BbCFEM7 plays an important role in the pathogenicity and gut microbial community formation in the co-infection of Beauveria bassiana with Metarhizium rileyi
Xu Zhang, Tianjiao Zhan, Qingqing Liu, Mengfei Xing, Wangjiao Yu, Bin Chen, Yuejin Peng

TL;DR
This study shows that the BbCFEM7 gene in a fungus boosts its ability to infect insects and changes the insect's gut microbes, offering new ways to control pests.
Contribution
The study reveals BbCFEM7's novel role in enhancing fungal pathogenicity and altering gut microbial communities during co-infection.
Findings
BbCFEM7 significantly increases the virulence of B. bassiana and reduces host survival time.
BbCFEM7 alters the host's gut microbial community, increasing Enterococcus and decreasing Thomasclavelia.
BbCFEM7 improves B. bassiana's stress tolerance and colonization of the host's hemolymph.
Abstract
This study investigated the effects of the BbCFEM7 gene in Beauveria bassiana on host pathogenicity and the gut microbial community during co-infection with Metarhizium rileyi in Spodoptera litura. Our results showed that BbCFEM7 significantly enhanced the virulence of B. bassiana and significantly reduced the host LT50 in the 1:0, 9:1, and 1:1 (Bb:Mr) groups. Sequencing of the gut microbial diversity showed that the presence of BbCFEM7 significantly altered the structure of the host gut microbial community, particularly in the 9:1 group. The presence of BbCFEM7 increased the relative abundance of Enterococcus and decreased that of Thomasclavelia. Furthermore, BbCFEM7 was found to play a pivotal role in enhancing B. bassiana’s tolerance to chemical stress and promoting its rapid colonization of the host’s hemolymph, thereby giving it an advantage over M. rileyi. These results suggest…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
