Prevalence and genetic diversity of intestinal parasites in Xilingol League herbivores, Inner Mongolia, Northern China
Ziran Mo, Jingwei Quan, Jiahao Dao, Xiaoping Luo, Penglong Wang, Yurun Miao, Lei Xiu, Gaowa Gong, Jian Li, Wenbin Yang, Junyan Li, Wei Hu

TL;DR
This study examines the prevalence and genetic diversity of intestinal parasites in herbivores in Inner Mongolia, finding high infection rates and zoonotic risks.
Contribution
The study provides the first comprehensive assessment of intestinal parasite co-infections and genetic diversity in Xilingol League herbivores.
Findings
The overall prevalence of intestinal parasites was 80.5% among herbivores, with a co-infection rate of 47.8%.
Genetic analysis revealed dominant zoonotic lineages of Giardia, Cryptosporidium, and Enterocytozoon.
Eimeria spp. infection was significantly associated with co-infections with other parasites.
Abstract
Intestinal parasites are common infectious agents that substantially impact the health and productivity of livestock, with several species posing zoonotic risks. In this study, 205 fecal samples were collected from cattle, sheep, and goats in Xilingol League. Microscopic examination was performed to detect Eimeria spp., nematodes, and trematodes, whereas nested polymerase chain reaction assays were used to identify Giardia duodenalis, Cryptosporidium spp., and Enterocytozoon bieneusi. The results showed that the overall prevalence of intestinal parasites was 80.5% (165/205) among herbivores, with a co-infection rate of 47.8% (98/205). Across host species, the prevalence rates were 92.8% (126/139) in sheep, 75.0% (12/16) in goats, and 52.0% (26/50) in cattle. The prevalences of Eimeria spp., nematodes, trematodes, G. duodenalis, Cryptosporidium spp., and E. bieneusi were 59.5%, 16.1%,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1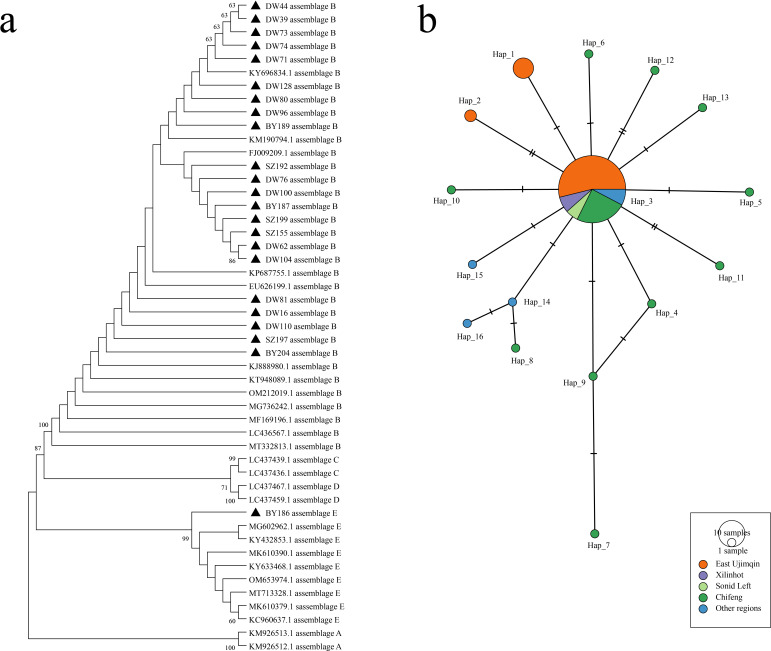 Fig 2
Fig 2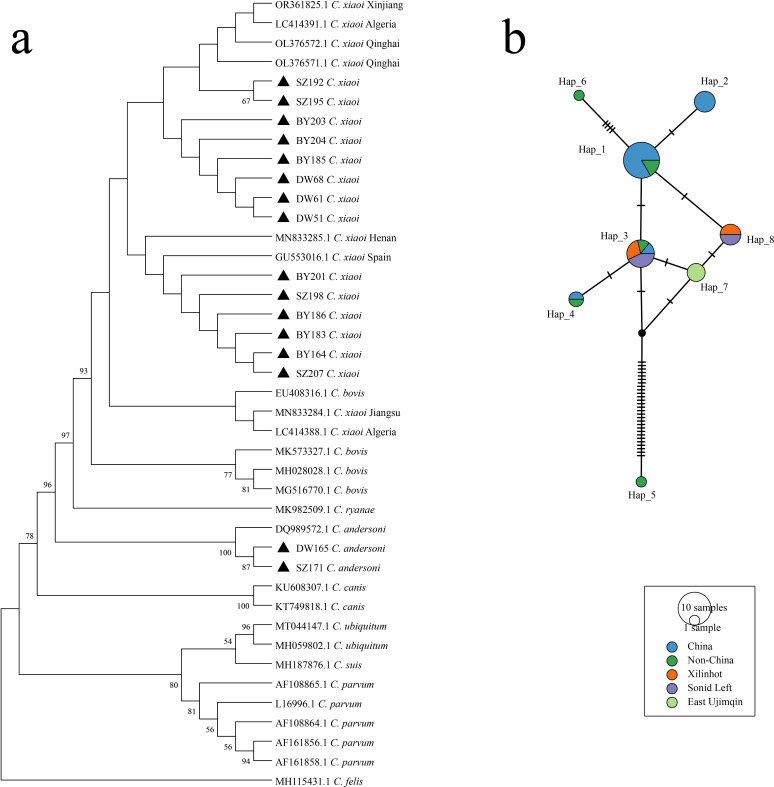 Fig 3
Fig 3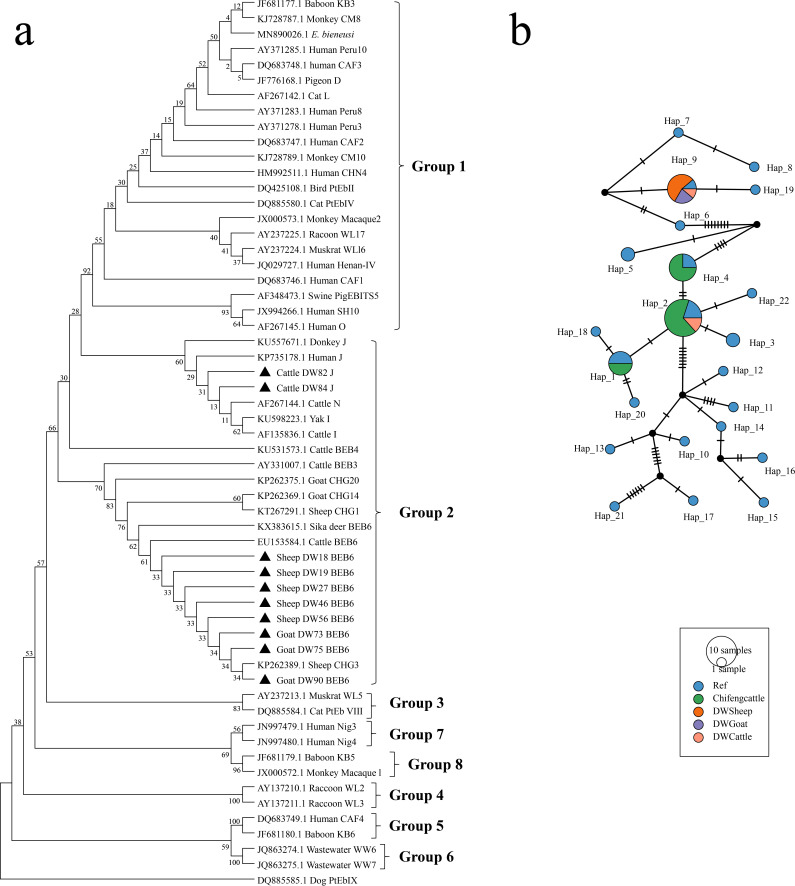 Fig 4
Fig 4| Region | Host | No. | No. positive for | No. positive for | No. positive for | No. positive for | No. positive for | No. positive for | Assemblage | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sonid Left | Cattle | 12 | 2 (16.7) | 0 (0) | 4 (33.3) | 1 (8.3) | 0 (0) | – | 1 (8.3) | B (1) | |
| Goat | 5 | 0 (0) | 1 (20.0) | 1 (20.0) | 2 (40.0) | 0 (0) | – | 1 (20.0) | B (1) | ||
