The microbiota of wooden cheese-ripening boards is a rich source of antimicrobial-producing bacteria against Listeria monocytogenes
Yuxing Chen, Ibrahim Zuniga Chaves, Jennifer C. McClure, Garret Suen, TuAnh N. Huynh

TL;DR
Wooden cheese boards host bacteria that produce antimicrobials, which can inhibit Listeria monocytogenes, a dangerous foodborne pathogen.
Contribution
Identification of Bacillus safensis and other antimicrobial-producing bacteria from wooden cheese boards as potential biocontrol agents.
Findings
L. monocytogenes declines on clean wooden board surfaces but can grow on cheese-covered areas.
Seven bacterial species, including Bacillus safensis, inhibit L. monocytogenes through antimicrobial factors.
B. safensis supernatant downregulates prophage elements and upregulates stress responses in L. monocytogenes.
Abstract
Wooden boards are essential tools in cheese ripening, and there is accumulating evidence that they have antimicrobial effects against foodborne bacterial pathogens, such as Listeria monocytogenes. However, poor bacterial recovery of bacteria from wood can confound the quantification of pathogen burden. To assess L. monocytogenes survival on wooden cheese boards, we applied a disruptive grinding method and tracked native board-associated bacterial counts as controls. Our data revealed that L. monocytogenes declines on clean zones of wooden boards but can replicate on areas