Microbiome source | Detection methods | Micro-organisms increased | Micro-organisms decreased |
|
Gao et al. (2025) [ | MRL/lpr mice at 10, 15, and 20 weeks | MRL/lpr mice at 4 weeks | Fecal pellets | 16S rDNA sequencing; metagenome sequencing |
Phylum: |
Phylum: Firmicutes; Genus: |
|
Silverman et al. (2022) [ |
C57BL/6 GF mice colonized with Lupus-derived | C57BL/6 GF mice colonized with healthy RG strain (RG1/ATCC29149) | Fecal samples (lupus nephritis patients S47/S107 for experimental strains; healthy donor for control strain) | Bacterial culture (BHI media plates); 16S rRNA-specific PCR amplification; whole-genome sequencing; RG-specific qPCR |
Species: | N/A |
|
Li et al. (2020) [ |
MRL/lpr mice treated with | PBS-treated MRL/lpr mice (vehicle group); C57BL/6J mice (normal control) | Gut |
N/A (no profiling; | N/A | N/A |
|
Pan et al. (2025) [ | MRL/lpr mice: MSC (hUC-MSC at weeks 6, 8, 10); MSC-FMT (MSC + FMT at weeks 9–13) | MRL/lpr mice treated with saline | Fecal samples from 15-week-old mice | Metagenomic sequencing |
Genus: |
Genus: |
|
Cabana-Puig et al. (2022) [ | In-house colony MRL/lpr mice | New MRL/lpr mice from JAX | Fecal samples (16-week-old female mice) | 16S rRNA sequencing |
Phylum: |
Family: |
| Author (year) | Study design | Groups (number of subjects) | Microbiome source | Detection methods | Micro-organisms increased | Micro-organisms decreased |
|
Azzouz et al. (2023) [ | Observational | SLE patients with LN flares (n = 16); HC (n = 22) | Gut (fecal) | 16S rRNA gene sequencing; shotgun metagenomic sequencing |
Species: | N/A |
|
Cheng et al. (2025) [ | Observational | SLE patients (n = 36: 18 SLE–non-LN, 18 SLE–LN); HC (n = 15) | Gut (fecal) | 16S rRNA gene sequencing; untargeted metabolomics |
Phylum: Proteobacteria ; Family: |
Phylum: Firmicutes; Genus: |
|
Mohd et al. (2023) [ | Narrative review | N/A (summarizes human and mouse studies) | Gut (fecal) - from cited studies | 16S rRNA sequencing (reported from cited studies) |
Genus: | N/A |
|
Wang et al. (2023) [ | Systematic review | SLE-LN patients (n = 138); SLE patients (n = 441); HC (n = 1526); animal studies (n = 5) | Gut (fecal) | 16S rRNA gene sequencing; metagenomic sequencing; PCR |
Phylum: |
Phylum: |
|
Parodi et al. (2025) [ | Narrative review | N/A | Gut (fecal) - from cited sources | 16S rRNA sequencing; metagenomic sequencing (from cited sources) |
Phylum: | Phylum: F/B ratio decreased |
|
Tan et al. (2024) [ | Narrative review | N/A | Gut (fecal) | 16S rRNA sequencing (from cited sources) |
Species: | N/A |
|
Gui et al. (2024) [ | Observational and experimental | SLE patients (n = 114: 62 non-LN, 52 LN); HC (n = 134); MRL/lpr mice | Gut (fecal) | 16S rRNA gene sequencing; shotgun metagenomic sequencing |
Genus: | N/A |
|
Wu et al. (2025) [ | Observational and experimental | SLE patients (n = 61: 11 non-LN, 26 active LN, 24 LN in remission); HC (n = 25); MRL/lpr mice (n = 20); MPJ mice (n = 5) | Gut (fecal); mouse spleen/MLN | 16S rRNA sequencing; flow cytometry; ELISA |
Phylum: |
Genus: |
|
Lau et al. (2021) [ | Narrative review | N/A | Gut (fecal) | 16S rRNA sequencing (from cited sources) |
Genus: |
Phylum: |
|
Wang et al. (2022) [ | Observational | SLE patients (n = 19); HC (n = 19) | Gut (fecal) | 16S rRNA sequencing |
Genus: |
Genus: |
|
Yu et al. (2022) [ | Observational | LN patients (n = 15); HC (n = 27) | Gut (fecal) | 16S rRNA sequencing |
Phylum: |
Phylum: |
| Author (year) | Study design | Groups (number of subjects) | Microbiome source | Detection methods | Micro-organisms increased | Micro-organisms decreased |
|
Chen et al.(2020) [ | Case-control | Human: PE (n = 67), NP (n = 85) Mouse PE-FMT (n = 8), NP-FMT (n = 10), controls (n = 8) | Gut (fecal) | 16S rRNA gene sequencing; qPCR in situ hybridization |
Genus: |
Genus: |
|
Jin et al. (2022) [ | Case-control | Human: PE (n = 35); healthy pregnant controls (n = 35) | Gut (fecal) | 16S rRNA gene sequencing |
Phylum: |
Phylum: |
|
Tian et al. (2024) [ | Case-control | EOPE (n = 48); LOPE (n = 32); HC (n = 96) | Gut (fecal) | 16S rRNA gene sequencing |
Species: |
Genus: |
|
Beckers et al. (2020) [ | Narrative review | N/A | Gut, placental, oral, vaginal | 16S rRNA gene sequencing; qPCR |
Phylum: |
Genus: |
|
Ishimwe, JA (2021) [ | Narrative review | N/A | Gut, placental, oral, vaginal | 16S rRNA gene sequencing |