| Sheep | 5 | 0 (0) | 2 (40.0) | 2 (40.0) | 2 (40.0) | 0 (0) | – | 2 (40.0) | B (2) | ||
| Xilinhot | Sheep | 43 | 29 (67.4) | 20 (46.5) | 11 (25.6) | 7 (16.3) | 0 (0) | – | 6 (14.0) | E (1), B (5) | |
| East Ujimqin | Cattle | 38 | 3 (7.9) | 0 (0) | 17 (44.7) | 1 (2.6) | 3 (7.9) | J (2), BEB6 (1) | 3 (7.9) | B (3) | |
| Goat | 11 | 8 (72.7) | 0 (0) | 4 (36.4) | 0 (0) |
| 2 (18.2) | BEB6 (2) | 6 (54.6) | B (6) | |
| Sheep | 91 | 80 (87.9) | 10 (11.0) | 30 (33.0) | 3 (3.3) | 5 (5.5) | BEB6 (5) | 34 (37.4) | B (34) | ||
| Subtotal | 205 | 122 (59.5) | 33 (16.1) | 69 (33.7) | 16 (7.8) | 10 (4.9) | J (2), BEB6 (8) | 53(25.9) | B (52), E (1) |
| Parasite species | Gene locus | Primer sequences(5′→3′) | Fragment size/bp | Annealing temperature/°C |
|---|---|---|---|---|
|
| F1: GAC ATA TCA TTC AAG TTT CTG ACC R1: CTG AAG GAG TAA GGA ACA ACC F2: CCT ATC AGC TTT AGA CGG TAG G R2: TCT AAG AAT TTC ACC TCT GAC TG | 587 | 58 | |
|
|
| F1: GGT CAT AGG GAT GAA GAG R1: TTC GAG TTC TTT CGC GCT C F2: GCT CTG AAT ATC TAT GGC T R2: ATC GCC GAC GGA TCC AAG TG | 392 | 57 |
|
|
| F1: AAG CCC GAC GAC CTC ACC CGC AGT GC R1: GAG GCC GCC CTG GAT CTT CGA GAC GAC F2: GAA CGA ACG AGA TCG AGG TCC G R2: CTC GAC GAG CTT CGT GTT | 511 | 65 |
- —Department of Science and Technology of Inner Mongolia Autonomous Region
- —Inner Mongolia Universityhttp://dx.doi.org/10.13039/501100003850
- —Department of Science and Technology of Inner Mongolia Autonomous Region
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Helminth infection and control · Parasites and Host Interactions
INTRODUCTION
Intestinal parasitic infections are a major global public health challenge, affecting over 2 billion people, particularly in low-resource regions lacking adequate sanitation (1, 2). These infections, caused by protozoa (e.g., Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi, Eimeria spp.) and helminths (e.g., Ascaris spp., Taenia spp., Trichuris spp.), are primarily transmitted via the fecal-oral route and are associated with gastrointestinal disorders, malnutrition, and potentially fatal outcomes in immunocompromised individuals (3–7). In livestock, intestinal parasites impair growth performance, reduce product quality, and impose substantial economic losses, while infected animals serve as reservoirs that contaminate the environment and facilitate zoonotic transmission (8–11). Therefore, systematic epidemiological investigations of intestinal parasites in livestock are essential to elucidate infection dynamics, transmission patterns, and population genetic diversity, thereby supporting sustainable animal husbandry, ensuring food safety, and protecting public health.