where there is suitable cheese. Our microbiota analysis revealed diverse bacterial communities on wooden board surfaces, with a prominent presence of bacteria in the genera Brevibacterium, Brachybacterium, and Staphylococcus. We further identified seven bacterial species that inhibit L. monocytogenes, belonging to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
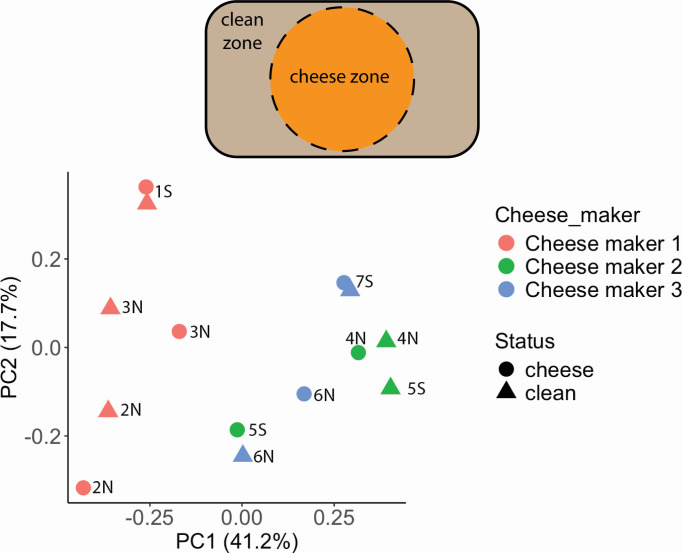 Fig 1
Fig 1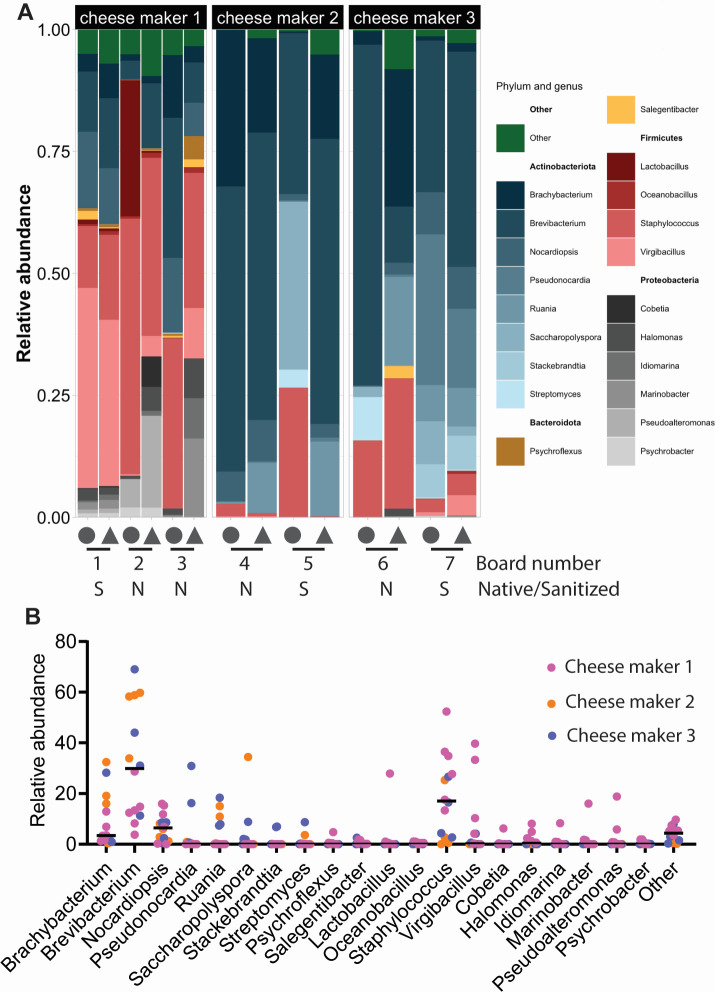 Fig 2
Fig 2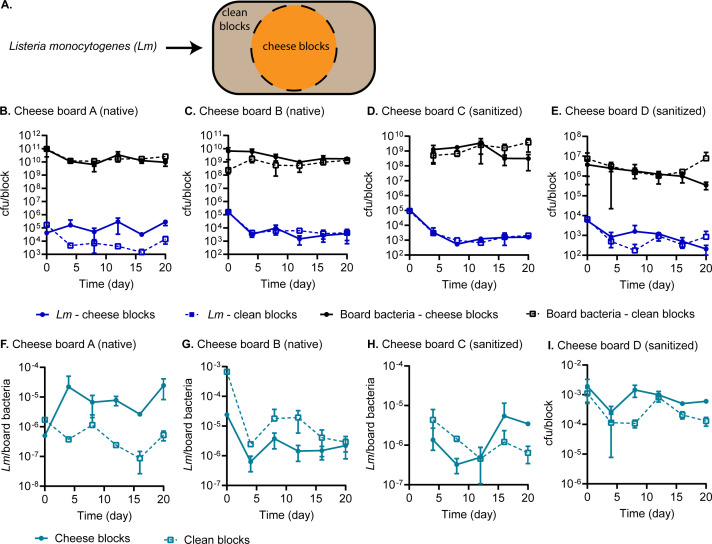 Fig 3
Fig 3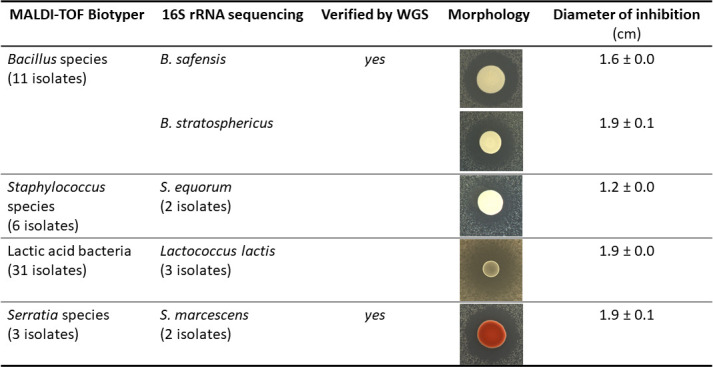 Fig 4
Fig 4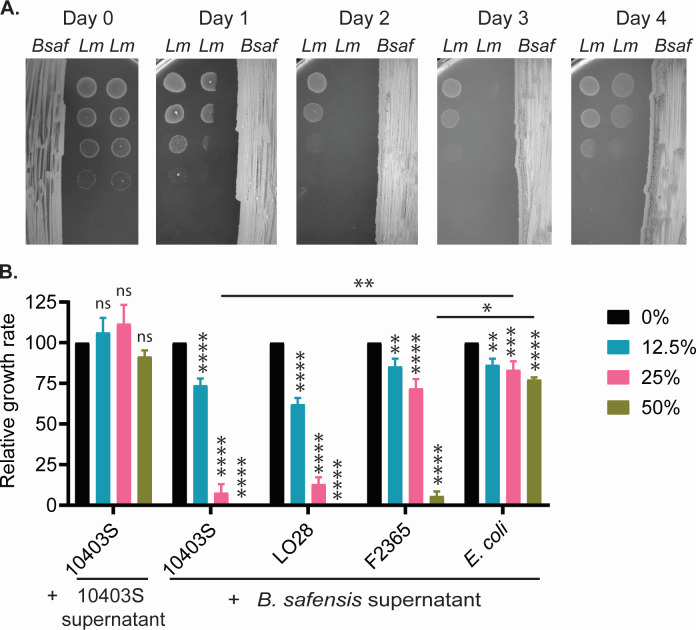 Fig 5
Fig 5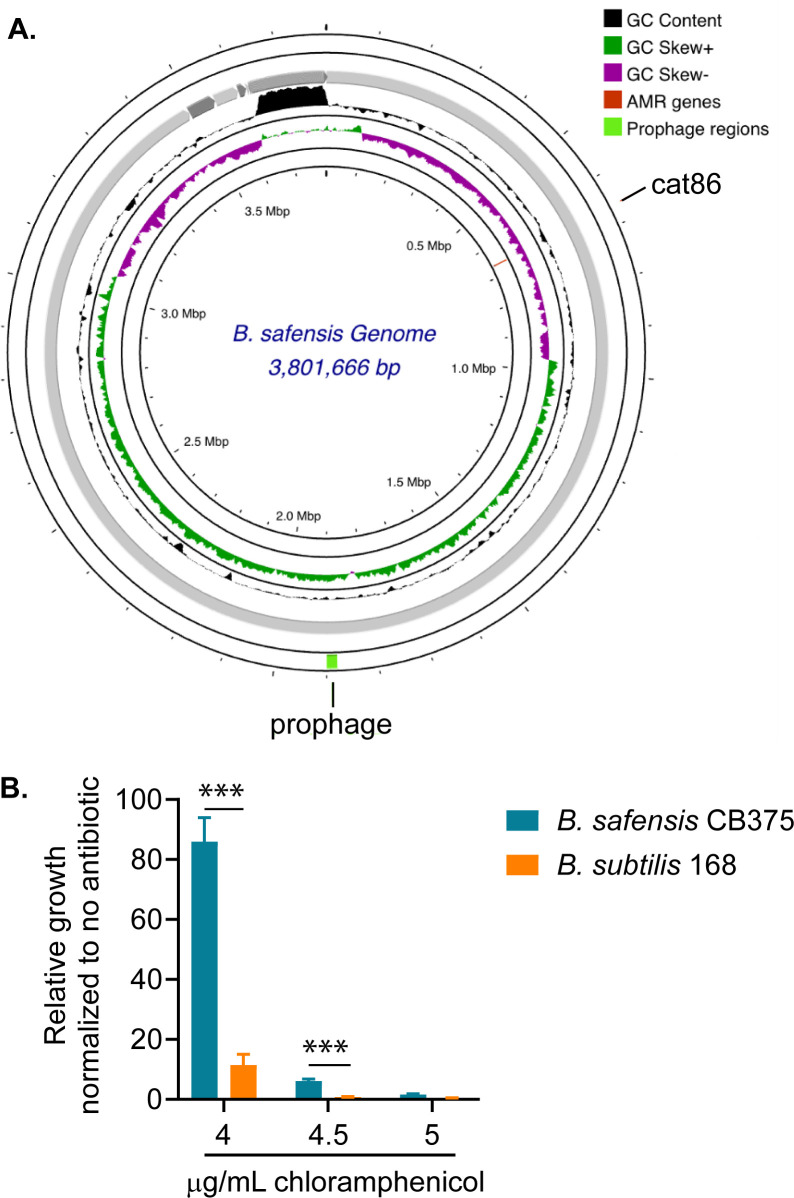 Fig 6
Fig 6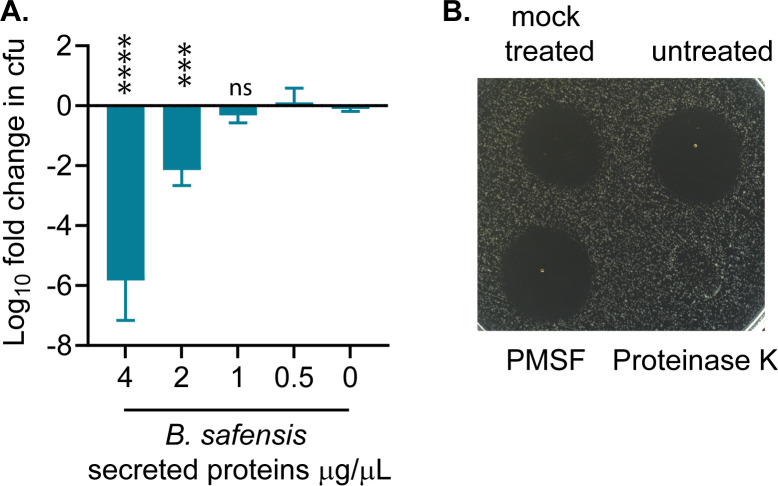 Fig 7
Fig 7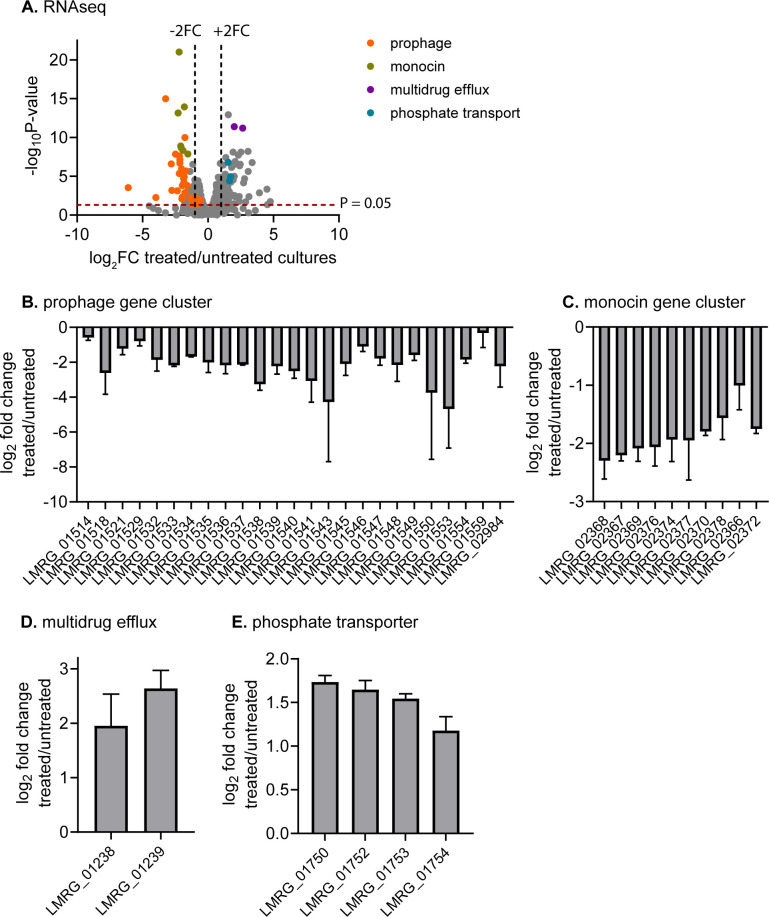 Fig 8
Fig 8| BGC no. | Annotation | Similar known BGC | Reference organism |
|---|---|---|---|
| 1 | Betalactone | Fengycin |
|
| 2 | LAP;RRE-containing | Plantazolicin |
|
| 3 | NI-siderophore;terpene | Schizokinen | |
| 4 | RRE-containing | No matches found | |
| 5 | NRP-metallophore;NRPS | Bacillibactin |
|
| 6 | Other | Bacilysin |
|
| 7 | Betalactone | No matches found | |
| 8 | RiPP-like | No matches found | |
| 9 | T3PKS | No matches found | |
| 10 | Terpene | No matches found | |
| 11 | NRPS | Lichenysin |
|
| 12 | Sactipeptide | Sporulation killing factor |
|
| 13 | NRPS | Surfactin |
|
| 14 | NRPS | Lichenysin |
|