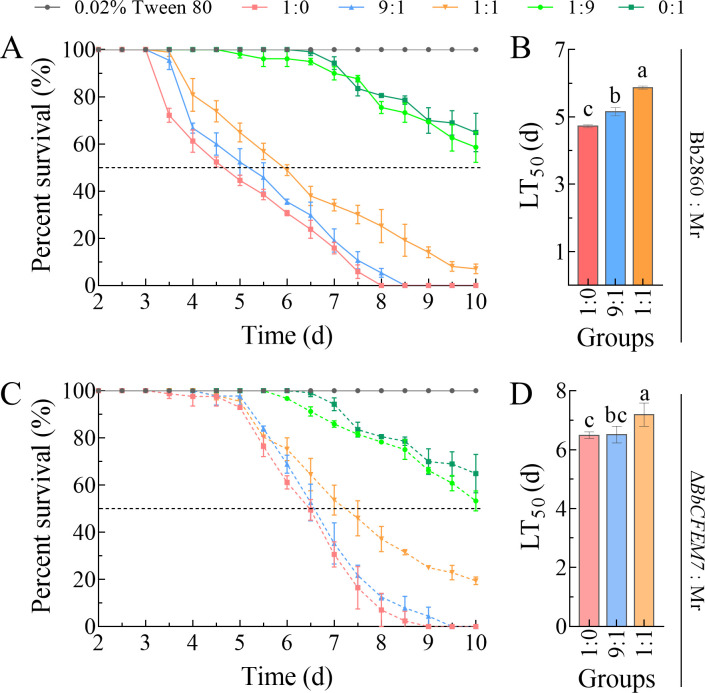 Fig 1
Fig 1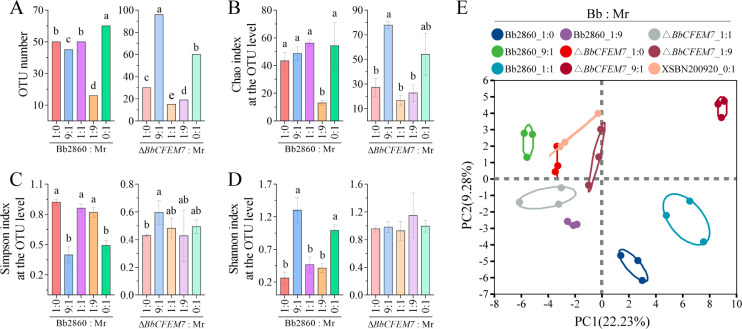 Fig 2
Fig 2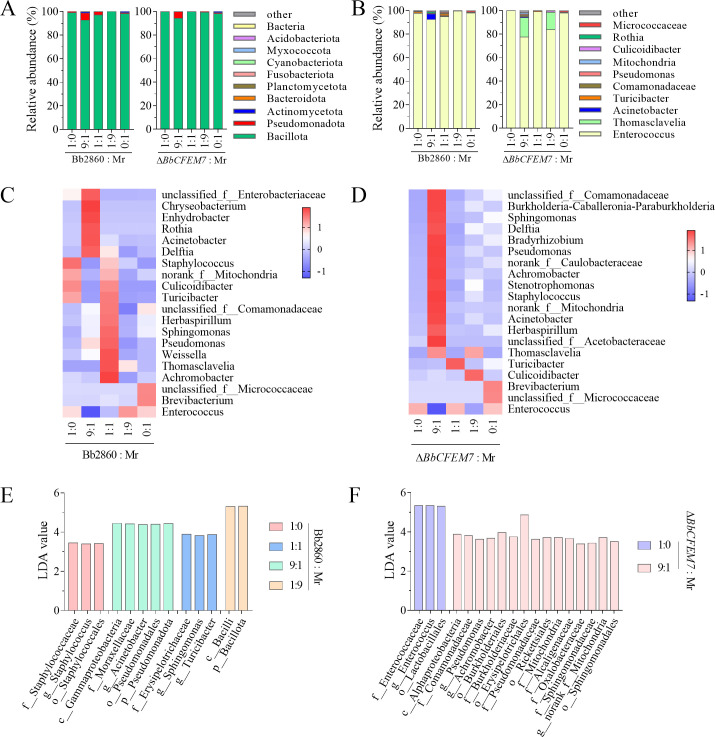 Fig 3
Fig 3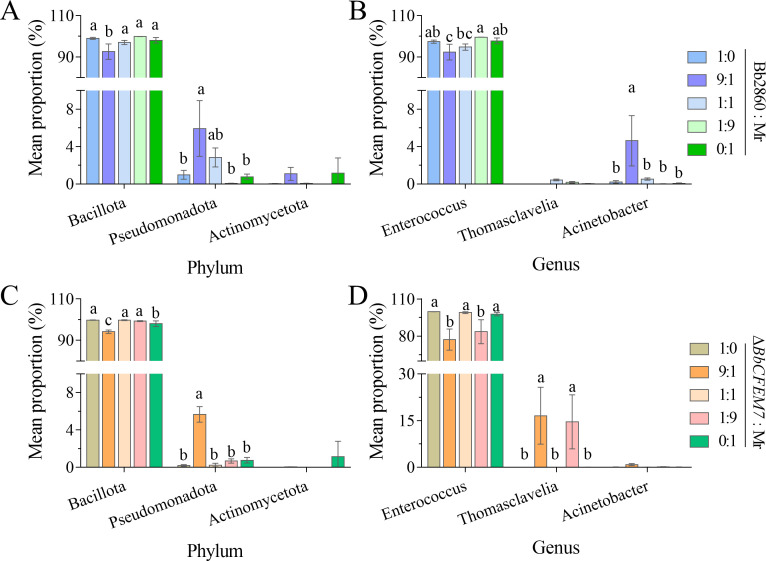 Fig 4
Fig 4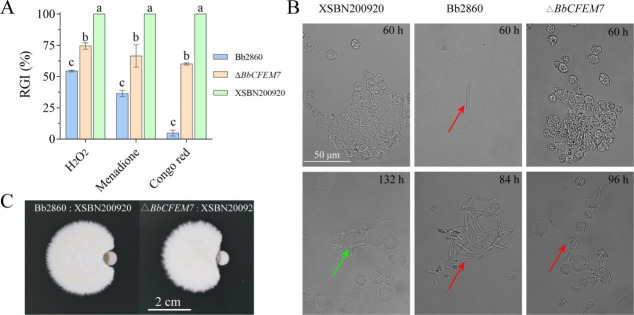 Fig 5
Fig 5- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Natural Science Foundation of Yunnan Province (Yunnan Natural Science Foundation)
- —Natural Science Foundation of Yunnan Province (Yunnan Natural Science Foundation)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Insect symbiosis and bacterial influences · Parasitic Diseases Research and Treatment
INTRODUCTION
Gut microbes are recognized as one of the protective barriers of arthropods, which play a key role in the physiology, ecology, and evolution of insects, as well as in nutrient metabolism, immune defense, behavioral regulation, adaptive evolution, competition with other pathogenic microorganisms, and niche differentiation of insects (1, 2). However, the competitive relationship between gut microbes and pathogens is complex. Due to the diversity of arthropods and pathogens, research in this field remains a serious challenge. Although Zaneveld et al. have summarized the changes of the host gut microbial community with the Anna Karenina principle (3), there are still many questions about the underlying mechanism. Some gut microbes can inhibit the invasion of pathogens (symbiotic microorganisms) (4, 5) or accelerate the death of the host (opportunistic pathogens) by occupying the gut niche, consuming nutrients, or secreting antimicrobial substances (6).