Intestinal parasitic infections are widespread in livestock worldwide, with infection prevalence often exceeding 70% in some regions (12–17). Inner Mongolia, particularly the Xilingol League, represents one of China’s most important pastoral areas, supporting more than 10 million grazing animals across approximately 180,000 km^2^ of grassland (18–20). While Cryptosporidium spp. and G. duodenalis infections in cattle have been reported (21, 22), comprehensive epidemiological data on intestinal parasites, especially multi-species infections, remain limited in this region. To date, only one study has documented nematode infections in local sheep (23), highlighting the lack of baseline information on intestinal parasite diversity and transmission risks in Xilingol’s grazing systems.
Given the region’s open grazing environment and constant exposure of livestock to environmental sources of infection, systematic surveillance is essential for assessing infection pressure and transmission potential. Intestinal parasitic infections frequently occur as co-infections involving multiple parasite species, which can exacerbate pathological outcomes and increase transmission potential (24, 25). However, most existing regional surveys have focused on single-parasite detection or overall prevalence, providing limited insight into co-infection dynamics and interspecies interactions. Therefore, comprehensive and systematic investigations are needed to clarify the occurrence, interactions, and epidemiological impacts of intestinal parasite co-infections in livestock populations.
To address this knowledge gap, a systematic investigation of herbivores from three major livestock areas in the Xilingol League was conducted, using both microscopic examination and nested PCR to detect Eimeria spp., nematodes, trematodes, G. duodenalis, and Cryptosporidium spp., and E. bieneusi. This study establishes baseline data on parasite prevalence and genetic diversity in the region and provides essential evidence for developing targeted control strategies to improve livestock productivity, ensure food safety, and reduce zoonotic risks.
