Emergence of coexisting blaNDM and mcr-1 genes in Escherichia coli isolates from the guts of healthy individuals
Shuang Wang, Lu Liu, Lei Du, Shanli Gao, Xiaolin Yu, Lixiao Cheng, Yuzhen Chen, Zengqiang Kou, Wenkui Sun

TL;DR
Researchers found drug-resistant E. coli strains in healthy people's guts, which could spread resistance genes through plasmids, highlighting a public health concern.
Contribution
Identifies co-occurrence of blaNDM and mcr-1 resistance genes in E. coli from healthy individuals and their presence on transferable plasmids.
Findings
All seven isolates were extensively drug-resistant, with susceptibility only to tigecycline and streptomycin.
blaNDM and mcr-1 resistance genes were located on transferable plasmids, indicating potential for horizontal gene spread.
Genomic analysis revealed diverse strains and plasmid types carrying these resistance genes.
Abstract
The emergence of blaNDM/mcr-1-positive Escherichia coli poses a huge challenge to infection control. The purpose of this study was to investigate the plasmid and genomic characteristics of blaNDM/mcr-1-positive E. coli isolates from the gut of healthy individuals. This study performed drug susceptibility testing, whole-genome sequencing (WGS), analysis of resistance genes, characterization of plasmids, and single-nucleotide polymorphism (SNP) analysis on blaNDM/mcr-1-positive E. coli isolates collected from the gut of 628 healthy individuals. Our results showed that all seven isolates were extensively drug-resistant, with susceptibility only to tigecycline and streptomycin. Notably, WGS revealed that blaNDM and mcr-1 were located on transferable plasmids, and SNP analysis placed the isolates in five distinct clades. These findings underscore the need for surveillance to monitor…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
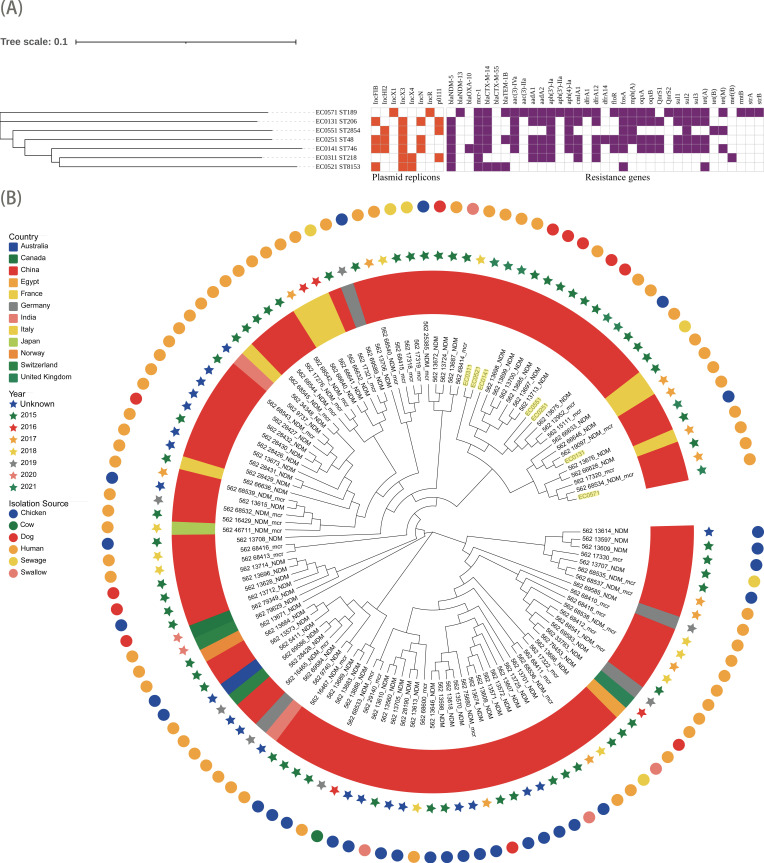 Fig 1
Fig 1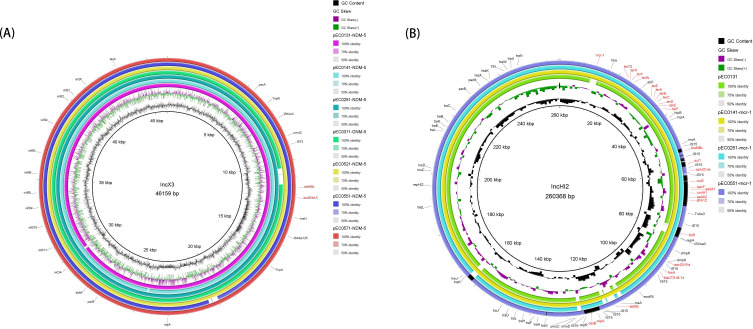 Fig 2
Fig 2- —Department of Science and Technology of Shandong Provincehttp://dx.doi.org/10.13039/100012905
- —Health and Family Planning Commission of Shandong Provincehttp://dx.doi.org/10.13039/501100010848
- —Health and Family Planning Commission of Shandong Provincehttp://dx.doi.org/10.13039/501100010848
- —Health and Family Planning Commission of Shandong Provincehttp://dx.doi.org/10.13039/501100010848
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Escherichia coli research studies · Bacterial Genetics and Biotechnology
