Metal uptake systems underpin Enterococcus faecalis virulence in both non-diabetic and diabetic wound infection models
Debra N. Brunson, Ling Ning Lam, Shivani Kundra, Shannon M. Wallet, José A. Lemos

TL;DR
This study shows that metal uptake systems are crucial for the bacteria Enterococcus faecalis to cause wound infections, and that these systems behave differently in diabetic versus non-diabetic wounds.
Contribution
The study reveals how metal acquisition systems influence E. faecalis virulence in wound infections and highlights differences in nutritional immunity between diabetic and non-diabetic wounds.
Findings
E. faecalis strains lacking iron, manganese, or zinc uptake systems show reduced virulence in non-diabetic wound models.
Diabetic wounds delay nutritional immunity, particularly iron restriction, compared to non-diabetic wounds.
The Δ5Fe strain temporarily colonizes diabetic wounds better than non-diabetic wounds.
Abstract
Wound infections remain an important medical problem, which is aggravated by the prevalence of multidrug-resistant bacteria. Among them, Enterococcus faecalis is a major pathogen of surgical site incisional and diabetic chronic wounds, but factors driving its colonization and persistence in wounds remain poorly understood. Iron, manganese, and zinc are essential cofactors in cellular processes, prompting the host to restrict their availability through mobilization of metal-sequestering proteins, a defense known as nutritional immunity. Previously, we showed that E. faecalis strains lacking key iron (Δ5Fe), manganese (Δ3Mn), or zinc (Δ2Zn) uptake systems have impaired virulence. Here, we used an excisional wound model in normoglycemic (C57Bl/6J or B6) and diabetic (C57Bl/6J lepR−/− or DB) mice to examine the role of these metal import systems in wounds. The strong upregulation of metal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
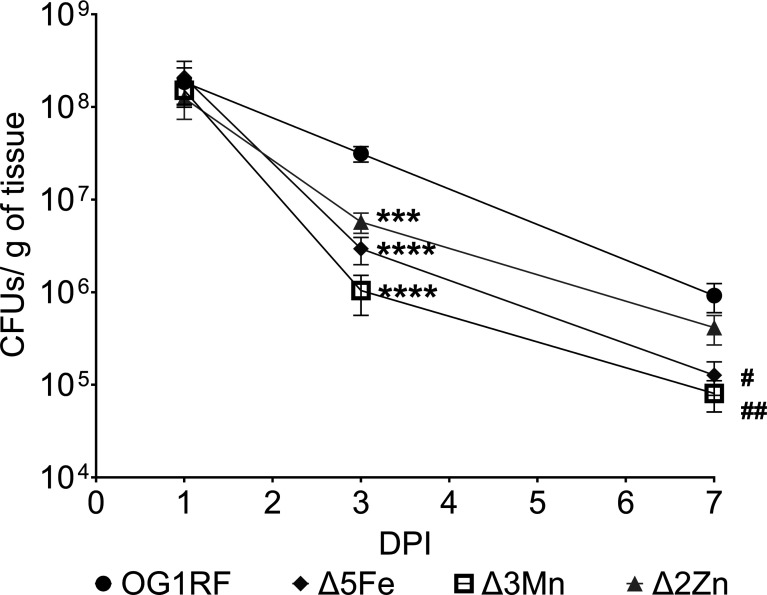 Fig 1
Fig 1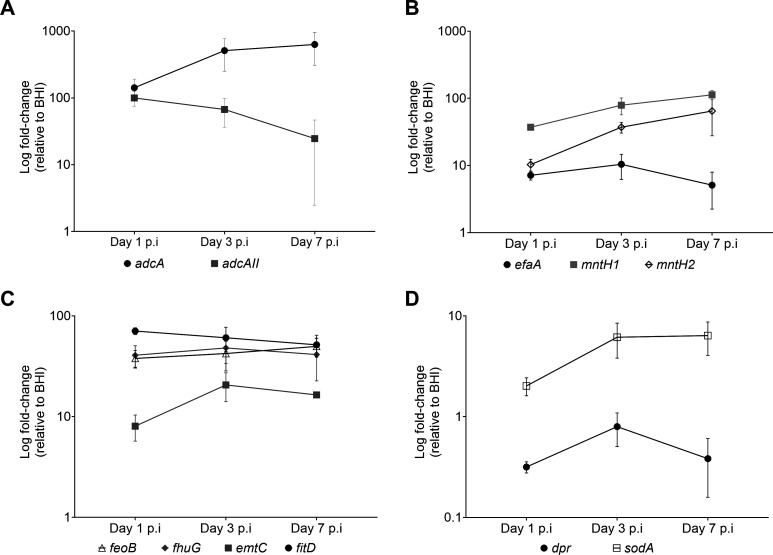 Fig 2
Fig 2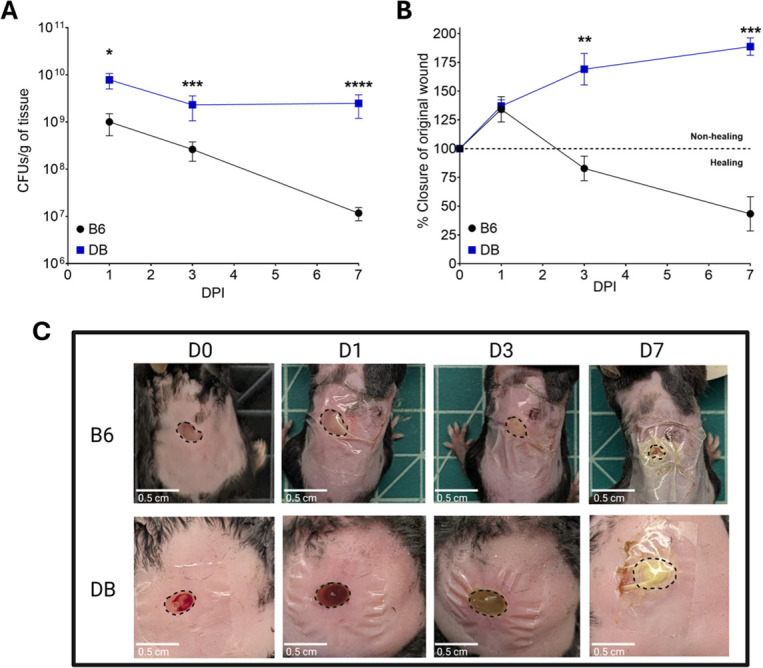 Fig 3
Fig 3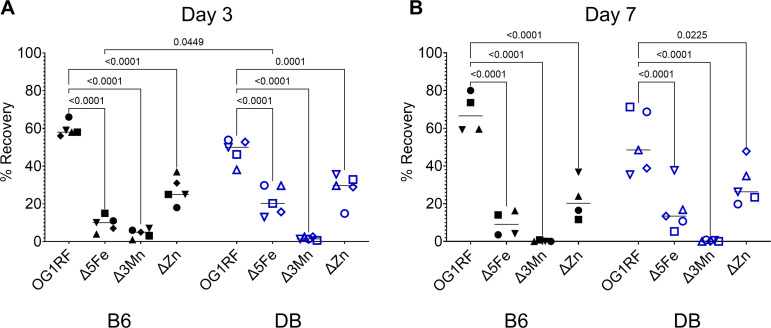 Fig 4
Fig 4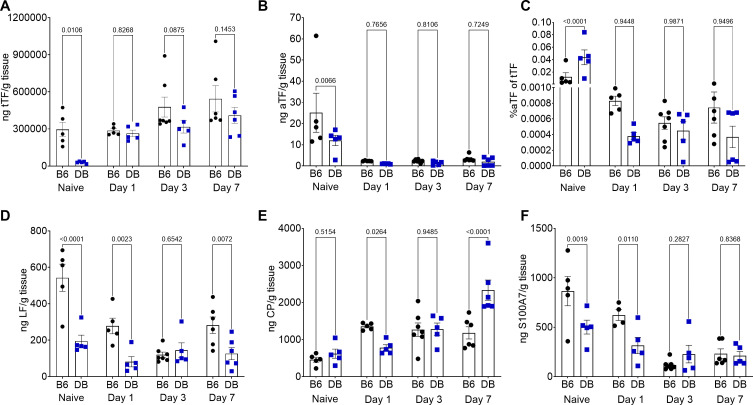 Fig 5
Fig 5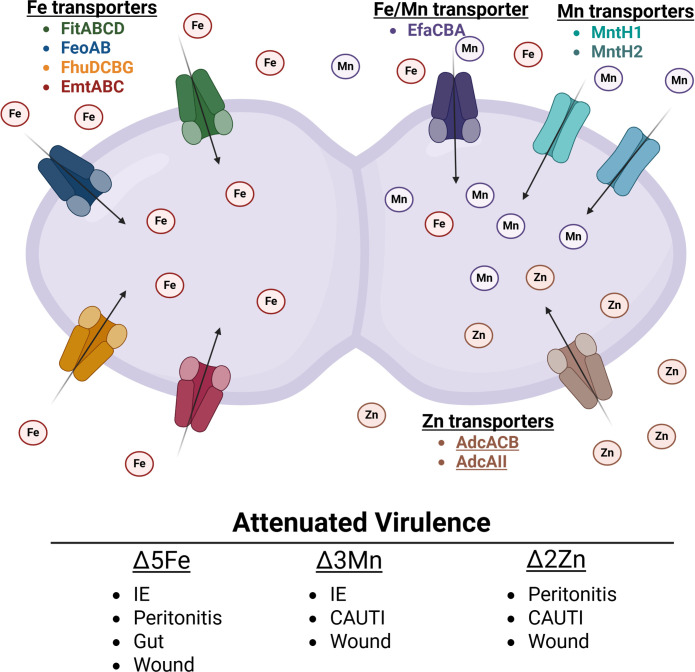 Fig 6
Fig 6| Primer name | Sequence (5′ → 3′) | Gene locus |
|---|---|---|
|
| OG1RF_RS00260 | |
|
| ||
|
| OG1RF_RS08040 | |
|
| ||
|
| OG1RF_RS04395 | |
|
|
| Primer name | Sequence (5′ → 3′) | Gene locus |
|---|---|---|
|
| OG1RF_RS00260 | |
|
| ||
|
| OG1RF_RS12625 | |
|
| ||
|
| OG1RF_RS08610 | |
|
| ||
|
| OG1RF_RS08040 | |
|
| ||
|
| OG1RF_RS04395 | |
| mntH2 R′ |
| |
|
| OG1RF_RS12060 | |