RESULTS
Epidemiological characteristics of Eimeria spp., nematodes, and trematodes
Microscopic examination was performed to investigate intestinal parasites in livestock across multiple sites within the Xilingol League. Among the detected parasites, Eimeria spp. oocysts were the most frequently observed. The overall infection rate of Eimeria spp. was 59.5% (122/205) (Fig. 1; Table 1), with variation observed among host species and sampling regions. Across regions, infection rates were highest in East Ujimqin County (65.0%, 91/140) and Xilinhot (67.4%, 29/43), both significantly higher than in Sonid Left Banner (9.1%, 2/22) (DW vs SZ: odds ratio [OR] = 18.57, 95% confidence interval [CI]: 4.41–81.86, P < 0.05; BY vs SZ: OR = 20.71, 95% CI: 4.34–94.79, P < 0.05). Host-specific analysis showed that sheep had a higher infection rate (78.4%) compared to goats (50.0%; OR = 3.63, 95% CI: 1.20–10.91, P < 0.05) and cattle (10.0%; OR = 32.7, 95% CI: 11.82–79.85, P < 0.05). Regional variation was also evident within the same host species, with infection in sheep from East Ujimqin County (87.9%) significantly higher than in Xilinhot (67.4%) (OR = 3.51, 95% CI: 1.37–8.58, P < 0.05). Overall, Eimeria spp. infection exhibited distinct regional and host-associated differences, with the highest prevalence recorded in sheep and in samples collected from East Ujimqin County and Xilinhot.
Oocysts or eggs of the infected gastrointestinal parasites ([a] Eimeria spp. oocyst; [b] Dicrocoelium spp. egg; [c] Paramphistomum spp. egg; [d] Haemonchus spp. egg).
Microscopic examination was performed to investigate nematode and trematode infections in livestock across multiple sites within this region. Nematode and trematode eggs were detected in 16.1% (33/205) and 33.7% (69/205) of fecal samples, respectively (Fig. 1; Table 1). The prevalence of nematode infections showed considerable regional variation, with the highest infection rate in Xilinhot (46.5%), significantly higher than that in East Ujimqin County (7.1%; OR = 10.29, 95% CI: 4.361–25.49, P < 0.05) and Sonid Left Banner (13.6%; OR = 12.03, 95% CI: 2.72–43.29, P < 0.05). By host species, the prevalence of nematodes was higher in sheep (28.8%) than in goats (6.3%) and cattle (0%), although no statistically significant difference was detected among hosts. Within the sheep population, infection in Xilinhot was significantly higher than in East Ujimqin County (OR = 6.41, 95% CI: 2.66–16.20, P < 0.05). The prevalence of trematodes was relatively consistent among regions, with East Ujimqin County (36.4%), Sonid Left Banner (31.8%), and Xilinhot (25.6%), and no significant regional variation was detected. Among the host species, cattle exhibited the highest prevalence (42.0%), followed by goats (31.3%) and sheep (30.9%), though these differences were not statistically significant. Overall, nematode infections exhibited pronounced regional and host-related variation, whereas trematode infections remained relatively consistent across regions and host species.
Collectively, gastrointestinal parasite infections in livestock from Xilingol League demonstrated distinct epidemiological patterns, with overall prevalence rates of 59.5% for Eimeria spp., 33.7% for trematodes, and 16.1% for nematodes.
Prevalence, phylogenetic analysis, and haplotype characterization of G. duodenalis
Molecular detection was performed to investigate G. duodenalis infection and genetic diversity in livestock across multiple sites within the Xilingol League. The overall prevalence of G. duodenalis was 25.9% (53/205) (Table 1). Across regions, infection rates were highest in East Ujimqin County (30.7%), followed by Sonid Left Banner (18.2%) and Xilinhot (14.0%). Host-specific analysis showed that goats (54.6%) and sheep (37.4%) had a significantly higher infection risk than cattle (7.9%) (goat vs cattle: OR = 8.94, 95% CI: 1.99–30.49, P < 0.05; sheep vs cattle: OR = 4.98, 95% CI: 1.81–13.54, P < 0.05). Within the sheep population, the infection rate in East Ujimqin County (37.4%) was significantly higher than in Xilinhot (14.0%) (OR = 3.69, 95% CI: 1.49–9.30, P < 0.05).
Phylogenetic analysis based on the β-giardin (bg) gene locus demonstrated that the G. duodenalis sequences obtained in this study clustered into distinct clades corresponding to assemblages B and E, which showed high sequence similarity to strains previously reported from Chifeng and other areas of Inner Mongolia (Fig. 2A). Haplotype network analysis further revealed regional variation in haplotype composition. The predominant haplotype, Hap_3 (75.6%), was widely distributed across all three regions (Fig. 2B). East Ujimqin County exhibited greater haplotype diversity than Xilinhot and Sonid Left Banner, with three distinct haplotypes (Hap_1, Hap_2, and Hap_3) identified in this region, whereas only Hap_3 was detected in Xilinhot and Sonid Left Banner (Fig. 2B).