Species: |
Phylum: |
|
Giugliano et al. (2025) [ | Case-control | PE (n = 30); normotensive controls (n = 25) | Gut (fecal) | 16S rRNA gene sequencing |
Order: |
Phylum: |
|
Lv et al. (2024) [ | Case-control | PE (n = 19); normotensive controls (n = 20) | Gut (fecal) | 16S rRNA sequencing; shotgun metagenomic sequencing |
Genus: |
Species: |
|
Han et al. (2025) [ | Case-control | PE (n = 34); LP control (n = 39) | Gut (fecal) | 16S rRNA gene sequencing |
Genus: |
Genus: |
|
Lin et al. (2020) [ | Case-control | Primary cohort: SPE (n = 30) vs controls (n = 30); Confirmation cohort: SPE (n = 580) vs controls (n = 55) | Vaginal | 16S rRNA gene sequencing; qPCR |
Species: | N/A |
|
Wu et al. (2024) [ | Case-control | HDP patients (n = 6); normotensive controls (n = 9) | Gut (fecal) | 16S rRNA gene sequencing; bioinformatics |
Species: |
Genus: |
|
Chang et al. (2020) [ | Case-control | Severe PE patients (n = 27); healthy controls (n = 36) | Gut (fecal) | 16S rRNA gene sequencing; gas chromatography for SCFAs |
Phylum: |
Species: |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPregnancy and preeclampsia studies · Systemic Lupus Erythematosus Research · Pregnancy and Medication Impact
Introduction and background
The purpose of this review is to identify whether microbiome profiles can distinguish preeclampsia (PE) from lupus nephritis (LN). To accomplish this, we compare microbiome differences between PE and LN across multiple body sites, including the gut, vaginal, and placental compartments. Drawing on high-quality literature published since 2020, we aim to identify distinct and shared microbiomes, explore how these relate to immune and vascular dysfunction, and evaluate their potential utility as diagnostic biomarkers, risk stratifiers, and therapeutic targets.
Hypertensive pregnancy disorders occur in 6-8% of pregnancies in the United States. PE is part of a spectrum of hypertensive pregnancy disorders ranging from chronic hypertension to HELLP (hemolysis, elevated liver enzymes, low platelet count) syndrome, each defined by specific criteria and management (Table 1). Metabolic diseases such as obesity and diabetes, cocaine use, advanced maternal age, chronic kidney disease (CKD), and nulliparity may lead to endothelial dysfunction, impaired placental implantation, and increased risk for the development of hypertensive pregnancy disorders [1]. PE is a complex, multisystem hypertensive disorder that arises after 20 weeks’ gestation, marked by new-onset hypertension (≥140/90 mmHg) and proteinuria or end-organ dysfunction [1]. Affecting approximately 5-7% of all pregnancies in the United States, PE remains a leading cause of maternal and perinatal morbidity and mortality, particularly in low- and middle-income women [1,2].
While historically considered a placental disorder, modern research highlights its multifactorial etiology, involving aberrant placentation, systemic endothelial dysfunction, chronic inflammation, and maladaptive maternal immune responses to the semi-allogeneic fetus [3,4]. At the center of PE pathogenesis is a failure of early trophoblast invasion and spiral artery remodeling, leading to placental hypoxia and ischemia [5]. This results in acute atherosis of the decidual vessels, characterized by arterial wall fibrinoid necrosis and lymphoid infiltration [5]. Due to the abnormal blood flow, hypertension with systemic vasoconstriction causes placental hypoperfusion. This hypoxic stress stimulates the release of antiangiogenic factors such as soluble fms-like tyrosine kinase-1 and soluble endoglin, which antagonize vascular endothelial growth factor and placental growth factor signaling, leading to maternal endothelial dysfunction and multisystem injury [5]. These pathological changes are further compounded by immune dysregulation, characterized by exaggerated natural killer cell activation, increased T-helper (Th)1/Th17 responses, and impaired regulatory T-cell function, all of which contribute to poor placental perfusion and systemic inflammation [6]. An increase in the placental factors of inflammation and dysfunction leads to endothelial lesions that result in microthrombosis [7]. There is an array of consequences from vasoconstriction and microthrombosis across a multitude of organs beyond the placenta, as illustrated in Table 2.