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
- —Dairy Managementhttp://dx.doi.org/10.13039/100007513
- —U.S. Department of Agriculturehttp://dx.doi.org/10.13039/100000199
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsListeria monocytogenes in Food Safety · Indoor Air Quality and Microbial Exposure · Bacterial biofilms and quorum sensing
INTRODUCTION
Wooden boards have long been used for cheese ripening and remain popular among artisanal cheese makers, accounting for the ripening of ~500,000 tons of cheese in Europe every year (1). The porous and hygroscopic properties of wood are considered essential in the development of cheese sensory quality during ripening. Indeed, cheese made with wooden tools has been reported to exhibit a richer volatile compound profile than cheese made with other tools, due to the impact of wood on cheese properties (2). Wood used in cheese ripening is typically dried to 15%–18% humidity without chemical treatments. Wood with high humidity might support mold or Pseudomonas fluorescens. By contrast, dry wood might promote the development of Serratia species, which cause thick, strong rinds and red defects (1). Therefore, cheese makers often intentionally select woods with suitable hydrometry to achieve desirable levels of cheese humidity and ripening kinetics.
Wooden boards harbor a surface microbiota that is rich in composition and diverse among boards (1, 3–6). For instance, a comprehensive survey of wooden boards in 18 dairy facilities in Sicily, Italy, revealed at least 43 bacterial genera on each board (5). Although those boards exhibited distinct microbiota compositions, some bacterial groups are commonly present in significant abundances, including Staphylococcus, Brevibacterium, and Corynebacterium. Interestingly, these three genera were also found to be dominant in a survey of wooden cheese boards in Wisconsin, United States (4). Given that these bacteria are among the core members of the cheese rind microbial community (7), it remains unclear what microorganisms are native to wood that transfer to the cheese surface.
Bacteria on wooden board surfaces are considered a major source of cheese rind microbiota whose metabolic activities drive cheese flavor development (1, 8). The biofilm formed by these bacteria is stable and can withstand some sanitation treatments. This persistence has raised food safety concerns, highlighted by a brief US Food and Drug Administration ban of wooden boards in cheese ripening in the United States. Although this ban was quickly reversed, and wooden boards have a long history of safe use, dairy processing facilities are frequently contaminated with foodborne pathogens that may transmit to food contact surfaces. Therefore, thorough assessments of pathogen behavior on cheese boards are necessary to guide ripening process design.