|
| ||
|
| OG1RF_RS12595 | |
|
| ||
|
| OG1RF_RS01950 | |
|
| ||
|
| OG1RF_RS00700 | |
|
| ||
|
| OG1RF_RS12695 | |
|
| ||
|
| OG1RF_RS01890 | |
|
| ||
|
| OG1RF_RS00040 | |
|
|
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —American Heart Associationhttp://dx.doi.org/10.13039/100000968
- —American Heart Associationhttp://dx.doi.org/10.13039/100000968
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Trace Elements in Health · Wound Healing and Treatments
INTRODUCTION
Wound infections cover a range of soft tissue complications, including those resulting from incisional surgery, burns, and traumatic injuries. Estimates for yearly global wound care expenditure are almost $300 billion, and with an expansion in the aging populations and comorbidities, such as diabetes and hypertension, this number is only expected to increase over time (1, 2). In healthy individuals, the wound healing process follows four tightly controlled overlapping stages: (i) hemostasis, which occurs immediately after injury and leads to the formation of blood clots, (ii) inflammation, which occurs 1–3 days after injury and involves clearance of any bacteria present and necrotic debris to prepare the wound bed for new growth, (iii) proliferation, which can last days or weeks and includes re-epithelialization, restoration of the vasculature, and formation of granulation tissue, and (iv) remodeling, which can take several weeks and even years and involves the development of tensile strength of the new tissue and complete closure of the wound (3, 4). This process depicts what typically occurs in a normal healing wound; however, due to internal and/or external factors, some wounds stall in their progression and become chronic non-healing wounds, commonly associated with unresolved inflammation, elevated levels of reactive oxygen species, limited blood circulation, and oftentimes, become infected with opportunistic wound pathogens, such as Staphylococcus aureus, Pseudomonas aeruginosa, Escherichia coli, and Enterococcus faecalis (2, 5). One of the most common and difficult types of chronic wounds to treat is “diabetic wounds,” notably diabetic foot ulcers, typically associated with type II diabetes (T2D) or insulin resistance (6–8). These infections can often lead to lengthy hospital stays, amputations, sepsis, and death (7). As such, a better understanding of both non-diabetic and diabetic wound environments and the underlying mechanisms that dictate host–pathogen interactions can pave the way for the development of new therapies to prevent and treat wound infections.