Systemic lupus erythematosus (SLE) is an autoimmune disease characterized by the production of autoantibodies and immune complex deposition. This leads to chronic systemic inflammation and multi-organ damage. Among its many manifestations, LN is one of the most serious and common complications, affecting up to 60% of patients with SLE at some point in their lives [8]. LN is a well-recognized cause of CKD and end-stage renal disease in young women, particularly among African Americans and Hispanics who exhibit higher prevalence and worse outcomes [9]. LN also disproportionately affects women of reproductive age and can both mimic or complicate the diagnosis of PE during pregnancy [9].
The pathogenesis of LN involves the formation of circulating immune complexes containing anti-double-stranded DNA (anti-dsDNA) antibodies and/or anti-Smith (anti-Sm) antibodies, which deposit in the mesangial or subendothelial regions of the glomerulus [8]. This triggers a local inflammatory response through complement activation and downstream cytokine release involving T cells, B cells, and kidney cells, ultimately resulting in varying degrees of glomerular injury, including thickening of the mesangium, capillary walls, and glomerular basement membrane [8]. This pathophysiology also shares several overlapping features with PE, including immune dysregulation, endothelial injury, and proteinuria [3,7]. The most common clinical manifestations of LN are hypertension, edema, and hematuria, which mirror those seen in PE. Diagnosis requires laboratory evaluation, including a complete blood count, basic metabolic panel (BMP), and urinalysis. The BMP typically reveals elevated creatinine, while the urinalysis may show proteinuria, hematuria, cellular casts, and a urine protein-to-creatinine ratio >0.5 [10]. A kidney biopsy is performed when indicated for the following reasons: an unexplained increase in creatinine, proteinuria >1.0 g/day, or proteinuria >0.5 g/day with hematuria or cellular urinary casts [10]. The biopsy findings demonstrate immune complex-mediated glomerulonephritis [10].
Histologically, LN is classified by the International Society of Nephrology/Renal Pathology Society into six distinct classes, ranging from minimal mesangial involvement to severe proliferative or sclerotic changes (Table 3). All patients with LN should be evaluated by a nephrologist, especially those in class III or IV [10]. These two classes typically require both induction and maintenance therapy. Induction therapy consists of intravenous (IV) glucocorticoids, such as methylprednisolone, and immunosuppressants, such as mycophenolate or cyclophosphamide [11]. Maintenance therapy includes oral prednisone and mycophenolate or azathioprine [11]. Despite advances in immunosuppressive regimens, including the use of corticosteroids, mycophenolate, and biologics such as belimumab and voclosporin, many patients do not achieve remission and may experience relapse [11]. Moreover, long-term immunosuppressant use carries substantial risks such as infection or renal and hepatic toxicity [11]. These challenges illustrate the urgent need for biomarkers and therapeutic targets that enable personalized treatment strategies, which emerging microbiome research might help guide. A renal biopsy is also used to confirm the class and thus guide therapy. Angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers are also commonly used for the control of proteinuria and as first-line antihypertensive agents [8].
The human microbiome refers to the vast collection of microorganisms, including bacteria, viruses, fungi, and archaea, that inhabit various anatomical sites of the body, such as the gut, skin, oral cavity, vagina, and urinary tract [12]. These microbial communities play an integral role in host immune function, metabolism, barrier integrity, and even neurocognitive development. Over the past two decades, advances in microbial profiling have allowed scientists to expand their understanding of the microbiome’s functional capacity. This has contributed to a paradigm shift in medicine, from viewing microbiomes solely as pathogens to recognizing them as critical modulators of health and disease across multiple organ systems.
Advances in sequencing technologies have revealed the microbiome’s involvement in a wide range of diseases, including inflammatory bowel disease, diabetes, cardiovascular disease, neuropsychiatric disorders, autoimmune conditions such as SLE, and reproductive complications such as PE [12]. Clinically, the microbiome is now used both diagnostically and therapeutically, with interventions such as fecal microbiota transplantation, probiotics, and dietary modulation in patients [12]. The evolving clinical applications of the microbiome extend beyond diagnostics. With the emergence of “precision microbiome editing, live biotherapeutics, and postbiotic interventions,” the microbiome is increasingly viewed as both a diagnostic biomarker and treatment target [13]. A myriad of trials are exploring microbiota modulation to improve outcomes in several pathologies such as sepsis, transplant medicine, and autoimmune diseases. With antibiotic resistance increasing and more adverse effects of biologic therapies coming to light, microbiome-based strategies may become a focus across many fields as potential treatment alternatives. As a result, understanding the microbiome’s compositional shifts and the resulting functional disruption in the host is vital, particularly in complex systemic conditions such as LN and PE, where the interplay between immunity, inflammation, and barrier dysfunction plays a central role in the patient’s health [13]. Despite extensive research on the microbiome in each disease separately, no comprehensive synthesis has compared their microbial profiles directly. This review aims to address this gap by evaluating and comparing the microbiota composition in SLE-LN and PE and identifying opportunities for diagnostic and therapeutic advancements.