B. bassiana is a widely studied entomopathogenic fungus that has to go through an infection cycle before it infects and kills its host. The conidia of fungi attach to the surface of the insect body and form a bud tube through germination under suitable temperature and humidity conditions (7, 8). After passing through the body wall and entering the insect, the bud tube rapidly grows into the mycelium and spreads in the hemolymph of the insect. The mycelium absorbs nutrients from the insect, such as water, proteins, and sugars, while producing toxins (such as beauverine) (9) that cause physiological disorders in the insect. As the mycelium multiplies, the tissues inside the insect are destroyed and die. When the insect dies, the mycelium penetrates the surface of the insect again, forming an airborne mycelium and conidial stalk, releasing new conidia, which will enter the next cycle of infection under the right conditions (8).
In the process of proliferation in the host, the competition mechanism between the fungus and the host is complex and diverse. Insect-pathogenic fungi do not exist in isolation within natural environments. Their infection processes are profoundly influenced by the symbiotic microbial communities residing on and within the insect’s body surface. For example, when M. rileyi infects cotton bollworm, the host secretes antimicrobial peptides (AMPs) to counter the proliferation of pathogenic fungi in the hemolymph (10). The gut microbiome, as one of the defensive barriers of insects, also takes some measures to defend itself. For example, the composition and function of gut microbes can change dramatically, and some are able to defend against infection by activating the host immune response or directly inhibiting the growth of pathogenic bacteria. The presence of Acinetobacter baumannii makes the insects more resistant to Metarhizium anisopliae (11).
Common in fungal extracellular membrane (CFEM) domain proteins are unique to fungi, which play an important role in the process of obtaining iron for fungi. In entomopathogenetic fungi, Peng et al. systematically revealed the role of CFEM protein in iron acquisition in B. bassiana (12). These proteins are not only involved in obtaining iron from heme and trivalent iron ions but are also essential for the entire life cycle of B. bassiana, including saprophytic and pathogenic growth. In addition, they reported for the first time the synergistic division of labor and compensation of CFEM family genes at different levels of iron starvation. In Metarhizium robertsii, the CFEM domain protein (Mcdc9) acts as a contact elicitor to induce hygienic behavior in flies against fungal parasite infection (13). In Metarhizium acridum, the absence of MaCFEM1 leads to rapid proliferation of opportunistic bacteria in the insect host’s gut, resulting in rapid insect death (14). BbCFEM7 proved to be an important virulence factor. However, what role BbCFEM7 plays in host hemolymph proliferation is unknown.
In this study, different proportions of B. bassiana and M. rileyi were combined and verified by gut microbial diversity sequencing and related in vivo and in vitro experiments to investigate the role of two entomopathogenic fungi in infecting S. litura. The effects of CFEM domain proteins on fungal virulence and gut microbial community structure reveal the functions of CFEM proteins in fungal host and gut microbial interactions, providing a new perspective for understanding the ecological relationship between insect health and pathogenic bacterial infection. In addition, the results help optimize the use of entomopathogenic fungi, improve their application efficiency in pest control, and provide new research directions for insect microbiome and immunology research.
MATERIALS AND METHODS
Strains and media
M. rileyi strain XSBN200920 and B. bassiana 2860 (wild-type [WT] and ΔBbCFEM7 mutants) were utilized (12, 15) as previously described. For culturing, Sabouraud Maltose Agar medium plates (SMAY: 1% peptone, 1% yeast extract, 4% maltose, and 1.5% agarose) and Sabouraud Dextrose Agar medium plates (SDAY: 1% peptone, 1% yeast extract, 4% glucose, and 1.5% agar) were used. These fungi were incubated at 25°C under a 12-h light/12-h dark cycle in a culture chamber (RG-300, Xi’an Hengli Instrumentation Co., Ltd.). Conidial suspensions were prepared from fungi grown on SMAY and SDAY plates for 10 days.