Phylogenetic tree and haplotype analysis of G. duodenalis. (a) Phylogenetic tree of the G. duodenalis bg gene. (b) Haplotype analysis results of G. duodenalis sequences detected in this study and published G. duodenalis sequences. Different colors represent different groups, including DW, BY, SZ, Chifeng, and other regions, and the size of the circles represents the number of haplotypes.
Overall, G. duodenalis infection in livestock from the Xilingol League exhibited distinct regional and host-associated variation, with assemblages B and E as the predominant genotypes and Hap_3 as the most prevalent haplotype across the study area.
Prevalence, phylogenetic analysis, and haplotype characterization of Cryptosporidium spp.
Nested PCR detected Cryptosporidium spp. infections in livestock across multiple sites within the Xilingol League, with C. andersoni and C. xiaoi identified as the detected species. The overall prevalence of Cryptosporidium spp. was 7.8% (16/205) in cattle and sheep (Table 1). Across regions, East Ujimqin County had the lowest infection rate (2.9%), which was significantly lower than that in Sonid Left Banner (22.7%, OR = 10.00, 95% CI: 2.55–34.26, P < 0.05) and Xilinhot (16.3%, OR = 6.61, 95% CI: 1.92–20.80, P < 0.05). Within the sheep population, the infection rate in East Ujimqin County (5.5%) was significantly lower than that in Xilinhot (16.3%, OR = 5.70, 95% CI: 1.43–20. 87, P < 0.05) and Sonid Left Banner (40.0%, OR = 19.56, 95% CI: 2.48–119.60, P < 0.05). No significant regional differences were observed in the infection rates among cattle.
Phylogenetic analysis based on the small subunit ribosomal RNA (SSU rRNA) gene locus revealed that C. andersoni and C. xiaoi clustered into two distinct clades (Fig. 3A). Haplotype network analysis identified eight haplotypes among the sequences obtained in this study. The predominant haplotypes were Hap_3 (38.4%), Hap_7 (23.1%), and Hap_8 (30.8%). Hap_3 and Hap_8 were identified in both Xilinhot and Sonid Left Banner, whereas only Hap_7 was detected in East Ujimqin County (Fig. 3B).
Phylogenetic tree and haplotype analysis of Cryptosporidium spp. (a) Phylogenetic tree of the Cryptosporidium spp. SSU rRNA gene. (b) Haplotype analysis results of C. xiaoi sequences detected in this study and published C. xiaoi sequences. Different colors represent different groups, including DW, BY, SZ, China, and non-China, and the size of the circles represents the number of haplotypes.
Overall, C. xiaoi was the most frequently detected Cryptosporidium species in livestock from the Xilingol League, with Hap_3 and Hap_8 identified as the predominant haplotypes. Marked regional variation was observed in infection rates and haplotype distribution among host populations.
Prevalence, phylogenetic analysis, and haplotype characterization of E. bieneusi
Sequence-based molecular analysis revealed E. bieneusi infections in livestock across multiple sites within the Xilingol League. The overall prevalence of E. bieneusi in cattle and sheep was 4.9% (10/205) (Table 1). Positive samples were detected in East Ujimqin County, where the infection rate reached 7.2% (10/140); no infections were identified in Sonid Left Banner or Xilinhot. The infection rates among different host species were as follows: cattle, 7.9% (3/38); goats, 18.2% (2/11); and sheep, 5.5% (5/91). Although the infection rate varied among host species, the differences were not statistically significant.
Phylogenetic analysis based on internal transcribed spacer (ITS) gene sequences revealed that E. bieneusi genotypes J and BEB6 clustered within group 2, a lineage predominantly found in ruminants (Fig. 4A). Haplotype network analysis incorporating sequences from the GenBank database identified 22 haplotypes in total. Among these, only 2 were detected in the present study: Hap_9, which was primarily found in goats and sheep from East Ujimqin County, and Hap_2, which was detected exclusively in cattle (Fig. 4B). Hap_9 was the predominant haplotype in East Ujimqin County, accounting for 80% of the detected sequences, and exhibited distinct regional and host-associated distribution patterns.
Phylogenetic tree and haplotype analysis of E. bieneusi. (a) Phylogenetic tree of E. bieneusi ITS gene. (b) Haplotype analysis results of E. bieneusi sequences detected in this study and published E. bieneusi sequences. Different colors represent different groups, including Chifeng cattle, DW sheep, DW goat, and DW cattle, and the sizes of the circles represent the number of haplotypes.
Overall, E. bieneusi infections in livestock from the Xilingol League were confined to East Ujimqin County, where genotype diversity was limited to two group 2 genotypes (J and BEB6) represented by distinct host-associated haplotypes.