Listeria monocytogenes is an invasive foodborne pathogen that is frequently associated with dairy product outbreaks. L. monocytogenes isolates from dairy products are particularly well-adapted to host conditions and gut colonization. A majority of those isolates are of clonal complex 1, which is hyper-virulent and strongly associated with human clinical samples (9). L. monocytogenes is ubiquitously present in dairy farms and is prevalently shed by cattle and therefore easily contaminates raw milk and dairy processing plants (10–13). Although pasteurization is effective at killing L. monocytogenes, this pathogen can contaminate dairy products at any processing step following pasteurization (14). The persistence of L. monocytogenes in dairy processing plants is well documented, and this pathogen can be detected at many locations within a plant, especially in difficult-to-reach areas such as drains, cracked surfaces, joints of equipment, and conveyor belts (14). From a physiological standpoint, L. monocytogenes has an array of stress response mechanisms that enable its persistence under dairy processing conditions and replication in dairy products. For instance, the general stress response regulator, σ^B^, is activated by many stress conditions and upregulates more than 200 genes to adapt to low pH, high osmolarity, and refrigeration, all of which are relevant in dairy production (15). Soft cheeses are particularly permissive to L. monocytogenes replication due to comparatively high humidity and neutral pH. Furthermore, L. monocytogenes is notorious for robust biofilm formation on different materials, such as glass, stainless steel, polystyrene, and polytetrafluoroethylene. These biofilms provide additional protection against antimicrobial treatments (16, 17).
Despite the widespread presence of L. monocytogenes in dairy processing plants, wooden boards have not been identified as the source of L. monocytogenes contamination in cheese. On the contrary, there is evidence for the antimicrobial effect of wooden boards. For instance, wood transfers significantly less L. monocytogenes to cheese than plastic and glass (18). Furthermore, L. monocytogenes burdens have been shown to significantly reduce following inoculation on native wooden boards (19). Interestingly, heat-inactivated wooden boards allow for L. monocytogenes replication, suggesting that the board surface microbiota confers inhibition (19). However, since L. monocytogenes recovery rate from wood is poor, at no more than 30% even with destructive methods (20), alternative experimental designs are necessary to accurately quantify L. monocytogenes survival rates on wooden boards. Nevertheless, a recent study found that culturable bacteria from wooden boards inhibit L. monocytogenes in laboratory culture, although antagonistic bacteria have not been systematically identified (21).
In this study, we aimed to assess L. monocytogenes survival on the wooden cheese board surface and to systematically identify board-associated bacteria that inhibit L. monocytogenes. To address the technical challenge of poor bacterial recovery from wood, we concurrently tracked both L. monocytogenes and wooden board-associated bacteria as controls. We found L. monocytogenes burdens to decline in most instances on sanitized and native boards, but at different rates and extents among boards. In one exception, we observed an increase in L. monocytogenes abundances relative to board-associated bacteria, although cheese residues likely contributed to this increase. From a small set of wooden boards from three cheese makers, we found total microbial DNA from wood shavings to be highly abundant in genera belonging to Staphylococcus, Brevibacterium, and Brachybacterium, consistent with two previous surveys of wooden boards elsewhere and with cheese rind microbiota (4, 5). However, our taxonomic survey of live, culturable bacterial communities from those boards revealed additional genera that are not typically associated with cheese rind, such as Bacillus. Exploiting this microbial diversity, we identified six bacterial species of the Bacillus, Staphylococcus, Lactococcus, and Serratia genera that inhibit L. monocytogenes. We focused on a novel isolate of Bacillus safensis as a potential biocontrol candidate and found it to potently inhibit L. monocytogenes, likely through secreted antimicrobial peptides. In response, L. monocytogenes significantly downregulated the prophage and monocin elements. Taken together, our findings indicate that the wooden cheese ripening board microbiota is a rich source of natural antimicrobials against L. monocytogenes.
RESULTS
Total microbial DNA from wooden boards harbors a diverse microbiota with similar compositions to cheese rind
Despite research interests in the microbial ecology of wooden cheese boards, only two studies have systematically determined the wooden board microbiota by next-generation sequencing (4, 5). To expand our knowledge of the microbial diversity on wooden cheese boards, we obtained seven wooden boards from three different cheese makers in Wisconsin, United States. These cheese makers are distinct from those surveyed in a previous study (4). Wooden boards are typically sanitized after cheese ripening, and we predicted that sanitation would significantly impact board microbiota. Therefore, we also obtained sanitized boards from each cheese maker. Boards from cheese maker 1 were made of cedar, including board 1 (sanitized), board 2 (native, used for a smear-ripened cheese), and board 3 (native, used for a hard cheese). Boards from cheese maker 2 were made of pine, including board 4 (native, used for a hard cheese) and board 5 (sanitized). Boards from cheese maker 3 were also made of pine, including board 6 (native, used for a hard cheese) and board 7 (sanitized). We noticed that each board had a circular cheese mark where a cheese block had been placed during ripening, as mapped out in Fig. 1. We therefore analyzed the bacterial compositions of “clean” zones separately from “cheese” zones on each board to avoid the confounding presence of cheese microbiota in our analyses.
Wooden boards harbor distinct microbiota compositions. Top panel: clean zone and cheese zone on each wooden board. Bottom panel: principal component analysis of bacterial communities on wooden boards from three cheese makers. Each data point represents the microbiota composition of a native or sanitized wooden board from a cheese maker. Each board is marked with the board number, N indicates native boards, and S indicates sanitized boards.