Of interest, an important host defense mechanism used to combat infections is nutritional immunity, which stems primarily from innate immune cells secreting copious amounts of proteinaceous metal chelators that starve invading pathogens of trace metals, such as iron, manganese, and zinc (9–11). These proteins include iron-binding proteins ferritin, which sequesters intracellular iron; transferrin, which binds iron in circulation (9, 12); lactoferrin, which restricts access to iron at mucosal surfaces and modulates the host inflammatory response (13–16), and lipocalin-2, which targets iron complexed to bacterial siderophores (17, 18). Additionally, S100A8/A9 or calprotectin, primarily produced and released by neutrophils at the site of infection, avidly binds divalent metals and is particularly known for depriving pathogens of manganese and zinc, while S100A7 (psoriasin), abundant in the epidermis, contributes to host defense through zinc sequestration (19–21). To overcome metal starvation, bacterial pathogens evolved sophisticated methods to maintain metal homeostasis that include expression of surface-associated high-affinity metal transporters, secretion of metallophores, and, eventually, activation of alternative metal-sparing enzymes (9, 22).
While our understanding of nutritional immunity and trace metal homeostasis in healthy (normoglycemic) individuals is relatively well understood (9, 16, 23–27), fundamental knowledge of these processes in the context of uncontrolled diabetes is limited. In agreement with clinical observations, prior studies that have used diabetic wound infection mouse models have shown that prevalent wound pathogens, including S. aureus, P. aeruginosa, and Streptococcus agalactiae, maintain significantly higher bacterial burdens in wounds of diabetic mice when compared to normoglycemic mice (28–31). These studies support the notion that higher bacterial titers are associated with an immunocompromised, nutrient-rich diabetic host environment that is less capable of restricting microbial growth (28, 30, 32–34). Along these lines, it is tempting to speculate that uncontrolled diabetes could also impair nutritional immunity activation in diabetic wounds, facilitating bacterial proliferation. In fact, a recent study has shown that S. agalactiae genes encoding zinc, manganese, and nickel transporters were highly expressed in infected wounds of non-diabetic mice compared to diabetic mice wounds (32). In a subsequent study, this same group demonstrated that the loss of these transporters impaired S. agalactiae colonization in non-diabetic wounds but had no effect in diabetic wounds, despite the higher levels of calprotectin present in diabetic wounds (28).
The World Health Organization has listed E. faecalis, a major nosocomial pathogen, as the fourth most common pathogen in surgical site infections (35). In addition, E. faecalis is amongst the most frequently isolated bacteria from diabetic wounds (36–39) and has been associated with a high degree of antibiotic treatment failure and poor outcomes that include progression to bone infection, eventual need for limb amputation, and sepsis-mediated death (37, 40, 41). Despite its high prevalence in wound infections, our understanding of the factors that promote E. faecalis colonization and persistence within the wound environment is still limited. A mouse model of enterococcal wound infection has demonstrated that E. faecalis modulates the wound micro-environment to evade immune clearance and delay healing (42). Furthermore, the ability of E. faecalis to undergo de novo purine biosynthesis and metabolize mannose and galactose has been shown to contribute to its capacity to colonize and persist within wounds (43). While previous works have focused on the microbial factors and host responses that contribute to wound infections, the importance of nutritional immunity and its possible association with chronic diabetic E. faecalis wound infections has not been investigated. For the past several years, our group has shown that the ability to acquire trace metals, iron, manganese, and zinc is critical for the virulence of E. faecalis (44–46). Here, we leveraged the availability of well-characterized E. faecalis mutants with major defects in manganese (46), iron (44), or zinc uptake (45) to assess their ability to colonize wounds in both normoglycemic and diabetic mice. While our findings underscore the importance of trace metal acquisition in E. faecalis wound infections and reveal nuanced differences in the expression of host-derived proteinaceous metal chelators in a T2D mouse model, a more definitive correlation between impaired nutritional immunity and chronic colonization of diabetic wounds by E. faecalis remains to be established.
RESULTS
Iron, manganese, and zinc uptake systems of E. faecalis are critical for wound colonization
We previously showed that the E. faecalis core genome encodes four iron ABC-type transporters, FeoAB, FhuDCBG, FitABCD, and EmtABC, as well as a dual iron/manganese ABC-type transporter, EfaCBA (44, 46). We also identified and characterized two manganese NRAMP-type transporters, MntH1 and MntH2, and an ABC-type zinc transporter that utilizes two interchangeable substrate-binding proteins, AdcA and AdcAII, which function with the shared AdcB permease and AdcC ATPase subunit (44–46). Using the invertebrate model Galleria mellonella and mammalian infection models (mouse and rabbit), we demonstrated that strains of E. faecalis with major defects in manganese (Δ3Mn), zinc (Δ2Zn), or iron (Δ5Fe) uptake exhibit significantly reduced virulence across all tested hosts (44–46). While previous work using the excisional wound mouse model showed that the Δ5Fe strain colonized wounds poorly (44), the roles of manganese or zinc acquisition during E. faecalis wound infection were not evaluated in those studies.