OBSERVATION
The further spread of carbapenemase/mcr-1-positive Enterobacteriaceae could lead to a rise in untreatable infections caused by gram-negative bacteria (1). The healthy human gut commensal microbiota has been described as a reservoir of antibiotic-resistance genes (ARGs) (2). In this study, we discovered the coexistence of blaNDM and mcr-1 genes in Escherichia coli isolates from healthy individuals. We performed an in-depth genomic analysis to explore the genetic contexts and molecular characteristics of the plasmids harboring blaNDM and mcr-1. Notably, the blaNDM and mcr-1 genes located on the plasmids could be transferred from each of the isolates to recipient strains via conjugation. To our knowledge, this is the first report of blaNDM/mcr-1-positive E. coli from the guts of healthy individuals in rural communities. The patterns of antibiotic resistance and transmission in blaNDM/mcr-1-positive E. coli, particularly in healthy individuals, should be closely monitored.
In 2023, we sampled 628 non-duplicate stool samples from 628 healthy subjects from a rural community in Shandong Province and screened for carbapenemase-positive Enterobacteriaceae isolates. The demographic characteristics and geographic information of the surveyed participants are presented in Tables S1 and S2. Isolate identification was performed using matrix-assisted laser desorption ionization time-of-flight mass spectrometry. The carbapenemase-encoding gene (blaNDM) and mcr-1 gene were identified using PCR (primer sequences for blaNDM: forward, ATGGAATTGCCCAATATTATGCAC; reverse, TCAGCGCAGCTTGTCGGC; primer sequences for mcr-1: forward, GGTGGCGTTCAGCAGTCA; reverse, GCAGATGGCGTTGTTGGT). The PCR products were purified using a PCR Purification Kit (Qiagen) and Sanger sequenced to confirm genetic identity. The minimum inhibitory concentrations of the E. coli isolates for meropenem, colistin, ertapenem, ceftazidime-avibactam, tigecycline, cefotaxime, ceftazidime, ciprofloxacin, azithromycin, chloramphenicol, nalidixic acid, streptomycin, trimethoprim-sulfamethoxazole, amikacin, ampicillin, ampicillin-sulbactam, and tetracycline were determined using the broth microdilution method according to the European Committee on Antimicrobial Susceptibility Testing standards (version 12.0, 2022). E. coli ATCC 25922 was used as the quality control strain.