Review
Methodology
Study Design
An in-depth scoping review was conducted to compare microbiome alterations between patients with LN and those with PE. This review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) recommendations. Human observational studies were analyzed as the primary evidence base, while animal studies were included separately to provide mechanistic support and were not interpreted as direct clinical evidence. This scoping review aimed to capture the breadth of existing literature and therefore included a range of study designs, including human primary studies, animal studies, and review articles.
Search Strategy
A literature search was performed using PubMed as the primary database, limiting articles published between January 2020 and April 2025. To ensure validity, two separate searches were conducted, one for LN and one for PE. Search terms included “lupus nephritis” AND “microbiome” for LN, and “preeclampsia” AND "microbiome” for PE.
Study Selection Criteria
Eligible studies included those published in English between 2020 and 2025, accessible through institutional resources, and focused on human or animal models that investigated the association between LN or PE and microbiome composition. Study designs included animal model experiments, human observational cohorts, review articles, and systematic reviews. Exclusion criteria included studies focused on non-microbiome mechanisms, studies lacking a control group, studies not relevant to the research question, and case reports.
Study Layout
Most human studies analyzed fecal bacterial communities using 16S rRNA sequencing, with sample sizes ranging from 38 to 221. Additional analytical methods included shotgun metagenomic sequencing, qPCR, in situ hybridization, gas chromatography, and bioinformatics.
For LN, 37 records were identified. After screening, 33 full-text articles were reviewed, and 16 met the inclusion criteria for this review (Figure 1). Studies focusing primarily on skin, genital, urinary, or fungal microbiota were excluded from the primary comparative analysis due to the limited availability of studies directly linking these microbial sites to LN-specific outcomes. While genital microbiome changes have been described in PE, comparable controlled data in lupus nephritis remain limited, which restricts disease comparison.
Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) flowchart for lupus nephritis microbiome alterations.
For PE, a total of 30 articles examining the microbiome alterations were retrieved. One article was excluded due to a lack of full-text access, leaving 29 articles for screening. Screening was performed by reviewing abstracts and removing out-of-scope articles, followed by full-text analyses. After screening, 11 articles met all inclusion criteria and were included in this review (Figure 2).
Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) flowchart for preeclampsia microbiome alterations.
Data Extraction
Data extraction was performed independently by three reviewers and included study type, population type, analysis method, sample size, and key findings. Google Docs was implemented to organize article reviews for improved extraction and analysis of overarching themes. Once finalized, EndNote was used to manage citations and references. No additional unpublished data were requested from the original study authors.
Results
Lupus Nephritis: Microbiome Findings
The phylum Proteobacteria was reported to be elevated in multiple studies, including Cheng et al., Wang et al., Parodi et al., Wu et al., and Yu et al. [14-18]. Expansions of Ruminococcus gnavus (R. gnavus) were frequently noted during LN flares or in patients with more severe disease [19,20]. Supporting this, Silverman et al. demonstrated that R. gnavus strains isolated from LN patients could trigger gut barrier dysfunction and activate the immune system in lupus-prone mice, suggesting these bacteria may play a direct role in worsening disease [21]. Additionally, Gui et al. reported that overgrowth of Escherichia, particularly Escherichia coli, was enriched in LN patients and associated with greater disease severity [22]. In this study, researchers also found that fecal transfer of these bacteria worsened kidney damage and immune activation in lupus-prone mice [22]. Similarly, Cheng et al. found significant enrichment of Enterobacteriaceae, Escherichia-Shigella, and *E. coli *among LN patients, alongside reduced abundance of Faecalibacterium and Clostridia [18]. Among systematic reviews, Wang et al. confirmed consistent reductions in diversity and the Firmicutes/Bacteroidetes (F/B) ratio in LN, along with enrichment of Proteobacteria and specific taxa such as R. gnavus and Bacteroides thetaiotaomicron [16]. However, Tan et al. noted that the F/B ratios in SLE and LN are not consistent across all studies [23]. Gui et al. found an increased Proteobacteria-to-Bacteroidetes (P/B) ratio in LN patients [22]. An additional study found that decreases in Firmicutes and the F/B ratio were not statistically significant [15].