Here, we assessed the ability of the Δ3Mn and Δ2Zn strains to colonize excisional wounds in C57BL/6J (B6) mice and included the Δ5Fe strain to directly compare the relative importance of each metal during infection. In addition to examining the acute phase of infection (1 and 3 days post-infection), we also quantified bacterial colonization 7 days post-infection to evaluate the potential for chronic wound colonization. While there were no differences in the bacterial burden of wounds infected with the parent strain (OG1RF) and any of the individual metal uptake mutants at 1 day post-infection, all mutants were recovered at significantly lower levels 3 days post-infection. Among them, the Δ2Zn strain exhibited the smallest reduction (~1 log) compared to OG1RF, whereas the Δ3Mn strain showed the greatest decrease (~1.8 log) in bacterial load (Fig. 1). By day 7 post-infection, the difference in bacterial titers between wounds infected with the Δ2Zn strain and those infected with the parent strain was no longer significant, while both Δ3Mn and Δ5Fe continued to be recovered at significantly lower levels (~1 log) compared to OG1RF (Fig. 1).
*Seven-week-old B6 mice were wounded with a 6 mm biopsy punch and infected with 108 CFU of either E. faecalis OG1RF, Δ5Fe, Δ3Mn, or Δ2Zn strains. Wounds were excised at 1, 3, and 7 days post-infection (p.i.) and homogenized in PBS for CFU determination by plating on brain heart infusion (BHI) agar. Each point represents the average and standard deviation from at least nine infected mice. Outliers were removed with a ROUT standard outlier test. Statistical significance compared to the parental strain was determined using a Mann-Whitney test. Day 3: ***P ≤ 0.001, ***P ≤ 0.0001; day 7: #P ≤ 0.05, ##P ≤ 0.01.
Collectively, the results described above are aligned with previous studies proposing that essential trace metals are not readily bioavailable in wound microenvironments (28, 32, 43). To support this observation, we compared the transcriptional levels of E. faecalis genes involved in the uptake of iron (fitD, emtC, feoB, and fhuG), manganese (mntH1 and mntH2), both manganese and iron (efaA), and zinc (adcA, adcAII) between cells grown in brain heart infusion (BHI) medium and those recovered from infected wounds of B6 mice on days 1, 3, or 7. Notably, we have consistently shown that BHI contains low concentrations of iron (~5 µM), manganese (<1 µM), and zinc (~10 µM) (44, 46, 47), concentrations at which supplementation of iron, manganese, or zinc in the millimolar range has been shown to repress transcription of E. faecalis iron, manganese, and zinc import genes (48–50). Consistent with our expectations, transcription of iron, manganese, and zinc uptake genes was significantly elevated in bacteria from infected wounds relative to BHI-grown counterparts (Fig. 2A through C). Notably, there is a divergence in expression between different components of zinc and manganese transporters (Fig. 2A and B). Specifically, for the zinc transporters, adcA increases over time, and adcAII decreases over time, while manganese transporters mntH1 and mntH2 increase over time, and efaA shows a moderate reduction in expression over time. We also assessed the expression of dpr and sodA as markers of iron overload (dpr) and oxidative stress (both genes) in infected wound samples (51–53). Consistent with the upregulation of iron transporters, and indicative of iron starvation, dpr, which encodes the major iron storage protein, was significantly downregulated (Fig. 2D). In contrast, sodA transcription was modestly upregulated in the wounds, suggesting that E. faecalis may also experience ROS stress in the wound microenvironment. (Fig. 2D).
*Transcriptional profile (relative to BHI) of E. faecalis metal transporters in infected wounds of B6 mice at 1, 3, and 7 days post-infection. (A) Zinc transporters (adcA and adcAII), (B) manganese transporters (efaA, mntH1, and mntH2), (C) iron transporters (feoB, fhuG, emtC, and fitD), and (D) iron storage (dpr) and oxidative stress management (sodA). Statistical significance was determined using ordinary one-way ANOVA and Fisher’s LSD test. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001.
E. faecalis maintains higher bacterial burden in diabetic wounds than in non-diabetic wounds
Previous studies have demonstrated that diabetic mice exhibit higher bacterial burdens in infected wounds compared to normoglycemic mice (28–30). These authors proposed that, in addition to impaired immune responses, the nutrient-rich environment of diabetic wounds significantly contributes to this phenomenon (28, 30, 32–34). Here, we investigated whether E. faecalis, like other prevalent wound pathogens, exhibits enhanced infection in diabetic wounds compared to non-diabetic wounds. To address this, we compared E. faecalis OG1RF bacterial burden at 1, 3, and 7 days post-infection in wounds of B6 (normoglycemic) mice and C57Bl/6J lepR^−/−^ (DB) mice, which recapitulates the physiological and pathological features of T2D in humans (54). Compared to B6 mice, DB mice exhibited significantly higher bacterial titers (~1 log) in wounds at 1 and 3 days post-infection (Fig. 3A). This difference became much greater by day 7 (~3 log), as B6 mice continued to clear the infection between days 3 and 7, whereas the bacterial burden in DB mice remained unchanged. Moreover, the infected wounds of DB mice not only failed to heal but increased in size by ~30% between days 3 and 7, whereas wounds of B6 mice showed ~50% reduction in size over the same period (Fig. 3B). Visual examination revealed progressive healing of infected wounds in B6 mice over time, whereas wounds in DB mice appeared highly inflamed and exhibited extensive accumulation of cellular exudates, particularly at 7 days post-infection (Fig. 3C).