Insect feeding
S. litura is raised indoors in an artificial climate room with a temperature set at 25°C, a photoperiod of 16 h light: 8 h darkness, and a relative humidity of 70% (16). The larvae are provided with fresh artificial feed every day. For adult rearing, the adults are placed in a 50 × 50 × 50 cm cage and provided with fresh honey water every other day.
Bioassay
To determine the virulence, fifth-instar S. litura larvae cultured over five generations were used for blood cavity injection experiments. A 1 × 10⁵ conidia/mL fungal conidial suspension was used to infect S. litura larvae, including M. rileyi and B. bassiana, according to the previous method. M. rileyi XSBN200920 was combined with WT or ΔBbCFEM7 mutants and set to five ratios (1:0, 9:1, 1:1, 1:9, and 0:1 [V:V]). Five microliters of spore suspension was injected into the blood cavity of each larva in each group. The experiment was repeated three times, with no fewer than 35 samples treated each time. After treatment, the larvae were raised indoors in an artificial climate. Mortality was monitored daily, and dead larvae were placed in Petri dishes to observe humidity and confirm whether they were infected by the tested strain. The dead insects were placed in Petri dishes for about 10 days and photographed (15).
Physiology experiments
A conidial suspension consisting of 2.5 μL 1 × 10^7^ conidia/mL was obtained from M. rileyi strain XSBN200920 and B. bassiana strains (WT and ΔBbCFEM7 mutants), respectively. The inoculum was added with the Czapek's agar (CZA) medium containing 50 μg/mL Congo red, 0.03 mM/L menadione, and 3 mM/L H_₂_O_₂_. The plate was cultured at 25°C, with 12 h of light/12 h of darkness for 7 days, and the colony diameter was measured. In the plate antagonism experiment of M. rileyi and B. bassiana, the SMAY plate was used as the vegetative growth medium for both strains. A fungal conidial suspension of 2.5 μL 1 × 10^7^ conidia/mL was inoculated on a plate, and photographs were taken on day 7. Conidia of the two strains were inoculated at a distance of 1.5 cm. The experiment was repeated three times with three parallel controls each time.
Morphological observation of fungi in hemolymph
To observe mycelium growth in insects, 5 microliters of 1 × 10⁵ conidia/mL conidial suspension was injected into the blood cavity of the host. The larvae were killed some time after injection (hours post-infection [HPI]), and their hemolymph was diluted 1:1 in a sterile anticoagulant (0.14 M NaCl, 0.1 M glucose, 25 mM sodium citrate, and 30 mM citric acid). The mycelium is then photographed under a microscope and viewed (17).
Gut microbial diversity sequencing
Referring to the previous method (15), a suspension of 1 × 10⁵ conidia/mL was injected into S. litura 5th-instar larvae. The injection procedures and specific operations are the same as those described in Section 2.3. On day 5 after injection, the intestines of the injected larvae were carefully dissected, and samples were sent to Majorbio Co. Ltd. (Shanghai) for paired terminal sequencing on the Illumina MiSeq PE300 platform (Illumina, San Diego, USA). Each fungus-treated sample was set up with three replicates, with each replicate containing six larvae. The Majorbio Cloud platform (https://cloud.majorbio.com) is used for gut microbiota bioinformatics analysis. Based on operational taxonomic units (OTUs), the dilution curve and α diversity index were generated, including the observed OTUs. Kruskal-Wallis rank sum test was used to compare the relative abundance of gut microbes at phylum and genus levels among the groups. α diversity-Chao richness estimates reflecting species richness and community diversity were calculated using QIIME2 (https://qiime2.org/). Principal component analysis (PCA) based on Bray-Curtis differences was used to assess the similarity of microbial communities in different samples using Vegan v2.5-3 packages. Linear discriminant analysis (LDA ≥ 2, P < 0.05) was used to identify species with significant differences in sample classification.
Data analysis
Data conforming to normal distribution and homogeneity of variance between two groups were compared using an unpaired t-test. Comparisons among multiple groups were conducted via a univariate analysis of variance (ANOVA) and bivariate ANOVA, followed by Tukey’s honest significance difference test (Tukey’s HSD). A P-value of < 0.05 was considered statistically significant. The software used is GraphPad Prism 7.00.