In addition to consistent taxonomic shifts across LN patient studies, animal models also support a link between gut microbial alterations and disease severity. A study comparing genetically identical MRL/lpr mice (a lupus-prone murine model with a Fas gene mutation) from two different facilities reported significant differences in alpha diversity as measured by the Shannon Index (SI), along with distinct microbial compositions that correlated with differences in disease severity [24]. Mice from one colony developed more severe glomerulonephritis, which was associated with a higher abundance of Lactobacillaceae, whereas the other colony showed enrichment of Lachnospiraceae [24]. Further evidence for the role of gut dysbiosis in LN comes from studies of mesenchymal stem cell (MSC) therapy. Early MSC transplantation in MRL/lpr mice increased gut microbial diversity and was associated with improved kidney outcomes and enrichment of potentially beneficial strains such as Lactobacillus johnsonii and Romboutsia ilealis [25]. Gao et al. similarly demonstrated that fecal microbiota transplantation from healthy donors into lupus-prone mice led to expansion of Lactobacillus johnsonii and the metabolite inosine [25]. In another study, oral administration of Bacteroides fragilis to lupus-prone MRL/lpr mice reduced kidney pathology and serum autoantibody levels compared to untreated controls [26]. Fecal microbiota transplantation is presented only as an experimental approach in animal studies to support mechanistic interpretation and is not intended to suggest clinical relevance or therapeutic use. A summary of microbiome findings in human studies and animal studies is provided in Table 4 and Table 5.
Preeclampsia: Microbiome
The studies summarized in Table 6 demonstrate consistent microbiome alterations in human patients and animal models with PE, characterized by an increase in pro-inflammatory microorganisms and a decrease in beneficial microorganisms. In a 2020 case-control study by Chen et al., fecal microbiota transplantation was used to determine the relationship between the gut microbiota and the development of PE [30]. Patients with PE showed an enrichment of opportunistic bacteria, including Fusobacterium and Veillonella, while levels of beneficial bacteria, including Faecalibacterium and Akkermansia, were significantly decreased [30]. Correspondingly, levels of cytokines were markedly elevated in placental tissue from PE patients and PE-fecal microbiota transplantation mouse models [30]. Jin et al. reported a reduction in short-chain fatty acid (SCFA)-producing bacteria among patients with PE, including decreased levels of anti-inflammatory bacteria, such as Lachnospira and Dialister, and an increase in pro-inflammatory bacteria, including Blautia species [31]. Continued analysis revealed increased gene expression of pro-inflammatory cytokines such as IL-6 and IL-8 in early preeclampsia (EP) when compared to late preeclampsia (LP) (p = 0.005), further emphasizing the correlation between increased inflammation and the development of PE [31]. Overall, pathogenic genera, including Fusobacterium, Veillonella, Prevotella, Bacteroides, Enterobacteriaceae, and phyla Proteobacteria and Actinobacteria, increased across studies, while levels of commensal and SCFA-producing bacteria such as Akkermansia, Faecalibacterium, Lachnospira, Bifidobacterium, Subdoligranulum, and* Eubacterium* species decreased. The results of these studies also suggest that restoration of SCFAs may facilitate trophoblast invasion and decrease the risk of PE [31]. A summary of these microbiome changes in PE may be seen in Table 6.
Discussion
Lupus Nephritis: Alpha Diversity
Across the included studies, a consistent reduction in gut microbiome alpha diversity was observed in LN patients compared with healthy controls or SLE patients without nephritis. Wu et al. assessed alpha diversity using the Chao1 and Simpson indices, both showing significantly reduced diversity in LN patients [14]. Gui et al. evaluated alpha diversity using observed operational taxonomic units (OTUs) and the SI, reporting significantly lower richness and diversity in SLE patients, with more pronounced reductions in those with LN [22]. Wang et al. assessed alpha diversity using observed OTUs, Chao1, and the SI, all of which were significantly reduced in SLE patients, though the equitability index did not differ significantly between groups, indicating preserved evenness despite reduced richness [29]. Yu et al. reported significantly reduced alpha diversity using observed OTUs, Shannon, and Chao1 indices in LN patients compared to healthy controls [15]. Additionally, the systematic review by Wang et al. and the review article by Parodi et al. summarized consistent evidence of decreased alpha diversity across LN studies [16,17]. Cabana-Puig et al. also demonstrated comparable reductions in alpha diversity in lupus-prone mice [24]. In contrast, Cheng et al. found no significant differences in alpha diversity between LN patients, SLE patients without nephritis, and healthy controls [18].
Lupus Nephritis: Beta Diversity
Across the included studies, significant differences in beta diversity were also observed. Azzouz et al., Wang et al., Cheng et al., and Gui et al. reported distinct clustering patterns or greater variability in microbial community composition among LN patients compared to healthy controls or SLE patients without nephritis [18,19,22,29].