*Seven-week-old B6 or DB mice were wounded with a 6 mm biopsy punch and infected with 108 CFU of E. faecalis OG1RF. (A) Wounds were excised at 1, 3, and 7 days post-infection, homogenized, and bacterial CFU determined by plating on BHI agar. (B) Wound area was measured using ImageJ, and percent closure was determined by comparing to the initial wound area. (C) Images of wounds immediately after wounding (D0) and at 1, 3, and 7 days post-infection. Black dashed lines indicate the wound bed margins where each wound was measured. Each point represents the average and standard deviation from at least five infected mice. Outliers were removed using a ROUT outlier test. Statistical significance was determined using a Mann-Whitney test at each time point. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001.
Trace metal acquisition facilitates E. faecalis colonization of both normoglycemic and diabetic wounds
Similar to our findings (Fig. 2), a group at the University of Colorado previously demonstrated that S. agalactiae strains lacking either manganese or zinc uptake systems exhibit impaired wound colonization in normoglycemic mice (28). They also demonstrated that the impaired wound colonization of metal transport mutants was no longer observed in streptozotocin-induced hyperglycemic mice, a model commonly used to study type 1 diabetes (T1D) (28). To further probe the role of iron, manganese, and zinc acquisition in E. faecalis colonization of diabetic wounds in DB (T2D) mice, we performed a competitive wound colonization experiment to directly compare the relative virulence potential of parent and mutant strains in both B6 and DB mice. Briefly, wounds created in either B6 or DB mice were infected with an inoculum containing equal proportions of each strain (OG1RF, Δ5Fe, Δ3Mn, and Δ2Zn), and the relative abundance of each strain recovered from wounds was determined at 3 and 7 days post-infection. Regardless of the mouse background, all three mutants were outcompeted by the parental strain at both 3 and 7 days post-infection (Fig. 4A and B). Consistent with the single-strain infection experiments in B6 mice (Fig. 2), loss of the zinc uptake system (Δ2Zn) had the least impact on colonization fitness, whereas disruption of manganese uptake (Δ3Mn) had the most pronounced effect. When compared to B6-infected mice, the Δ5Fe strain was recovered at slightly higher rates (P = 0.0449) in wounds of the DB mice on day 3; however, this apparent fitness advantage did not persist by day 7. Collectively, these findings validate the critical role of trace metal acquisition in E. faecalis wound colonization but do not support that impaired nutritional immunity alone explains the increased permissiveness of the diabetic wound environment to infection.
Competitive wound colonization. B6 and DB mice were infected with an inoculum of 108 CFU containing an equal ratio of the E. faecalis OG1RF, Δ5Fe, 3Mn, and Δ2Zn strains. At (A) 3 and (B) 7 days post-infection, wounds were excised and homogenized in PBS for CFU determination by plating on BHI agar. Bacterial colonies were screened using gene-specific primers (Table 2). At least 100 colonies were screened from each animal to calculate the percentage recovery of each strain. Statistical significance was determined using a two-way ANOVA with a Šidák’s multiple comparison test. P values <0.05 were considered statistically significant, and the exact values are shown in the figure.
Metal homeostasis may differ between diabetic and non-diabetic wounds
The observation that the Δ5Fe strain was recovered at a modest yet significantly higher rate in DB wounds on day 3 (Fig. 4A) led us to suspect that trace metal bioavailability in the diabetic wound microenvironment may be temporally altered compared to non-diabetic wounds, with greater access to iron in the early stages of healing that diminishes over time. One possibility is that the production and mobilization of host-derived metal chelators, such as proteins from the S100 family, transferrin, and lactoferrin (9, 20), to the site of infection differ in diabetic wounds when compared to non-diabetic wounds. To test this, we quantified total transferrin (tTF), apo-transferrin (aTF—transferrin that is not bound to iron), lactoferrin (LF), calprotectin (CP/S100A8/A9), and psoriasin (S100A7) levels in skin tissues harvested from naïve and OG1RF-infected wounds in both B6 and DB mice. Interestingly, transferrin levels were approximately 10-fold higher in naïve skin tissue of B6 mice compared to DB mice; however, this difference was no longer evident at any time point in the infected wounds (Fig. 5A). Similarly, apo-transferrin levels were approximately twofold higher in naïve skin tissue of B6 mice compared to DB mice but were markedly reduced upon infection in both groups (Fig. 5B). This supports the notion that transferrin plays an active role in the host’s nutritional immunity strategy. Nevertheless, comparison of the percentage of apotransferrin of the total transferrin in B6 and DB mice revealed a slightly different pattern. Specifically, the percentage of aTF was approximately 3.5-fold higher in the naïve skin tissue of DB mice compared to B6 mice but declined by about 50% after 1 day of infection. By days 3 and 7, no statistically significant differences were observed between the groups (Fig. 5C).
Abundance of host metal chelators in naïve skin and infected wounds of B6 and DB mice. Naïve skin from biopsy punch and excised wounds from 1, 3, and 7 days post-infection were homogenized in metal-free PBS containing protease inhibitor. ELISA kits were used to determine the abundance of each host metal chelator and were normalized to grams of tissue. (A) Total transferrin (tTF), (B) apo-transferrin (aTF), (C) percentage aTF (apo-transferrin) over tTF (total transferrin) [%aTF = (aTF/tTF) × 100%], (D) lactoferrin (LF), (E) calprotectin (CP), and (F) S100A7 (psoriasin). Each data point represents the value from a single animal, with at least five animals used. Statistical significance was determined using a two-way ANOVA with a Šidák’s multiple comparison. P values <0.05 were considered statistically significant, and the exact values are shown in the figure.