Overall prevalence of gastrointestinal parasites in cattle and sheep from Xilingol League
Epidemiological analysis identified widespread intestinal parasite infections and frequent co-infections among cattle and sheep in the Xilingol League. Statistical analysis showed that the overall prevalence of intestinal parasites in cattle and sheep was 80.5% (165/205), with at least one of the following parasites detected in each positive sample: Eimeria spp., nematodes, trematodes, Cryptosporidium spp., E. bieneusi, and G. duodenalis (Table S1). Significant regional differences in infection rates were observed. The infection rate in livestock from Sonid Left Banner (50.0%, 11/22) was significantly lower than that in East Ujimqin County (85.0%, 119/140; OR = 0.18, 95% CI: 0. 07–0.47, P < 0.05) and Xilinhot (79.1%, 34/43; OR = 0.26, 95% CI: 0.10–0.80, P < 0.05). Host species and regional factors both influenced parasite prevalence. Sheep (90.6%) exhibited a significantly higher infection rate than cattle (52.0%) (OR = 8.95, 95% CI: 4.09–20.12, P < 0.05). Within the same host species, goats in East Ujimqin County (90.9%) had a significantly higher infection rate than those in Sonid Left Banner (40.0%) (OR = 15.00, 95% CI: 1.27–208.10, P < 0.05). Sheep in East Ujimqin County (95.6%) also had a significantly higher infection rate compared to those in Xilinhot (79.1%) (OR = 5.76, 95% CI: 1.61–17.56, P < 0.05).
Further analysis of co-infection with two or more parasites revealed an overall mixed infection rate of 47.8% (98/205). Among the livestock population, 30.7% (63/205) had two parasites, 15.1% (31/205) had three parasites, 1.5% (3/205) had four parasites, and 0.5% (1/205) were infected with five parasites (Table S1). The most common co-infection type was protozoan-helminth co-infection, detected in 38.0% (78/205) of animals, followed by protozoan-protozoan (8.9%, 18/205) and nematode-nematode (0.9%, 2/205) combinations. Livestock infected with Eimeria spp. showed a higher prevalence of other intestinal parasites (71.3%, 87/122) than non-infected animals (47.0%, 39/83). Statistical analysis indicated that Eimeria spp. infection was associated with an increased probability of co-infection with other intestinal parasites (OR = 2.80, 95% CI: 1.58–5.08, P < 0.05).
Overall, intestinal parasite co-infections in cattle and sheep from the Xilingol League were detected across all surveyed regions, showing clear regional and host-associated variation, with protozoan-helminth combinations being the most common type identified.
DISCUSSION
Inner Mongolia, characterized by a semi-nomadic pastoral system that allows large-scale livestock movement, represents a critical ecological interface for zoonotic disease transmission in the northern grasslands of China (26). In regions such as the Xilingol League, where free-range grazing predominates, shared water sources and communal pastures facilitate fecal contamination and cross-species transmission of intestinal parasites, which remain major constraints on livestock health and productivity. Despite their economic and zoonotic importance, intestinal parasitic infections have received limited attention in regional livestock health programs, and comprehensive data on mixed infections are lacking (27, 28). To address this gap,the prevalence and genetic diversity of key intestinal parasites (G. duodenalis, Cryptosporidium spp., and E. bieneusi) in grazing livestock across Xilingol League was investigated, thereby establishing a baseline for understanding co-infection dynamics and informing integrated control strategies.
Regional and host-specific variations in parasite prevalence
Significant regional and host-associated differences were observed in the prevalence of gastrointestinal parasites among livestock in the Xilingol League. E. bieneusi prevalence was substantially lower than values reported in Hulunbuir, Inner Mongolia, suggesting regional variation in parasite distribution (29). Compared with other regions of China, the overall prevalence of intestinal helminths (nematodes and trematodes) was relatively low, whereas Eimeria spp. and G. duodenalis exhibited notably higher infection rates (30). Marked spatial differences were also evident across the study area (31–33). Livestock from East Ujimqin County and Xilinhot showed consistently higher infection rates than those from Sonid Left Banner. These regional differences likely reflect local climatic and ecological conditions that influence parasite transmission, including precipitation, vegetation, and pasture contamination. Humid environments such as those in East Ujimqin County and Xilinhot may favor parasite survival and dissemination, whereas the arid conditions of Sonid Left Banner may constrain their persistence.