Lupus Nephritis: Microbiome Findings
Several prior studies have identified a reduced F/B ratio as a hallmark of gut dysbiosis in SLE when compared to healthy controls, with a lower ratio associated with increased disease activity [41-43]. This pattern has also been noted in other inflammatory diseases such as rheumatoid arthritis [44]. However, whether a reduced F/B ratio is consistently present in LN remains unclear. In the observational studies reviewed here, we did not consistently observe a reduced F/B ratio among LN patients, contrasting with findings more commonly described in narrative reviews and highlighted in one systematic review [16]. Our analysis yielded three main findings: increased abundance of Streptococcus,* E. coli*, and R. gnavus in LN patients. Additional research is needed to clarify the role of the F/B ratio in LN, as it is possible that this ratio might shift during progression from SLE to SLE-LN. Additionally, it is important to consider that these studies may have prioritized microbial taxa that directly contribute to LN pathogenesis rather than broad metrics such as the F/B ratio. For example, the lack of reduced F/B ratio reported by Gui et al. may reflect a subset of SLE-LN patients with distinct microbial profiles [22]. The prominence of R. gnavus and other specific taxa suggests that SLE-LN may have a unique microbial pattern that amplifies renal inflammation, potentially extending beyond the systemic dysbiosis noted in SLE. The limited reporting of the F/B ratio in primary LN studies emphasizes the need for additional research directly comparing the microbiome between SLE without nephritis and SLE-LN.
Preeclampsia: Alpha and Beta Diversity
A study conducted by Tian et al. analyzed the gut microbiome of early-onset PE (EOPE), late-onset PE (LOPE), and healthy pregnant controls (HPC) using 16S rRNA gene sequencing [32]. EOPE was associated with reduced alpha diversity (α-diversity) and depletion of SCFA-producing bacteria, which may contribute to the endothelial and placental dysfunction observed in PE patients [32]. A 2025 study by Han et al. analyzed the diversity and overall composition of the gut microbiota in PE patients by using fecal samples [37]. PE patients showed significant differences in beta diversity (β-diversity) when compared to the control group, while α-diversity did not differ significantly. More specifically, levels of Prevotella,* Dorea*, and Erysipelotrichaceae_UCG-003 were significantly increased in the PE group compared to the control group, while Subdoligranulum,* Parabacteroides*, and Bacteroides were significantly decreased [37]. Although α-diversity findings vary across studies, reductions have been associated with notable dysbiosis in PE [32,39].
Preeclampsia: Short-Chain Fatty Acids and Their Impact on the Microbiome
SCFAs are short-chain metabolic byproducts produced through the gut and have been associated with the development of hypertension [45]. Among various SCFAs, the three found in the highest concentrations in the gut include acetic acid (acetate), propionic acid (propionate), and butyric acid (butyrate) [45]. Studies have shown that patients with PE exhibit lower levels of SCFAs, which may contribute to an increased inflammatory state [46]. It has been hypothesized that SCFAs are able to modify blood pressure and inflammation through binding to secondary messenger systems; more specifically, to G protein-coupled receptors (GPCRs) expressed in renal tissue [45]. By binding to GPCRs and modulating the NF-κB signaling pathway, higher levels of SCFAs promote increased downstream production of T-regulatory cells (T regs) and a decreased state of inflammation [45]. Jin et al. found that treating PE-model rats with a propionate-producing bacterium such as *Akkermansia muciniphila *led to decreased blood pressure and improved placental outcomes [31]. The variability of SCFA content found in patients with PE may be of diagnostic value when used in early screening for PE. A study by Li et al. found significantly lower levels of caproic acid and butyrate in women with PE when compared to healthy controls [47].
Lupus Nephritis: Short-Chain Fatty Acids Impact on the Microbiome
Narrative reviews by Mohd et al. and Parodi et al., as well as a systematic review by Wang et al., report that gut microbiota dysbiosis in SLE patients is associated with increased fecal SCFA levels and a reduced F/B ratio [16,17,28]. These reviews suggest that imbalances in SCFAs may contribute to SLE pathogenesis by promoting a pro-inflammatory, dysbiotic state. In SLE patients, particularly those with LN, Tan et al. noted that reduced levels of SCFA-producing bacteria, including Faecalibacterium and Roseburia, lead to lower SCFA levels, which may disrupt regulatory T-cell function and elevate pro-inflammatory cytokines such as IL-6 [23]. This shift may potentially exacerbate kidney damage through increased gut permeability. Similarly, Wu et al. demonstrated in a mixed human-animal study that SCFA supplementation through *Bifidobacterium-*derived metabolites restored immune balance and mitigated kidney damage in lupus-prone mice, suggesting a potential protective role for SCFAs [14]. Collectively, these findings regarding SCFAs further emphasize the need for primary research directly comparing the microbiome role in SLE-non LN versus LN. SCFAs may play a role in the progression from SLE to LN, and supplementation could offer therapeutic benefits.
Overlap and Distinguishing Characteristics
This review reveals both shared and distinct gut microbiome features in LN and PE. Enrichment of Proteobacteria (5/11 LN studies; 3/10 PE studies) and Escherichia (4/11 LN; 2/10 PE) reflects a shared pro-inflammatory dysbiosis, as Proteobacteria expansion can disrupt gut barrier integrity, increase intestinal permeability, and promote systemic inflammation with renal consequences [48]. Both conditions demonstrate reduced Faecalibacterium (2/11 LN; 4/10 PE) and a less consistent depletion of Bifidobacterium (reported in LN by Wu et al. and in PE by Chang et al.), suggesting that the loss of anti-inflammatory taxa may further exacerbate inflammation [39,40].