Lactoferrin levels followed a similar pattern to total transferrin, with levels approximately 2.5-fold higher in B6 naïve skin tissue compared to DB mice. This difference persisted at day 1 (approximately threefold) and day 7 (approximately twofold) post-infection but was not observed on day 3 (Fig. 5D). While calprotectin levels were similar in naïve skin tissue of B6 and DB mice, infected B6 wounds showed significantly higher calprotectin levels (~50%) at 1 day post-infection compared to DB mice. Notably, calprotectin levels remained relatively stable over the 7-day infection period in B6 mice, whereas they increased steadily in DB mice, rising approximately 2.5-fold between days 1 and 7 post-infection (Fig. 5E). By day 7, calprotectin levels were nearly twice as high in DB mice compared to B6 mice. Considering that calprotectin is produced in large quantities by neutrophils and other immune cells in response to infection and is used as a gold standard marker for inflammation (20), this latter result was expected, given the higher bacterial burden and the highly inflammatory appearance of DB-infected wounds at day 7. Finally, psoriasin levels were approximately twofold higher in naïve skin tissue and at 1 day post-infection in B6 mice compared to DB mice (Fig. 5F). However, by days 3 and 7 post-infection, psoriasin levels declined to comparable levels in both mouse strains.
DISCUSSION
It is estimated that one-third of the bacterial proteome requires a metal cofactor (55), making the ability of bacteria to maintain an adequate intracellular supply of biometals essential for bacterial fitness (10). Accordingly, metal transporters (both import and export systems), metal chaperones, and metal storage proteins are consistently linked to bacterial virulence (10, 22, 56–58). Using an excisional wound mouse model, we confirmed previous findings that the loss of multiple iron transporters impairs wound colonization and extended this observation to include manganese and zinc transporters. In addition, we demonstrate that diabetes greatly increases host susceptibility to enterococcal wound infection during both the acute and chronic phases.
While the roles of iron, manganese, and zinc import in E. faecalis virulence are relatively well defined (Fig. 6) (44–46), the factors regulating the expression and activity of individual trace metal transporters, particularly within the wound microenvironment, remain poorly understood. Previous studies have shown that transcription of the N-ramp manganese transporter mntH2 and dual iron and manganese ABC transporter (efaCBA) is strongly induced when E. faecalis is grown ex vivo in human blood or human urine, as well as in vivo using an intra-peritoneal challenge (peritonitis) mouse model (59–61). Furthermore, our lab has shown that the three high-affinity transport systems associated with manganese uptake are differentially regulated in two different animal models (46). Specifically, we showed that efaA is highly upregulated, the N-ramp transporter mntH1 is downregulated, while mntH2 remains unchanged in the IE rabbit model. On the other hand, mntH2 is highly upregulated, and efaA and mntH1 remain unchanged in the CAUTI mouse model. In contrast, we show here that mntH1 is the most highly upregulated manganese transporter in infected wound beds, closely followed by mntH2, with efaA showing only a modest increase. This is also in stark contrast to what is observed in vitro, where efaA and mntH2 transcription greatly increase after manganese starvation but not mntH1 (62). While we lack information on how zinc transporters are regulated in vivo, we have shown previously that both adcABC and adcAII are repressed under high zinc conditions and strongly induced after treatment with the zinc-chelating agent TPEN [N,N,N′,N′-tetrakis(2-pyridinylmethyl)-1,2-ethanediamine] (45, 48, 49). Here, we show that transcription of both adcA and adcAII is strongly induced in wounds 1 day post-infection. However, while adcA transcription declines thereafter, adcAII expression continues to rise. These findings suggest that AdcAII becomes the predominant zinc-binding lipoprotein during the chronic stages of wound infection. In addition to the dual iron transporter efaA, the other four iron transporters, fitD, emtC, feoB, and fhuG, were strongly and consistently upregulated in infected wounds. Together, these results support the idea that these biometals are not readily accessible in the wound microenvironment while also underscoring the importance and complexity of trace metal homeostasis regulation in E. faecalis.
High-affinity iron, manganese, and zinc transporters promote E. faecalis virulence in systemic and localized infections.
To investigate the possible role of nutritional immunity, or a defect thereof, in chronic diabetic wounds, we compared the fitness profiles of iron, manganese, and zinc transport mutants in wounds of both B6 (non-diabetic) and DB (T2D) mice. We also monitored the temporal changes in the levels of major metal-sequestering host proteins, namely transferrin, lactoferrin, calprotectin, and psoriasin, in these wounds. Unlike S. agalactiae, in which deletion of either manganese or zinc transporters impaired its ability to colonize wounds of normoglycemic mice but not of diabetic mice (28), E. faecalis strains with impaired manganese (Δ3Mn) or zinc (Δ2Zn) uptake exhibited similar infectious potential in either B6 or DB mice. Several, not mutually exclusive, reasons may explain these apparently discrepant results. It is conceivable that E. faecalis has a higher demand for these metals than S. agalactiae. Although less likely given the similar number of high-affinity metal uptake systems shared by the two species (28, 44–46, 63, 64), it is also possible that S. agalactiae is simply better equipped to scavenge trace metals than E. faecalis. Furthermore, our study utilized a competitive model of infection, which would increase the competition for metals between strains and may have contributed to the Δ3Mn, Δ2Zn, and Δ5Fe strains having defects in colonization in the DB mice that might not be present in a single infection model. Lastly, differences in experimental models might also have contributed to these differences, particularly considering that we used C57Bl6J lepR^−/−^ mice, which model T2D, whereas the S. agalactiae study employed the streptozotocin-induced diabetes model, which more closely resembles T1D (28).