RESULTS
Virulence of BbCFEM7 in host co-infection of B. bassiana and M. rileyi
The virulence of B. bassiana (WT and ΔBbCFEM7 mutants) and M. rileyi, alone or in combination, was assessed by injection into larvae. Survival curves and half-lethal times (LT_50_) were analyzed (Fig. 1).The results showed that the presence of BbCFEM7 significantly affected the virulence of B. bassiana co-infected with M. rileyi. Specifically, compared with the ΔBbCFEM7 mutant, the survival curve of larvae infected with B. bassiana WT decreased more sharply (Fig. 1A and C), and the LT_50_ was significantly smaller (Fig. 1B and D). This observation was further supported by LT_50_ values, with WT strains dying faster than mutant strains in all test ratios (1:0, 9:1, and 1:1) (P = 0.0000008, P = 0.0000132, and P = 0.0000173). By comparing the larval survival rate and LT_50_ under different combination ratios, the role of the BbCFEM7 gene in the co-infection of B. bassiana and M. rileyi and its influence on the pathogenicity of the host can be evaluated.
Virulence of BbCFEM7 in host co-infection of B. bassiana and M. rileyi. (A and B): Larva survival curves and LT50 of B. bassiana 2860 (WT) and M. rileyi XSBN200920 conidia suspensions at 1 × 105 conidia /mL under five groups of infection, respectively. Larva survival curves and LT50 of (C and D): the ΔBbCFEM7 mutant and M. rileyi XSBN200920 conidia suspension under 1 × 105 conidia /mL infection in five groups, respectively. Categories include Bb:Mr (1:0, 9:1, 1:1, 1:9, and 0:1). Tukey’s HSD test: P < 0.05. Error bars represent standard deviation.
α and β analyses of host gut microbial diversity by BbCFEM7 during co-infection
The diversity of gut microbiota was assessed during co-infection with B. bassiana (both WT and ΔBbCFEM7 mutant) and M. rileyi (Fig. 2). Changes in the gut microbial communities were quantified using α-diversity indices, which include OTU), species richness (Chao index), evenness (Simpson index), and diversity (Shannon index) (Fig. 2A through D). The results indicated significant differences between the WT and ΔBbCFEM7 mutant groups, demonstrating that BbCFEM7 influences both the abundance and evenness of the gut microbiota. For instance, larvae infected with the WT strain exhibited a higher Chao index compared to those infected with the ΔBbCFEM7 mutant, suggesting enhanced species richness due to the presence of BbCFEM7 (Fig. 2B). Additionally, a β-diversity analysis, employing principal coordinate analysis (PCoA) based on Bray-Curtis dissimilarity, showed distinct clustering patterns of the gut microbiome between the WT and mutant groups (Fig. 2E). This analysis confirms that BbCFEM7 significantly modifies the overall structure of the gut microbiome during co-infection.
α and β analyses of host gut microbial diversity by BbCFEM7 during co-infection with B. bassiana and M. rileyi. (A–D): OTU index (A), Chao Index (B), Simpson Index (C), and Shannon Index (D). a, b, and c denote statistically significant differences between groups. (E) PCoA of the gut microbial β diversity index following co-infection with WT and ΔBbCFEM7 mutants of B. bassiana, after treatment with a 1 × 105 conidia/mL suspension for 3 days. Statistical significance tested using Tukey’s HSD test: P < 0.05. Error bars represent standard deviation.
Effects of BbCFEM7 on gut microbial composition during co-infection
The gut microbial community structure at both phylum and genus levels was analyzed in depth during co-infection with B. bassiana (both WT and ΔBbCFEM7 mutants) and M. rileyi. The top 10 microbial taxa at different infection rates were identified and compared at the phylum level (Fig. 3A) and genus level (Fig. 3B). At the phylum level, the dominant bacteria in both the WT and M. rileyi co-infection group and the ΔBbCFEM7 mutant and M. rileyi co-infection group were Pseudomonadota and Bacillota, among others, with no significant differences between the two groups. At the genus level, Enterococcus was the dominant bacterium in the WT and M. rileyi co-infection group. In the 9:1 and 1:9 (Bb2860:Mr) groups, the abundance of Thomasclavelia was significantly increased. Compared to the WT and M. rileyi co-infected groups, significant changes in the composition of various gut bacteria were observed in larvae co-infected with the ΔBbCFEM7 mutant and M. rileyi across five groups. For instance, in the ΔBbCFEM7 mutant-only infection group (1:0), populations of Turicibacter and Culicoidibacter were significantly reduced. In the 9:1 (Bb:Mr) group, the population of Sphingomonas was significantly increased, whereas that of Brevibacterium was significantly decreased. Heat maps of the top 20 microbial taxa (Fig. 3C and D) further highlight the pattern of abundance differences between the WT and mutant groups. Additionally, the microbial abundance profiles (Fig. 3E and F) revealed distinct patterns of microbial community aggregation during co-infection, with significant variations in the gut characteristic microbes of insect hosts infected by B. bassiana and M. rileyi at different ratios. For example, in the 9:1 (Bb:Mr) group, the ΔBbCFEM7 mutant and M. rileyi co-infected hosts exhibited as many as 15 insect gut microbes compared to the five characteristic microbes in the WT and M. rileyi co-infected hosts. The specific effects of BbCFEM7 on the host gut microbial community during co-infection were elucidated by analyzing the changes in gut microbial species and abundance under various proportional combinations.