Whole-genome sequencing (WGS) was performed using Illumina NovaSeq 6000 and PacBio platforms at Novogene (Beijing, China). Hybrid assembly of Illumina and PacBio reads was performed using CLC Assembly Cell software (v3.2.2) and SMRT Link (v.5.0.1). Resistance genes were identified in silico using the Antibiotic Resistance Genes Database and ResFinder (3). Insertion sequence (IS) elements and integrons were identified using ISfinder (https://www-is.biotoul.fr/) and MobileElementFinder (https://cge.food.dtu.dk/services/MobileElementFinder/)http://integrall.bio.ua.pt/. BacWGSTdb provided online analyses of multilocus sequence typing (4). To obtain sequence information for the plasmids, comparisons with the National Center for Biotechnology Information (NCBI) nucleotide database (NT) were performed using the BLAST tool on the NCBI website (http://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome). Plasmid and chromosome sequences were differentiated by sequence length and alignment, with plasmid circularization confirmed by resequencing depth. Plasmid size was determined by calculating the number of bases in the plasmid sequence. Comparative analysis of different plasmids harboring the blaNDM/mcr-1 genes was performed using the Blast Ring Image Generator. The kSNP3 analysis software (with the kmer_length parameter set to 13) was used for single-nucleotide polymorphism (SNP) analysis and phylogenetic tree construction. The phylogenetic tree was then refined and visualized using Evolview (5). The transfer of two resistance genes (blaNDM and mcr-1) was studied using standard conjugation for all seven isolates.
Seven blaNDM/mcr-1-positive E. coli isolates were obtained from seven healthy subjects across 12 villages, originating from six different villages. All seven isolates exhibited resistance to ertapenem, meropenem, colistin, cefotaxime, ceftazidime, ceftazidime-avibactam, tetracycline, azithromycin, ampicillin, and ampicillin-sulbactam. However, all isolates remained susceptible to tigecycline and streptomycin. Antimicrobial susceptibility testing revealed that all isolates were extensively drug resistant, defined as non-susceptibility to at least one agent in all but two or fewer antimicrobial classes (6). The results of antimicrobial susceptibility testing are shown in Table S3. WGS results showed that the seven blaNDM/mcr-1-positive E. coli isolates harbored multiple ARGs (Fig. 1B). The most prevalent carbapenemase gene detected was blaNDM-5 (n = 6), followed by blaNDM-13 (n = 1) and blaOXA-10 (n = 1). Notably, one isolate harbored both blaNDM-5 and blaOXA-10. Three ESBL genes were detected: blaCTX-M-14 was identified in six isolates, blaCTX-M-55 in one isolate, and blaTEM-1B in two isolates. Plasmid-mediated quinolone resistance genes (oqxA, oqxB, qnrS1, and qnrS2) were detected in four isolates. Sulfonamide resistance genes sul1, sul2, and sul3 co-occurred in four isolates. Six isolates carried multiple aminoglycoside resistance genes, with aph(3′)-Ia being the most common. Furthermore, the blaNDM and mcr-1 genes were transferable from each isolate into recipient E. coli strains via conjugation, confirmed by PCR. The transfer frequency was 4.5 × 10^−8^ for blaNDM-positive plasmids and 3.6 × 10^−8^ for mcr-1-positive plasmids.
(A) ARGs and types of plasmids among blaNDM/mcr-1-positive E. coli isolates across the phylogenetic tree. (B) Phylogenetic tree of 7 blaNDM/mcr-1-positive E. coli isolates in this study and another 113 blaNDM-positive E. coli isolates in the NCBI database. Abbreviations: ARG, antibiotic resistance gene; NCBI, National Center for Biotechnology Information.
The genetic similarities between the 7 isolates in this study and another 113 blaNDM-positive E. coli isolates from various sources (human, chicken, dog, swallow, and cow) in previous studies available in public databases were characterized. SNP analysis showed that the blaNDM/mcr-1-positive E. coli isolates in this study clustered into five lineages within a major clade, with the minimum pairwise difference (between EC0131 and EC0521) being 341 SNPs. The ST206 E. coli isolate EC0131 clustered with clinical isolates of human origin from Zhejiang Province, China, showing a difference of 1,365 SNPs. The isolate EC0571 differed by 1,448 SNPs from a blaNDM/mcr-1 positive E. coli isolate from a patient in Hong Kong SAR, China. The remaining 111 isolates were more distantly related to the isolates in this study, with differences ranging between 2,176 and 37,675 SNPs (Fig. 1B).