Although the Δ3Mn and Δ2Zn strains do not appear to benefit from the more permissive microenvironment of DB mice wounds, the Δ5Fe strain displayed a slight advantage, given its higher recovery rate from DB wounds at 3 days post-infection when compared to B6 mice wounds. In alignment with this observation, both total transferrin and lactoferrin levels were significantly lower in DB naïve skin tissue when compared to B6 mice. In the case of lactoferrin, this difference could also be observed on days 1 and 7 post-infection. Levels of the S100 proteins calprotectin and psoriasin in DB wounds were also significantly lower at 1 day post-infection. Although colonization defects were observed only in the Δ5Fe strain at a single post-infection time point, these findings suggest that normoglycemic mice may be better equipped to combat opportunistic wound infections than diabetic mice. While the results support the idea that diabetes can impair nutritional immunity, particularly the timely activation of iron-restricting mechanisms, comparisons between this study and the S. agalactiae study, despite differences in the diabetic mouse models used, also suggest that this effect and the types of metals involved may be pathogen-specific.
In conclusion, here we show that the ability to scavenge essential trace metals is critical to E. faecalis wound infection. We also show that, as expected, diabetic mice struggle to clear infections and are more susceptible to developing chronic wound infections compared to normoglycemic mice. While this study provides some evidence for impaired activation of nutritional immunity in the context of T2D, based on apparent differences in the surveillance and mobilization of metal-sequestering proteins at infection sites in B6 and DB mice, further research is needed to substantiate this possibility. In particular, studies that directly compare bioavailable metal pools in the skin and wound beds of diabetic versus non-diabetic animals are necessary. Additionally, hyperglycemia has been associated with aberrant protein glycosylation, which may further compromise host immune responses and metal-regulating pathways (65–67). Both transferrin and lactoferrin undergo N-linked glycosylation, and alterations in their glycosylation patterns have been associated with defects in protein stability, trafficking, and activity (68, 69). Prior research has shown that transferrin glycosylation patterns vary among individuals and are influenced by factors such as age, sex, and body weight (68). Furthermore, long-term T2D has been associated with the accumulation of advanced glycation end products (AGE), a heterogeneous group of molecules with irreversible, non-enzymatic glycation that can lead to aberrant protein folding, function, and aggregation (70). Building on this, it is plausible to speculate that the hyperglycemic environment characteristic of uncontrolled diabetes could promote aberrant glycosylation and/or AGE accumulation that negatively impacts iron-sequestering proteins like transferrin and lactoferrin, thereby compromising the timely activation of nutritional immunity. Beyond this work with E. faecalis and previous investigations involving S. agalactiae (28), further research is needed to explore the role of trace metal homeostasis in wound infections caused by other prominent pathogens, such as S. aureus and P. aeruginosa, as well as in polymicrobial infections in order to determine whether any potential defects in nutritional immunity contribute to enhanced infection in a pathogen-specific manner. In closing, this investigation expands our understanding of host-pathogen interactions in wound infections and identifies differences in metal availability that may inform new therapeutic strategies, particularly for at-risk populations.
MATERIALS AND METHODS
Bacterial strains and growth conditions
The strains used in this study are from −80°C freezer stocks previously generated by our lab (44–46). All strains were routinely grown aerobically in brain heart infusion (Difco Laboratories) at 37°C. The Δ2Zn strain was supplemented with 100 µM ZnSO_4_, and the Δ3Mn strain was grown with 100 µM MnSO_4_ to overcome growth defects observed in non-supplemented BHI (45, 46). Chemical and biological reagents were purchased from Sigma Aldrich unless stated otherwise.
Mouse wound excision model
Overnight cultures and starter cultures were obtained as previously (44), with minor modifications. Briefly, the bacterial inoculum was prepared by washing overnight-grown cells in 0.5 mM EDTA in PBS once and twice in trace metal grade PBS. For single-strain infection, cell pellets were concentrated to 1 × 10^10^ CFU mL^−1^ and stored on ice until use. For competitive index infections, cells were prepared as above, and all four strains were mixed in an equal ratio. Seven- to 10-week-old C57BL6J (B6) or 10-week-old B6.BKS(D)-Lepr^db^J (DB) mice (transgenic mice of the C57BL6J background with a mutation in the leptin receptor- stock #000697) were purchased from Jackson Laboratories (54). These mice have been evaluated by the Jackson Labs and are known to have elevated blood glucose (an average of 310 mg/mL) and HbA1c levels (average of 7.4 NGSP%—National Glycochemical Standardization Program) by 8 weeks of age. Animals were allowed to acclimate at our facilities for at least 3 days. On the day of the experiment, animals were anesthetized using isoflurane, their backs were shaved, and the excisional wound was created using a 6 mm biopsy punch. Infection of wounds and enumeration of bacterial burden were performed as previously described (42, 44), using tissue homogenate collected at 1, 3, or 7 days post-infection. For competitive index infections, the inoculum and wound homogenates were plated on selective TSA plates, and PCR was performed to assess the number of each mutant strain recovered using primers specific to iron (fitD), manganese (mntH2), and zinc (adcA) transport (Table 1). At least 100 colonies were screened per wound to determine the competitive index of each strain.