Effects of BbCFEM7 on gut microbial diversity when B. bassiana and M. rileyi co-infected S. litura larvae. (A): Distribution of the top 10 microbial species in the gut of S. litura larvae infected by five proportions of 1 × 105 conidia/mL M. rileyi and conidial suspensions of B. bassiana2860 (WT) or ΔBbCFEM7 mutant, respectively, at the phylum level. (B): Distribution of the top 10 microbial species in the gut of S. litura larvae infected with conidial suspensions of B. bassiana and the ΔBbCFEM7 mutant, respectively, at the genus level. (C and D): Heat maps of the relative abundance of the top 20 microorganisms in the gut of insects infected by M. rileyi and B. bassiana2860 (WT) in five different ratios and those infected with M. rileyi and the ΔBbCFEM7 mutant under five different ratios, respectively. (E and F): Analysis of the gut microbial abundance spectra of insects infected with different combinations of M. rileyi, B. bassiana, and the ΔBbCFEM7 mutant.
Changes in gut microbiota of S. litura infected by B. bassiana and M. rileyi
At the phylum and genus levels, significant changes were observed in the gut microbiome of larvae co-infected with B. bassiana (both WT and ΔBbCFEM7 mutants) and M. rileyi (Fig. 4). The results indicated that infection with Bacillus cocciformis by WT strains led to significant changes in the gut microbial community structure compared with the ΔBbCFEM7 mutant. In the groups with 9:1 and 1:9 ratios, the population of Enterococcus, when co-infected by the ΔBbCFEM7 mutant and M. rileyi, decreased significantly compared with the groups co-infected with the WT and M. rileyi. Additionally, the population of Thomasclavelia increased significantly.
Changes in the gut flora of S. litura infected by B. bassiana and M. rileyi. (A and B): Changes in the host gut flora of M. rileyi and B. bassiana2860 (WT) at the phylum and genus levels, respectively. (C and D): The gut flora changes in hosts infected with M. rileyi and the ΔBbCFEM7 mutant at the phylum and genus levels, respectively. Statistical significance was tested using Tukey’s HSD test: P < 0.05. The error bars represent the standard deviation.
Competition between B. bassiana and M. rileyi in vitro and in vivo
The competitive relationship between B. bassiana (WT and ΔBbCFEM7 mutants) and M. rileyi has been confirmed both in vitro and in vivo. In vitro competitive analysis using chemical stress media (Fig. 5A) showed that the ΔBbCFEM7 mutants were significantly less resistant to stress agents such as Congo red, H_2_O_2_, and menadione compared to the WT strains of B. bassiana. In contrast, M. rileyi exhibited almost complete loss of resistance to these three stressors at the same concentrations. In vivo competitive analysis demonstrated different growth patterns of fungal spores in the hemolymph of caterpillars (Fig. 5B). Compared to the WT, the ΔBbCFEM7 mutant strain colonized the hemolymph more slowly, with M. rileyi displaying the slowest proliferation rate in the host hemolymph. Additionally, antagonistic experiments on SDAY plates (Fig. 5C) showed that B. bassiana 2860 (WT) was more effective at inhibiting the growth of M. rileyi compared to the ΔBbCFEM7 mutant, with significant differences observed in colony growth and inhibition zones.
In vitro and in vivo competition between B. bassiana and M. rileyi. (A): Growth status of B. bassiana 2860 (WT), ΔBbCFEM7 mutant, and M. rileyi XSBN200920 on various chemical stress media on day 7. Stressors include Congo red, H2O2, and menadione. Differences were statistically significant as determined by Tukey’s HSD test: P < 0.05. Error bars represent standard deviation. (B): Growth of fungal conidia in the hemocoel of S. litura larvae was assessed at various time intervals following the injection of a suspension containing 1 × 105 conidia/mL of either B. bassiana 2860 (WT), the ΔBbCFEM7 mutant, or M. rileyi XSBN200920. The red arrow indicates B. bassiana in the blood cavity, and the green arrow indicates M. rileyi in the hemolymph. (C): Growth confrontation of M. rileyi with B. bassiana 2860 (WT) or ΔBbCFEM7 mutant on an SDAY plate. The starting point of colony growth is 1.5 cm away. Tukey’s HSD test: P < 0.05. Error bars represent standard deviation.