Distinct microbial patterns were found to be disease-specific: *Ruminococcus gnavus (R. gnavus) *correlated with LN severity, potentially by promoting autoantibody generation or immune over-activation, while Fusobacterium enrichment and Akkermansia depletion in PE may reflect pregnancy-specific inflammation or placental dysfunction [49]. LN also demonstrated increased Streptococcus, whose antigens may trigger molecular mimicry and cross-reactive autoantibody production, further contributing to renal injury [50]. These findings suggest therapeutic potential for microbiome modulation. Restoring Faecalibacterium and Bifidobacterium through probiotics or high-fiber diets may benefit both diseases; while targeting R. gnavus may be more relevant to LN, and reducing Fusobacterium, via diet or vaginal probiotics, may be more pertinent to PE. Moreover, these taxa hold promise as biomarkers, with R. gnavus linked to LN severity and Fusobacterium associated with PE.
Beyond the gut, Monticolo et al. identified Ureaplasma urealyticum and Ureaplasma parvum in the vaginal microbiota of SLE patients, though these findings may have been confounded by concurrent steroid or antibiotic use; importantly, data on genital dysbiosis in LN remain limited [51]. Shi et al. further demonstrated reduced urinary microbiome diversity and identified potential urinary biomarkers in SLE [52]. In PE, interest has expanded to maternal microbial niches, including the gut, vagina, oral cavity, and placenta, where enrichment of Bacteroides,* Fusobacterium*, and Prevotella,* *and depletion of Lactobacillus, Faecalibacterium, and Bifidobacterium have been reported [38-40,53]. These microbial shifts are associated with increased lipopolysaccharide burden, heightened Toll-like receptor signaling, systemic inflammation, and reduced SCFA production, while microbial metabolites such as SCFAs, trimethylamine N-oxide, and bile acids may further influence endothelial and placental function [31,40].
Limitations and Future Directions
Future research can take several directions to address key gaps and advance this comparative work. For SLE-LN, the inconsistent findings regarding the F/B ratio highlight the need for studies directly comparing SLE patients with and without nephritis. Additional research is needed for further characterization of microbiome findings in EOPE and LOPE, as well as to explore how differing dietary and nutritional factors influence microbiome composition.
One limitation of this review includes the comparison of findings across varying sample types. For example, microbial samples were derived from gut, vaginal, and placental microbiomes, which differ significantly in baseline microbial composition. Moreover, microbial detection methods were not standardized across studies and included techniques such as 16S rRNA sequencing, gas chromatography, shotgun metagenomic sequencing, and polymerase chain reaction. Future research directions may benefit from focusing on specific microbiome sample types and comparing the content of organisms found in each.
Recent research has begun to explore the role of the gut and urinary microbiome in modulating immune responses in SLE and its renal manifestation, LN. Microbial dysbiosis, particularly in the gut, may contribute to both disease initiation and progression, offering a novel lens through which to understand autoimmunity. This emerging insight has opened new avenues for examining host-microbiome interactions not only as potential biomarkers but also as therapeutic targets in LN and other inflammatory renal conditions, such as PE. However, unlike PE, which arises primarily from placental dysfunction, LN develops from systemic autoimmunity. Multiple studies in our review demonstrated that patients with SLE and LN exhibit significant gut dysbiosis, characterized by reduced abundance of immunoregulatory taxa such as Faecalibacterium and increased levels of pro-inflammatory bacteria, including R. gnavus, Enterococcus, and other pathobionts [14,18,19,21,22].
Lupus Nephritis: Possible Therapeutic Implications
Azzouz et al. reported an increased abundance of R. gnavus in nearly half of patients experiencing LN disease flares [19]. This expansion was accompanied by the expression of a novel lipoglycan recognized by elevated immunoglobulin G2 (IgG2) antibody levels, suggesting that *R. gnavus *may serve as a potential biomarker for disease monitoring and a target for further therapeutic inventions [19].
Additionally, Cheng et al. identified distinct bile acid profiles in SLE patients with and without nephritis [18]. Elevated levels of Glycocholic acid and Glycochenodeoxycholic acid were observed in SLE-non LN patients, whereas reduced bile acid levels and increased Mead acid levels were noted in SLE-LN patients [18]. A negative correlation between Escherichia-Shigella abundance and bile acid concentrations was also reported. These metabolic alterations suggest dysfunction of the gut-kidney axis in LN, highlighting a potential role for dietary modulation in managing renal inflammation. Zhu et al. further explored the impact of synbiotics on gut microbiota in new-onset LN patients through metagenomic and metabolomic sequencing [54]. Following administration of synbiotics in addition to prednisone acetate and cyclophosphamide, analysis of patients’ fecal samples revealed a decrease in pathogenic bacteria, including Prevotella, Bacteroides, and Enterobacteriaceae unclassified, and an increase in Actinobacteria and Firmicutes [54]. Corresponding alterations in metabolic pathways, such as amino acid biosynthesis and purine metabolism, were observed and correlated with clinical data. These findings suggest that synbiotics may serve as a promising adjunctive therapy for LN.