We first performed a taxonomic survey using board wood shavings as previously described (4). Briefly, total DNA was extracted from the top 2 mm of each board, obtained by grinding, and subjected to short-read Illumina-based next-generation sequencing of the 16S rRNA gene. After denoising and decontamination, we obtained a total of 183,918 high-quality reads and grouped them into 62 operational taxonomy units (OTUs). These OTUs belong to 12 phyla, 16 classes, 44 orders, 72 families, and 93 genera. Due to the limited availability of boards, we did not obtain sufficient biological replicates of each board type to reliably compute α-diversity indices. However, we found these communities to have both rich and diverse microbiota, with each board harboring between 8 and 58 genera (median of 22).
Overall, the bacterial compositions from board wood shavings were reminiscent of cheese rind microbiotas and consistent with previous surveys of wooden board microbiotas (4, 5, 7). The three most abundant bacteria across all boards belonged to three genera: Staphylococcus (from 0.06% to 52%, median 17%), Brevibacterium (from 4% to 69%, median 30%), and Brachybacterium (from 0.4% to 32%, median 3.5%) (Fig. 2). As expected, bacterial compositions were distinguishable for wooden boards from different cheese makers, although those from cheese maker 1 were the most dissimilar (Fig. 1). From each cheese maker, microbiota composition was also distinguishable between native and sanitized boards, with most notable changes in the abundances of Staphylococcus and Virgibacillus.
Bacterial communities on wooden boards are diverse. Relative abundances of the twenty most abundant phyla on seven wooden boards are organized for each board (A) or each phylum (B). In panel A, the circle symbol indicates the cheese zone, and the triangle indicates the clean zone on each board. Bars in panel B are median abundances.
While sharing some common bacteria, each board in our microbiota survey exhibited a unique composition (Fig. 2). Two native boards from cheese maker 1 were dominated by Staphylococcus (13% and 52%), Brachybacterium (1% and 13%), and Brevibacterium (4% and 29%). We also noticed that cheese blocks from native board 2 had a high abundance of Lactobacillus (~28%), likely transferred from cheese. Somewhat distinct from others, board 3 was also abundant in Nocardiopsis (7%–10%). By comparison, the sanitized board from this cheese maker (board 1) was significantly reduced in Staphylococcus and substantially increased in Nocardiopsis.
Boards from cheese maker 2 (boards 4 and 5) were dominated by Brevibacterium (34%–60%) and Brachybacterium (16%–32%), with an additional abundance of Nocardiopsis (1.2%–8%). Additionally, Ruania was abundant on clean blocks from this cheese maker (11%–15%), but not on cheese blocks, suggesting that these bacteria are of wood or environmental origins.
Wooden boards from cheese maker 3 (boards 6 and 7) were overall abundant with Brachybacterium (1%–28%), Brevibacterium (11%–69%), and Staphylococcus (3%–27%). As observed in cheese maker 1, Staphylococcus was much reduced in abundance on a sanitized board than on a native board (~3% compared with ~20%, respectively). By contrast, Nocardia and Pseudonocardia were much more abundant on a sanitized board from this cheese maker (~8% and ~23%, respectively).
The compositions of live bacteria on wooden boards are distinct from total DNA footprints
Our taxonomic survey of wood grindings revealed a high prevalence of Staphylococcus, Brevibacterium, and Brachybacterium. These bacteria are easily grown under laboratory conditions and have distinct colony morphologies (22). However, when we isolated and identified bacteria from cheese boards using 16S rRNA sequencing, we rarely found Brevibacterium, although we still recovered Brachybacterium and Staphylococcus. Similarly, we frequently isolated Bacillus and Lactococcus, despite their extremely low abundances in our microbiota analysis. This led us to consider whether the total DNA extracted from wood grindings accurately reflects the live bacterial communities present on board surfaces after cheese ripening.
As a preliminary assessment of live bacteria on wooden boards, we used agar media to recover bacteria from the same wood shaving suspensions used in our initial survey. Based on a previous study indicating that most cheese rind bacteria are culturable (7), we plated these suspensions on TSA (rich, non-selective) and plate count agar supplemented with 0.1% milk and 1% salt (PCAMS; designed for cheese rind bacteria) (22) and incubated them for up to 5 days to maximize recovery. DNA sequencing of these cultured bacterial communities, of the near-full-length 16S rRNA gene using Nanopore-based sequencing, generated 171,926 high-quality reads, grouped into 36 OTUs, which belong to five phyla, seven classes, 14 orders, 25 families, and 36 genera. Compared with our analysis of wood grindings, this sequencing was comparable in quality but revealed a much-reduced diversity, which could be partially due to our incomplete method of bacterial recovery.