Quantitative real-time PCR
To obtain total RNA from BHI-grown cultures**,** E. faecalis OG1RF was first grown overnight in BHI. Starter culture was normalized to an OD_600_ of 0.5, inoculated at a ratio of 1:20 into fresh BHI, and incubated at 37°C for 1 h. Post-incubation, cells were harvested via centrifugation at 4,000 rpm for 10 min, washed once with PBS, and incubated with lysozyme (Sigma-Aldrich) (20 mg mL^-1^) for 30 min at 37°C. Next, cells were harvested by centrifugation and total RNA extracted using the PureLink RNA minikit (Invitrogen), according to the manufacturer’s instructions. To isolate E. faecalis total RNA from infected wounds, skin encompassing the wound site was excised and placed into 2 mL of RNAlater stabilization solution (Invitrogen) and incubated on ice for 1 h. The mouse skin was then transferred into 1 mL of TRIzol (Life Technologies), processed according to the manufacturer’s protocols using the Max Bacterial Enhancement Reagent (Invitrogen), with minor modifications. Briefly, the entire suspension, including the mouse tissue, was homogenized using a hand-held homogenizer (Benchmark Scientific), followed by three cycles of bead-beating (40 s each and chilled on ice for 2 min). Samples were centrifuged for 20 min at 12,000 rpm at 4°C to pellet cell debris, and the aqueous suspension was transferred to a new tube. To the homogenized aqueous suspension, chloroform (Sigma-Aldrich) was added, according to TRIzol (Life Technologies) Reagent’s standard protocol. The top layer (aqueous phase) containing RNA was transferred to a new tube, and total RNA was isolated by ethanol precipitation. Total RNA was pooled from at least three mice to generate one biological replicate (n = 3). RNA purification and removal of DNA were performed using a Turbo DNA-free kit (Thermo Fisher). Removal of mammalian RNA and isolation of bacterial RNA were performed using the MicrobEnrich kit (Invitrogen). The tissue-extracted bacterial RNA was quality checked using the RNA ScreenTape on a TapeStation instrument (Agilent Technologies) and quantified using Qubit at the University of Florida Interdisciplinary Center for Biotechnology Research Gene Expression and Genotyping Core facility. Synthesis of cDNA was performed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Quantitative real-time PCR (qRT-PCR) was performed using iTaq Universal SYBR supermix (BioRad) with the primers listed in Table 2. For quantification of transcript numbers, E. faecalis OG1RF gDNA was used as a template to generate standard curves. Relative quantification of gene transcripts was performed as previously described (71), using gyrA as the control gene.
Analysis of proteinaceous metal chelators in wounds by ELISA
Naïve tissues were collected on the day of wounding by collecting the intact skin tissue removed by the biopsy punch. Tissues (naïve or from infected wounds) were excised and mechanically homogenized in 1 mL metal-free PBS with 1 U of protease inhibitor (Halt Protease Inhibitor, ThermoScientific). The tissue was filtered using Flowmi 40 µm cell strainer tips (Sigma-Aldrich) to remove fibrous tissue. Samples were prepared following the instructions provided by each ELISA kit (all purchased from MyBioSource). For calprotectin (cat no. MBS7606640) and S100A7 (cat no. MBS763369) ELISAs, total protein content of the sample was determined using a BCA assay, and samples were diluted to 1 mg mL^−1^ total protein content. The dilution factor for each sample was recorded for downstream analysis. For aTF ELISA (cat no. MBS729302), undiluted samples were used, and for total transferrin ELISA (cat no. MBS2700560), a 1:10 (naïve skin tissue) or 1:100 (infected wound tissues) dilution of the sample was used. For lactoferrin ELISA (cat no. MBS269515), samples were diluted 1:10 (naïve skin tissue) or 1:50 (infected wound tissues). ELISAs were performed according to the manufacturer’s protocol with no deviation. The analyte available in undiluted samples was calculated based on dilution factors of the sample and normalized to the weight of tissue in grams.
Statistical analysis
All data sets were analyzed using GraphPad Prism 10 software. Data from multiple experiments conducted on non-consecutive days were collated. Data that falls within a Gaussian distribution was analyzed using either one-way or two-way ANOVA; however, a non-parametric test, such as the Kruskal-Wallis or Mann-Whitney, was used if there was non-normality of data, both with appropriate comparison tests. Two-sided P values were calculated, with P < 0.05 considered statistically significant.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Queen D, Harding K. 2024. Estimating the cost of wounds both nationally and regionally within the top 10 highest spenders. Int Wound J 21:e 14709. doi:10.1111/iwj.1470938379224 PMC 10830400 · doi ↗ · pubmed ↗
- 2Saeed S, Martins-Green M. 2024. Assessing animal models to study impaired and chronic wounds. Int J Mol Sci 25:3837. doi:10.3390/ijms 2507383738612647 PMC 11011258 · doi ↗ · pubmed ↗
- 3Landén NX, Li D, Ståhle M. 2016. Transition from inflammation to proliferation: a critical step during wound healing. Cell Mol Life Sci 73:3861–3885. doi:10.1007/s 00018-016-2268-027180275 PMC 5021733 · doi ↗ · pubmed ↗
- 4Leoni G, Neumann PA, Sumagin R, Denning TL, Nusrat A. 2015. Wound repair: role of immune-epithelial interactions. Mucosal Immunol 8:959–968. doi:10.1038/mi.2015.6326174765 PMC 4916915 · doi ↗ · pubmed ↗
- 5Norton P, Trus P, Wang F, Thornton MJ, Chang CY. 2024. Understanding and treating diabetic foot ulcers: Insights into the role of cutaneous microbiota and innovative therapies. Skin Health Dis 4:e 399. doi:10.1002/ski 2.39939104636 PMC 11297444 · doi ↗ · pubmed ↗
- 6Solomkin J, Egger M, de Jonge S, Latif A, Loke YK, Berenholtz S, Allegranzi B. 2018. World Health Organization responds to concerns about surgical site infection prevention recommendations. Anesthesiology 128:221–222. doi:10.1097/ALN.000000000000194129232227 · doi ↗ · pubmed ↗
- 7Armstrong DG, Tan TW, Boulton AJM, Bus SA. 2023. Diabetic foot ulcers: a review. JAMA 330:62–75. doi:10.1001/jama.2023.1057837395769 PMC 10723802 · doi ↗ · pubmed ↗
- 8Hurlow J, Bowler PG. 2022. Acute and chronic wound infections: microbiological, immunological, clinical and therapeutic distinctions. J Wound Care 31:436–445. doi:10.12968/jowc.2022.31.5.43635579319 · doi ↗ · pubmed ↗