DISCUSSION
This study analyzed the function of BbCFEM7 in B. bassiana and its influence on host pathogenicity during co-infection with M. rileyi in S. litura. The results demonstrated that the presence of BbCFEM7 significantly enhanced the virulence of B. bassiana (Fig. 1). Specifically, the WT exhibited a shorter LT_50_ compared to the ΔBbCFEM7 mutant across all tested ratios. To further validate the role of BbCFEM7 in natural infection pathways, we performed virulence assays using cuticle immersion treatments (Fig. S1). The LT_50_ values were 7.13 days for the WT strain and 6.97 days for the complementation mutant, whereas the ΔBbCFEM7 mutant did not achieve 50% cumulative mortality, making LT_50_ calculation unfeasible. This finding directly demonstrates that BbCFEM7 is crucial in natural infection pathways. It corroborates previous studies that suggest that BbCFEM7 plays a key role in iron acquisition and the fungal life cycle (12). Given the involvement of CFEM domain proteins in iron ion uptake and utilization and the importance of iron metabolism in fungal growth and pathogenicity, it is plausible that BbCFEM7 enhances the pathogenicity of B. bassiana by regulating iron metabolic pathways. Notably, spore germination assays showed that germination rates for the WT (65.89%) and the complementation mutant (65.65%) were significantly higher than those of the ΔBbCFEM7 mutant (43.60%) (Fig. S1). This suggests that BbCFEM7 may also affect conidial germination capacity, thereby providing new insights into its pathogenic mechanism (12).
The gut microbiota of insects plays a crucial role in their host defense system, combating pathogen invasion through multiple mechanisms (18). These include enhancing immune responses, participating in pathogen clearance, metabolic detoxification, and maintaining intestinal homeostasis. Research has shown that in the hindgut of termites, Cononympha leidyi and other symbiotic protozoa participate in the nitrogen cycle and inhibit infections by entomopathogenic fungi through chitin degradation (19). In Bactrocera dorsalis, gut microorganisms enhance the expression of AMPs by regulating the host’s Imd signaling pathway, thereby resisting the invasion of foreign pathogens (20). Some gut bacteria secrete AMPs or organic acids that inhibit the reproduction of pathogenic bacteria (21). Additionally, certain gut bacteria can regulate the host’s immune signaling pathways by secreting metabolites, thus enhancing the host’s immune response (22). These mechanisms not only assist insects in coping with pathogens and toxins in their environment but also have significant implications for their health and survival. In response, insect pathogenic fungi have evolved complex genomic strategies to counteract these host defenses, including targeting the humoral immunity of insects (23) and producing antibacterial substances (24) to enhance their survival and pathogenicity within the host. In M. robertsii, COA1 conceals cell wall antigenic structures (25), and Fkp1 targets the host enzyme cathepsin CtsK1 (26), facilitating immune evasion. In B. bassiana, oosporein enhances fungal survival by suppressing the production of AMPs and the activity of immune cells (27, 28).
This study further examined the influence of BbCFEM7 on the gut microbiota of S. litura during co-infection, as assessed by microbial diversity sequencing (Fig. 3 and 4). In the analysis of gut microbiomes, PCA two-dimensional plots capture only a portion of the total variation in the data. In contrast, LDA models, by integrating information across all dimensions, reveal differences between treatments more sensitively. As a result, the findings from these two analytical approaches are not entirely consistent. The presence of BbCFEM7 significantly altered the composition and abundance of gut microbes. Notably, infection with the WT strain significantly increased the relative abundance of Enterococcus in the insects, a phenomenon absent in the ΔBbCFEM7 mutant infection group. This suggests that BbCFEM7 can reshape the gut microbiome, an effect potentially closely linked to fungal virulence. This linkage has been demonstrated in locust studies where a deficiency in CFEM domain proteins leads to opportunistic pathogen overgrowth and accelerated host mortality (14). Therefore, the gut microbiota disruption induced by the ΔBbCFEM7 mutant may underlie its reduced virulence, supporting the dual role of BbCFEM7 in regulating microbial dynamics and pathogenicity. Additionally, other CFEM proteins influence host behavior and immunity; for example, in M. robertsii, Mcdc9 triggers antifungal behavioral defenses in flies (13). The reduced virulence of ΔBbCFEM7 may thus also reflect enhanced host immune responses. Future studies should aim to clarify the molecular mechanisms by which BbCFEM7 mediates fungus-host interactions.