Our analysis of live bacterial communities revealed that they share many common members with the total DNA from wood grindings (Fig. S1). Most notably, Staphylococcus was the most dominant bacterium, consistently making up 10%–98% of the cultured communities across all boards. Brevibacterium and Brachybacterium were also universally present, although their abundance varied significantly. Brevibacterium was found at 0.06%–8%, whereas Brachybacterium accounted for 0.04%–85% of the communities. In addition to these core members, several other salt-tolerant bacteria were found in relatively high numbers within the active, culturable communities. These included Psychrobacter (up to 33.6%), Virgibacillus (up to 16%), Halomonas (up to 5%), Vibrio (up to 3%), Oceanobacillus (up to 3.4%), and Cobetia (up to 2.3%). Importantly, we also identified an enrichment of certain bacteria whose abundances were too low in the total DNA composition of wood grindings. These included Bacillus, Lactococcus, and Vibrio, as well as Mammaliicoccus (related to Staphylococcus) and Niallia (related to Bacillus). Interestingly, sanitized boards also harbored many bacteria that were dominant on native boards, especially Staphylococcus, Brevibacterium, and Brachybacterium (Fig. S1). Furthermore, the sanitized board from cheese maker 1 harbored somewhat similar communities to native boards, suggesting that these bacteria are part of the facility microbiome (Fig. S2).
L. monocytogenes declines in the clean zones of most wooden cheese boards
A previous study showed L. monocytogenes to be killed on the surface of native wooden boards (19). However, recovery of L. monocytogenes from wood is notoriously poor and could confound quantification of L. monocytogenes burdens on wood under prolonged incubation. We therefore examined both L. monocytogenes burdens and native cheese board bacteria as controls for our recovery method and incubation conditions.
We obtained four boards from cheese maker 1, including two native boards similar to boards 2 and 3, and two sanitized boards similar to board 1, in our microbiota analyses. We again separated clean blocks from those inside the cheese zone because cheese residues can impact L. monocytogenes growth and survival (Fig. 3A). Each board was cut into square blocks, surface inoculated with L. monocytogenes, and incubated at 12°C, 85% relative humidity for 20 days. To recover bacteria from those blocks, we applied a destructive method, shown to have the highest recovery rate, which involved shaving, grinding, and rigorous suspension in phosphate-buffered saline.
L. monocytogenes generally decline on wooden cheese boards following surface inoculation. (A) Schematic diagram of a wooden board with clean and cheese blocks, which were separated prior to surface inoculation with L. monocytogenes. Boards A and B are equivalent to boards 2 and 3, and boards (C and D) are equivalent to board 1 in Fig. 1 and 2. (B–E). Burdens of L. monocytogenes and board-associated bacteria on each block. (F–I) Relative burdens of L. monocytogenes, normalized to those of board-associated bacteria from each block. Native board-associated bacteria were not collected from board C at time point 0.
Over 20 days, board-associated bacterial counts remained steady (Fig. 3B through E). The native boards harbored ~10^9^–10^10^ cfu/block (~1.7 × 10^7^–1.7 × 10^8^ cfu/cm^2^), whereas the sanitized boards harbored ~2 × 10^6^–10^9^ cfu/block (~1.7 × 10^4^–1.7 × 10^7^ cfu/cm^2^). Examining raw counts of L. monocytogenes on wooden blocks, we found them to generally decline over the incubation period. The reduction in L. monocytogenes was most evident on clean blocks, with a ~1.5 log cfu decrease over the first 4 days, and a similar magnitude of reduction was also apparent on cheese blocks of boards B, C, and D. The only exception to this trend was on cheese blocks of board A, where L. monocytogenes burdens exhibited a modest increase on day 20 compared with day 0 (Fig. 3B through E).
Upon normalizing L. monocytogenes counts to those of board-associated bacteria on each block, we observed an apparent increase in the relative abundances of L. monocytogenes on cheese blocks of board A, with a nearly 2-log increase in cfu counts between days 0 and 20 (Fig. 3F through I). However, L. monocytogenes burdens otherwise declined on cheese blocks of other boards and on clean blocks of all boards. Clean blocks of board B were the most inhibitory, causing a ~2 log cfu reduction in L. monocytogenes over 20 days. The magnitude of L. monocytogenes decline on other blocks was ~0.5- and 1-log cfu.
The wooden board microbiota harbor bacteria that inhibit L. monocytogenes
A previous study identified Leuconostoc mesenteroides and Staphylococcus equorum, isolated from wooden boards, to inhibit the growth of L. monocytogenes (21). The microbial diversity of wooden boards in our study suggests that there might be more bacterial species from these boards that antagonize L. monocytogenes. Therefore, we systematically isolated cheese board-associated bacteria by recovering wood chip suspensions in different growth media, including TSA, Brain Heart Infusion (BHI) agar, De Man–Rogosa–Sharpe agar (MRS), and PCAMS (22). This effort yielded approximately 500 bacterial isolates that were screened for antimicrobial activity by spotting on a L. monocytogenes lawn on suitable agar. Around 220 isolates produced a zone of clearance, indicating antimicrobial activity, and these were further purified through multiple rounds.