In this study, the presence and genetic locations of blaNDM/mcr-1 in seven isolates were determined through hybrid Illumina and PacBio platform sequencing data analysis. All isolates harbored the carbapenem resistance gene blaNDM on their plasmids. The blaNDM-positive plasmids were all IncX3 type, ranging from 45 to 126 kb in size, and exhibited remarkably high sequence similarity (99.80%– 100.0% pairwise nucleotide sequence identity) (Fig. 2A). Of the mcr-1 genes in the seven isolates, three were located on IncHI2-type plasmids; one was located on an IncX4-type plasmid; and the other three were located on chromosomes. Two plasmids (pEC0141-mcr-1 and pEC0251-mcr-1) harboring mcr-1 genes displayed high sequence similarity (96% sequence coverage and 100% nucleotide identity). They shared the same genetic arrangement: dfrA12-IS15-floR-aac (3)-IVa-IS15-fosA-blaCTX-M-14-IS15-ecoRII-IS15-oqxA/B (Fig. 2B).
Circular alignments of homologous contigs from blaNDM/mcr-1-positive plasmids in this study. (A) IncX3 plasmid. (B) IncHI2 plasmid (contains mcr-1-negative plasmid pEC0131). Note: Each ring represents the corresponding plasmids shown in each figure, and plasmid types are given in the center of the rings.
The insertion sequences ISKox3, IS15, and ISAba125 were located both upstream and downstream of blaNDM-5 within the IncX3 plasmids. As a characteristic element surrounding blaNDM, ISAba125 typically formed a composite transposon named Tn125, with the blaNDM type IV secretion system genes cluster (virB1/ virB2/ virB4/ virB5/ virB6/ virB8/ virB9/ virB10/ virB11/ virD4) located upstream of the blaNDM-5 gene (7). The genetic arrangement of plasmids harboring blaNDM-5 genes was consistent: parA-ISKox3-umuD-IS15-bleMBL-blaNDM-5-insH-ISAba125-tnpA-stpA-parB-trbM-virD4virB11-virB10-virB9-virB8-virB6-virB5-virB4-virB2-virB1-taxA. These plasmids were similar to the blaNDM-5-carrying IncX3 plasmid pKP-13-8-NDM-5 (99.80%–99.99% nucleotide sequence identity and 90%–100% coverage) from a Klebsiella pneumoniae isolate from a patient blood sample in Henan Province, China, in 2020 (GenBank: MN175389.1). Additionally, genetic environment characterization revealed that blaNDM-5 in the plasmid pEC0131-NDM-5 was located in a class 1 integron named In907, with a conserved module structure of IS15-bleMBL-blaNDM-5-CH-ISAba125. Studies have reported that this conserved structure may be strongly correlated with the horizontal transfer of blaNDM-carrying plasmids (8). Three plasmids carrying mcr-1 harbored tellurium resistance-encoding genes (terZ, terA, terB, terC, terD, terE, terW, terY, terX, and terF), which reportedly provide protection against the toxic effects of tellurite, enabling bacteria to survive in tellurite-rich environments (9). Aminoglycoside resistance genes [aac(3)-IV and aph(3')-Ia], beta-lactam resistance gene (blaCTX-M-14), sulfonamide resistance gene (sul), and colistin resistance gene (mcr-1) were located in these multidrug-resistant plasmids. Three clusters of tra-encoding genes (traG/H/F/L/D/J, traL/E/K/B/C, and traW/U/N) might provide the conjugation machinery to the bacterial cell and are involved in the translocation of proteins such as virulence factors and other elements (10). We obtained seven blaNDM/mcr-1-positive E. coli isolates with different STs. The blaNDM and mcr-1 genes in several isolates were located within the same genetic environment on identical plasmid types. This phenomenon is likely driven by horizontal gene transfer of conjugative plasmids between strains. In the seven isolates tested, blaNDM and mcr-1 were located on separate IncX3 and IncHI2 plasmids, respectively. This finding suggests that plasmids carrying blaNDM and mcr-1 can mobilize and disseminate ARGs via multiple mobile genetic elements (such as transposons and insertion sequences) under strong antibiotic selective pressure.