Genetic variability of parasites across hosts and regions
Molecular analyses confirmed the presence of G. duodenalis assemblages B and E in cattle and sheep from Xilingol League, with assemblage B recognized as a zoonotic genotype (34). This finding suggests a potential public health risk associated with G. duodenalis infections in this region. For Cryptosporidium spp., only C. andersoni was detected in cattle, whereas sheep and goats were exclusively infected with C. xiaoi. Previous studies in Inner Mongolia have reported the presence of C. parvum, C. andersoni, C. ryanae, and C. bovis in cattle feces, and C. ubiquitum, C. andersoni, and C. xiaoi in sheep and goats (35, 36), indicating regional specificity in Cryptosporidium spp. distribution. Regarding E. bieneusi, genotypes BEB6 and J were identified as the predominant types in cattle and sheep, consistent with those previously described in ruminants (3). Positive samples were mainly found in East Ujimqin County, where Hap_9 was the most common haplotype, suggesting limited genotype diversity and localized distribution in this region.
Eimeria spp. infection facilitates mixed intestinal parasitic infections
Eimeria infection was significantly associated with increased co-infection by other intestinal parasites in herbivores from Xilingol League, consistent with findings in ruminants elsewhere (37). Mixed infections are particularly common under free-grazing systems, where environmental exposure enhances transmission risk (37, 38). Mechanistically, Eimeria infection may promote secondary colonization by disrupting intestinal integrity and modulating host immunity (37). The resulting epithelial damage and inflammation can increase susceptibility to additional infections and maintain chronic intestinal disorders. Collectively, these findings suggest that Eimeria acts as a key predisposing factor for mixed parasitic infections, underscoring the need for integrated management strategies to improve livestock health and productivity.
This study revealed clear host- and region-associated patterns of intestinal parasitic infections in livestock from Xilingol League. Eimeria spp. were the most prevalent parasites, followed by helminths and intestinal protozoa, with sheep showing higher infection levels than cattle and goats. Molecular analyses demonstrated limited genetic variation but regionally shared haplotypes of G. duodenalis and Cryptosporidium spp. Mixed infections were common, and Eimeria infection notably increased the likelihood of co-infection with other intestinal parasites. Although species-level identification of Eimeria, nematodes, and trematodes was not performed, the findings provide a comprehensive overview of parasite distribution and co-infection dynamics. These results fill an important epidemiological gap and support the need for integrated, species-targeted parasite control strategies in grazing livestock.
MATERIALS AND METHODS
Collection of fecal samples
Fecal samples were collected between 1 August and 5 August 2021, from three pastures in Xilingol League, Inner Mongolia: Sonid Left Banner (SZ), East Ujimqin County (DW), and Xilinhot (BY). A total of 205 fresh fecal samples were obtained, including 139 from sheep, 16 from goats, and 50 from cattle. Specifically, in Sonid Left Banner, samples were collected from 5 sheep, 5 goats, and 12 cattle; in East Ujimqin County, from 91 sheep, 11 goats, and 38 cattle; and in Xilinhot, from 43 sheep. All fecal samples were immediately stored at 4°C after collection and aliquoted for further analysis.
Detection of parasite eggs using the saturated saline flotation method
The saturated saline flotation method was used for parasite egg or oocyst enrichment and identification in fecal samples, following a standardized protocol. Ten grams of fecal sample was accurately weighed and transferred into a 50 mL clean beaker. Twenty milliliters of saturated sodium chloride solution was then added and thoroughly stirred with a glass rod until homogenized. The mixture was then filtered through a 60-mesh stainless steel sieve, with the filtrate collected in a 50 mL centrifuge tube. The sample was centrifuged at 3,500 × g for 5 min at 4°C. The upper layer was discarded, and the remaining liquid was gradually replaced with saturated sodium chloride solution until a convex liquid meniscus formed at the tube opening. Immediately, an 18 mm × 18 mm coverslip was placed on the liquid surface. After 2 min, the coverslip was carefully lifted vertically and placed onto a glass slide. Parasite eggs were observed under an optical microscope. The identification of egg morphology was based on standard parasitology atlases (39).
Extraction of fecal DNA
Genomic DNA was extracted from fecal samples using the FastDNA Stool Kit (MP Biomedicals, Irvine, CA, USA). Approximately 500 mg of fecal sample was processed in accordance with the manufacturer’s protocol (35). Extracted DNA samples were stored at −20°C for further analysis.
Molecular identification of intestinal protozoa using multiplex nested PCR
Because protozoan cysts are small (2 μm–5 μm in diameter), the saturated salt flotation method showed limited sensitivity for detection (27, 28). To improve diagnostic accuracy and enable genotyping, nested PCR was used for molecular identification of G. duodenalis, Cryptosporidium spp., and E. bieneusi, targeting the bg, SSU rRNA, and the ITS loci, respectively (40–42). The primer sequences, annealing temperatures, and expected amplicon sizes are provided in Table 2. Each PCR batch incorporated both a positive control (previously confirmed positive samples) and a negative control (sterile ultrapure water). The second-round PCR products (5 μL) were subjected to 1.0% (wt/vol) agarose gel electrophoresis, stained with GelstainRed (UElandy, Suzhou, Jiangsu, China), and visualized under a UV transilluminator. Positive amplicons were sent to Sangon Biotech Co., Ltd. (Shanghai, China) for bidirectional sequencing using the ABI PRISM 3730 XL DNA Analyzer (Applied Biosystems, USA). The quality of the sequence was then assessed using Chromas software, and the sequence alignment was performed via BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The genotype of E. bieneusi was determined based on established nomenclature systems (43), while Cryptosporidium spp. and G. duodenalis genotyping followed national guidelines (36).