Preeclampsia: Possible Therapeutic Implications
A 2021 study analyzing various maternal microbiome sites, including placental, vaginal, and gut samples, demonstrated increased dysbiosis among patients with PE [34]. Probiotics, prebiotics, or nutritional modification may serve as new therapeutic strategies for PE by combating this dysbiosis and restoring physiologic homeostasis [34]. Patients with PE consistently exhibit reduced fecal microbiome diversity. Notably, fecal levels of butyrate were significantly decreased in PE, whereas exogenous butyrate supplementation significantly lowered blood pressure in the LPS-induced hypertensive rat models [40]. This correlation supports the potential role of dietary modifications aimed at increasing butyrate levels as a treatment approach for patients with PE [40]. Similarly, a study by Sun et al. concluded that PE rat models treated with probiotics experienced increased levels of Bifidobacterium, Lactobacillus, and nitric oxide (p < 0.05), as well as normalized blood pressure compared to untreated controls [53]. These findings further highlight the potential benefits of probiotic-mediated microbiome restoration. Additional studies have highlighted the role of an overactive innate immune system in PE, particularly through the Toll-like-receptor 4 system signaling pathway, which promotes the secretion of inflammatory cytokines and inhibits effective trophoblast invasion [55]. Management approaches that attenuate Toll-like-receptor-mediated inflammation may help reduce PE severity.
Restoring SCFA levels in patients with PE may support trophoblast implantation and outweigh negative microbiome changes associated with the disease. Additionally, vitamin D has recently emerged as a potential modulator of the microbiome. Lower levels of vitamin D have been associated with PE, and a 2023 study by Ma et al. showed that vitamin D supplementation reduced inflammation and increased SCFA-producing bacteria in the microbiome [37,55].
Conclusions
This study underscores the critical role of a pro-inflammatory microbiome in the pathogenesis of both PE and LN. Shared microbial features, such as elevated Proteobacteria and Escherichia, suggest dysbiosis as a common driver of inflammation, while disease-specific alterations, including Fusobacterium enrichment and Akkermansia depletion in PE, and* Ruminococcus gnavus* expansion in LN, highlight distinct mechanisms. These microbial shifts contribute to systemic inflammation, endothelial dysfunction, immune dysregulation, and organ damage, converging on inflammatory pathways characterized by elevated tumor necrosis factor-α, interleukin-6, and related cytokines. The identification of both shared and unique microbial signatures offers opportunities for developing diagnostic biomarkers, targeted microbiome-modulating therapies, and personalized treatment strategies. Future longitudinal and interventional studies are needed to clarify causal relationships and determine whether modifying the microbiome can improve maternal-fetal outcomes in PE or mitigate renal injury in LN. Such insights may pave the way for precision medicine approaches in both obstetric and autoimmune disease contexts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1The global impact of pre-eclampsia and eclampsia Semin Perinatol Duley L 1301373320091946450210.1053/j.semperi.2009.02.010 · doi ↗ · pubmed ↗
- 2Soluble endoglin and other circulating antiangiogenic factors in preeclampsia N Engl J Med Levine RJ Lam C Qian C 992100535520061695714610.1056/NEJ Moa 055352 · doi ↗ · pubmed ↗
- 3Preeclampsia: pathophysiology, challenges, and perspectives Circ Res Rana S Lemoine E Granger JP Karumanchi SA 1094111212420193092091810.1161/CIRCRESAHA.118.313276 · doi ↗ · pubmed ↗
- 4Immunology of pre-eclampsia Am J Reprod Immunol Redman CW Sargent IL 5345436320102033158810.1111/j.1600-0897.2010.00831.x · doi ↗ · pubmed ↗
- 5Gut microbiota changes in preeclampsia, abnormal placental growth and healthy pregnant women BMC Microbiol Huang L Cai M Li L 2652120213460755910.1186/s 12866-021-02327-7PMC 8489045 · doi ↗ · pubmed ↗
- 6Review: maternal health and the placental microbiome Placenta Pelzer E Gomez-Arango LF Barrett HL Nitert MD 30375420172803446710.1016/j.placenta.2016.12.003 · doi ↗ · pubmed ↗
- 7Vascular dysfunction in preeclampsia Cells Opichka MA Rappelt MW Gutterman DD Grobe JL Mc Intosh JJ 30551020213483127710.3390/cells 10113055 PMC 8616535 · doi ↗ · pubmed ↗
- 8Lupus nephritis Nat Rev Dis Primers Anders HJ Saxena R Zhao MH Parodis I Salmon JE Mohan C 7620203197436610.1038/s 41572-019-0141-9 · doi ↗ · pubmed ↗