Our study has several limitations. First, the number of strain genomes downloaded from the database was limited, so no strains closely related to the genetic relationship of the studied isolates were acquired. Second, delays in isolate analysis may have affected the accuracy of temporal trends in resistance gene dissemination. Third, the unavailability of clinical metadata, including antibiotic usage history, underlying health conditions, or travel history, constrained our ability to identify potential risk factors associated with carriage. Finally, the lack of animal or environmental sampling prevents assessment of potential zoonotic or environmental reservoirs, which is important for understanding the transmission potential of resistance genes in animal hosts and environmental reservoirs.
This study reveals the prevalence and transmission potential of blaNDM/mcr-1-positive E. coli among healthy individuals in a rural community in China. The coexistence of blaNDM and mcr-1 in Enterobacterales poses a critical public health threat, as their rapid dissemination to clinical strains could compromise the efficacy of last-resort antibiotics. Our findings emphasize the need for a One Health approach to surveillance, integrating human, animal, and environmental monitoring to curb the spread of resistance. Public health interventions, such as antibiotic stewardship programs and community education, are urgently required in endemic regions. Future studies should explore zoonotic and environmental transmission routes to inform targeted control measures.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang X-F, Peng L, Ke Y-F, Zhao D-D, Yu G-Q, Zhou Y, Li X, Weng X-B. 2023. Emergence of a clinical isolate of E. coil ST 297 co-carrying bla NDM-13 and mcr-1.1 in China. J Infect Public Health 16:1813–1820. doi:10.1016/j.jiph.2023.09.00737741016 · doi ↗ · pubmed ↗
- 2Taft DH, Liu JX, Maldonado-Gomez MX, Akre S, Huda MN, Ahmad SM, Stephensen CB, Mills DA. 2018. Bifidobacterial dominance of the gut in early life and acquisition of antimicrobial resistance. m Sphere 3:1–24. doi:10.1128/m Sphere.00441-18 · doi ↗
- 3Wang Q-J, Sun J, Li J, Ding Y-F, Li X-P, Lin J-X, Hassan B, Feng Y-J. 2017. Expanding landscapes of the diversified mcr-1-bearing plasmid reservoirs. Microbiome 5:1–9. doi:10.1186/s 40168-017-0288-028086968 PMC 5234115 · doi ↗ · pubmed ↗
- 4Luo Q, Wang Y, Fu H, Yu X, Zheng B, Chen Y, Berglund B, Xiao Y. 2020. Serotype Is associated with high rate of colistin resistance among clinical isolates of Salmonella. Front Microbiol 11:592146. doi:10.3389/fmicb.2020.59214633391208 PMC 7775366 · doi ↗ · pubmed ↗
- 5Wang S, Xie H-J, Chen Y-Z, Liu L, Fang M, Sun D-P, Xu L-C, Bi Z-W, Sun G-X, Li Y, Yu X-L, Zhang H-N, Kou Z-Q, Zheng B-W. 2022. Intestinal colonization with ESBL-producing Klebsiella pneumoniae in healthy rural villager: a genomic surveillance study in China, 2015-2017. Front Public Health 10:1–12. doi:10.3389/fpubh.2022.1017050 · doi ↗
- 6Song J, Oh S-S, Kim J, Park S, Shin J. 2020. Clinically relevant extended-spectrum β-lactamase–producing Escherichia coli isolates from food animals in South Korea. Front Microbiol 11:604. doi:10.3389/fmicb.2020.0060432390965 PMC 7188773 · doi ↗ · pubmed ↗
- 7Ma J, Xu R, Li W, Liu M, Ding X. 2024. Whole-genome sequencing of clinical isolates of Citrobacter europaeus in China carrying bla OXA−48 and bla NDM−1. Ann Clin Microbiol Antimicrob 23:1–9. doi:10.1186/s 12941-024-00699-y 38172897 PMC 10765802 · doi ↗ · pubmed ↗
- 8Yao X, Doi Y, Zeng L, Lv L-C, Liu J-H. 2016. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect Dis 16:288–289. doi:10.1016/S 1473-3099(16)00057-826842777 · doi ↗ · pubmed ↗