This study also explores the competitive relationship between B. bassiana and M. rileyi, particularly in the presence or absence of BbCFEM7 (Fig. 5). In vitro experiments demonstrated that the WT B. bassiana exhibited greater tolerance under chemical stress conditions, while the ΔBbCFEM7 mutant displayed a significant disadvantage. In vivo experiments confirmed that the WT strain colonized insect hemolymph more rapidly than the mutant. These findings suggest that BbCFEM7 may confer competitive advantages to B. bassiana against pathogens like M. rileyi by enhancing the fungus’s stress tolerance.
Competitive relationships between fungi are common in nature, particularly within insect hosts. These interactions not only directly influence fungal growth and reproduction but may also affect the host’s health status and immune response. For example, certain gut microbes can inhibit the growth of pathogenic bacteria by secreting antimicrobial substances or by activating the host’s immune system (1). In this study, the presence of BbCFEM7 significantly enhanced the competitive ability of B. bassiana, likely due to its role in iron acquisition and stress resistance. Future research should aim to elucidate the specific molecular mechanisms through which BbCFEM7 participates in fungal competition and systematically assess the potential impact of this competition on host physiology and health. Additionally, although the effects of BbCFEM7 on fungal competition have been observed in vitro and in vivo, further studies are needed to explore how this competition manifests in the natural environment.
Conclusion
In conclusion, this study reveals the importance of BbCFEM7 in host-pathogen interactions by comprehensively analyzing its role in the pathogenicity of entomopathogenic fungi, gut microbial community structure, and fungal competiftion. These findings provide a new theoretical basis for the ecological study of insect health and pathogen infection and provide a new method for biological control of pests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wei G, Lai Y, Wang G, Chen H, Li F, Wang S. 2017. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc Natl Acad Sci USA 114:5994–5999. doi:10.1073/pnas.170354611428533370 PMC 5468619 · doi ↗ · pubmed ↗
- 2Raymann K, Shaffer Z, Moran NA. 2017. Antibiotic exposure perturbs the gut microbiota and elevates mortality in honeybees. P Lo S Biol 15:e 2001861. doi:10.1371/journal.pbio.200186128291793 PMC 5349420 · doi ↗ · pubmed ↗
- 3Zaneveld JR, Mc Minds R, Vega Thurber R. 2017. Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat Microbiol 2:17121. doi:10.1038/nmicrobiol.2017.12128836573 · doi ↗ · pubmed ↗
- 4Zhang JF, Li F, Zhong HY, Chen JM. 2022. Nilaparvata lugens (Hemiptera: Delphacidae) midgut microbial community responses to exposure to Metarhizium flavoviride (Sordariomycetes: Hypocreales). J Entomol Sci 57:587–602. doi:10.18474/JES 22-11 · doi ↗
- 5Zhang F, Sun XX, Zhang XC, Zhang S, Lu J, Xia YM, Huang YH, Wang XJ. 2018. The interactions between gut microbiota and entomopathogenic fungi: a potential approach for biological control of Blattella germanica (L.). Pest Manag Sci 74:438–447. doi:10.1002/ps.472628888066 · doi ↗ · pubmed ↗
- 6Peng Y, Wen S, Wang G, Zhang X, Di T, Du G, Chen B, Zhang L. 2023. Reconstruction of gut bacteria in Spodoptera frugiperda infected by Beauveria bassiana affects the survival of host pest. J Fungi (Basel) 9:906. doi:10.3390/jof 909090637755014 PMC 10532432 · doi ↗ · pubmed ↗
- 7Shah PA, Pell JK. 2003. Entomopathogenic fungi as biological control agents. Appl Microbiol Biotechnol 61:413–423. doi:10.1007/s 00253-003-1240-812764556 · doi ↗ · pubmed ↗
- 8Wanchoo A, Lewis MW, Keyhani NO. 2009. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria bassiana. Microbiology (Reading) 155:3121–3133. doi:10.1099/mic.0.029157-019608611 · doi ↗ · pubmed ↗