Haplotype network and phylogenetic analysis
A neighbor-joining phylogenetic tree was constructed to analyze the genetic structure of intestinal parasites in Xilingol League. Reference sequences were downloaded from GenBank to form a comparative data set, including Cryptosporidium spp. (C. xiaoi, C. bovis, C. ryanae, C. andersoni, C. canis, C. ubiquitum, C. suis, C. parvum, and C. felis) based on SSU rRNA gene sequences; G. duodenalis assemblages A, B, C, D, and E based on bg gene sequences; E. bieneusi based on ITS gene sequences. Multiple sequence alignment and phylogenetic analyses were performed using MEGA 11 software (version 11.0.13). Genetic distances were estimated using the Kimura two-parameter model, followed by neighbor-joining phylogenetic tree construction. Bootstrap analysis with 1,000 replicates was conducted to assess the robustness of tree topology (44). Outgroup sequences were selected as follows: C. felis (MH115431.1) for Cryptosporidium spp., G. duodenalis assemblage C (LC437439.1) for G. duodenalis, and E. bieneusi genotype PtEbIX (DQ885585.1) for E. bieneusi.
To achieve further genetic characterization of local strains, reference sequences were retrieved from GenBank, including the C. xiaoi SSU rRNA gene, G. duodenalis assemblage B bg gene, and E. bieneusi ITS gene sequences. Haplotype identification was performed using DnaSP software (version 6.12.03). Haplotype network analysis was conducted using the TCS method with a 95% connection limit (45). The resulting haplotype network was visualized in PopART (version 1.7), with sample frequency annotations for each identified haplotype (46).
Statistical analysis
Differences in intestinal parasite prevalence between groups were assessed using the χ^2^ test. ORs and 95% CIs were calculated to evaluate the association between risk factors and parasitic infections. All statistical analyses were performed using SPSS software (version 25.0). Statistical significance was set at P < 0.05.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmed M. 2023. Intestinal parasitic infections in 2023. Gastroenterology Res 16:127–140. doi:10.14740/gr 162237351081 PMC 10284646 · doi ↗ · pubmed ↗
- 2Wolday D, Gebrecherkos T, Arefaine ZG, Kiros YK, Gebreegzabher A, Tasew G, Abdulkader M, Abraha HE, Desta AA, Hailu A, Tollera G, Abdella S, Tesema M, Abate E, Endarge KL, Hundie TG, Miteku FK, Urban BC, Schallig HHDF, Harris VC, de Wit TFR. 2021. Effect of co-infection with intestinal parasites on COVID-19 severity: a prospective observational cohort study. E Clinical Medicine 39:101054. doi:10.1016/j.eclinm.2021.10105434368662 PMC 8324426 · doi ↗ · pubmed ↗
- 3Ryan U, Hijjawi N. 2015. New developments in Cryptosporidium research. Int J Parasitol 45:367–373. doi:10.1016/j.ijpara.2015.01.00925769247 · doi ↗ · pubmed ↗
- 4Li W, Xiao L. 2021. Ecological and public health significance of Enterocytozoon bieneusi. One Health 12:100209. doi:10.1016/j.onehlt.2020.10020933426263 PMC 7779778 · doi ↗ · pubmed ↗
- 5Ryan UM, Feng Y, Fayer R, Xiao L. 2021. Taxonomy and molecular epidemiology of Cryptosporidium and Giardia - a 50 year perspective (1971-2021). Int J Parasitol 51:1099–1119. doi:10.1016/j.ijpara.2021.08.00734715087 · doi ↗ · pubmed ↗
- 6Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, et al.. 2013. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. The Lancet 382:209–222. doi:10.1016/S 0140-6736(13)60844-2 · doi ↗
- 7Garcia LS, Procop GW. 2014. Diagnostic medical parasitology, p 284–308. In Manual of commercial methods in clinical microbiology
- 8Coop RL, Kyriazakis I. 1999. Nutrition-parasite interaction. Vet Parasitol 84:187–204. doi:10.1016/s 0304-4017(99)00070-910456415 · doi ↗ · pubmed ↗