Through the above culture-based method, we obtained 54 bacterial isolates representing four major types of colony morphology. Identification by a MALDI-TOF Biotyper revealed that those isolates belong to Bacillus, Staphylococcus, Lactococcus, and Serratia genera, although the species could not be unambiguously identified. Therefore, we selected nine isolates for 16S rRNA sequencing and identified them as shown in Fig. 4. These bacteria include two Bacillus species, many isolates of Staphylococcus equorum, Lactococcus lactis, and Serratia marcescens. The bactericidal effect of L. lactis is most likely due to bacteriocins, which are well-studied (23). Inhibition of L. monocytogenes by other bacteria could be due to secreted organic acids. To assess this, we measured pH values of their culture supernatants and found them to be mildly acidic or basic, ranging from pH 6.5 to 8.5 (Fig. S3A). In BHI broth adjusted to these pH values, L. monocytogenes growth was modestly reduced or unaffected (Fig. S3B and C). Therefore, L. monocytogenes killing was not due to the effects of pH alone, although it remains possible that these bacteria produce organic acids that exert membrane stress, independent of acid stress. Furthermore, several of these bacteria are known to produce diffusible antimicrobials. Staphylococcus equorum secretes micrococcin P1, which is bacteriostatic (21, 24), and S. marcescens produces a serrawettin lipopeptide that is bactericidal (25).
Identification of antimicrobial-producing bacteria from wooden cheese boards. Bacterial isolates that inhibited L. monocytogenes were initially identified by MALDI-TOF. A subset of those isolates was identified by Sanger sequencing of the 16S rRNA gene, and by whole genome sequencing as indicated. Inhibition of L. monocytogenes lawns was tested on 50% BHI agar and MRS agar (for Lactococcus lactis). Pictures of inhibition zones are not scaled.
Bacillus safensis secretes antimicrobials against L. monocytogenes
B. safensis and B. stratosphericus were the most potent and consistent inhibitors of L. monocytogenes in our screen (Fig. 4). We chose to focus on B. safensis, an environmental bacterium of soil origin with antifungal activity (26).
The zone of L. monocytogenes clearance surrounding B. safensis growth suggests that B. safensis might produce and secrete antimicrobials against L. monocytogenes. To further test this possibility, we plated serial dilutions of L. monocytogenes away from B. safensis on an agar medium. L. monocytogenes was indeed inhibited by B. safensis despite spatial separation, confirming that B. safensis secretes anti-Listeria factors (Fig. 5A). In this experimental setup, antimicrobial synthesis and secretion were somewhat delayed, since L. monocytogenes was only inhibited if B. safensis was pre-grown for at least 1 day prior to L. monocytogenes plating. The inhibitory effect was diminished by 4-day-old B. safensis, suggesting that secreted antimicrobials were unstable by this time point. Together, these data also reveal that B. safensis-derived antimicrobials accumulate as the culture grows, and their synthesis is not triggered by L. monocytogenes.
*B. safensis inhibits L. monocytogenes growth via secreted antimicrobial factors. (A) Inhibition of L. monocytogenes growth on BHI agar. B. safensis was streaked on one half of the agar and pre-grown for 0– 4 days at 30°C. Tenfold dilutions of L. monocytogenes cultures were spotted on the other half of the agar, on the indicated number of days after B. safensis was streaked. (B) Inhibition of L. monocytogenes growth in BHI broth at varying concentrations of B. safensis supernatant. Relative growth rate was calculated as the growth rate in the presence of added supernatant, normalized to that in BHI broth only. Statistics: one-way ANOVA within each strain, and two-way ANOVA among strains. ns: non-significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001.
To evaluate secreted antimicrobials in liquid cultures, we obtained a cell-free culture supernatant from B. safensis that had been grown to a high density (OD_600_ > 10) and tested it against L. monocytogenes growth. As controls, we also obtained L. monocytogenes culture supernatants from the same culture condition. We found L. monocytogenes to be potently inhibited by supernatant from B. safensis, but not L. monocytogenes (Fig. 5B). Furthermore, E. coli MG1655 was not inhibited by B. safensis supernatant. This observation indicates that L. monocytogenes inhibition likely occurs through antimicrobial activity of B. safensis rather than nutrient depletion in spent media.
Our assessments of L. monocytogenes inhibition thus far have focused on strain 10403S (sequence type 85). Because L. monocytogenes is genetically diverse, we expanded our analysis to include other commonly studied strains: F2365 (sequence type 1), EGD (sequence type 12), EGD-e (sequence type 35), and LO28 (sequence type 210) (27, 28). Strain F2365 was the most resistant, but all strains were completely inhibited by 50% B. safensis supernatant (Fig. 5B).
Cheese board-associated B. safensis encodes many biosynthetic gene clusters with predicted antimicrobial functions
As a first step toward developing B. safensis for antimicrobial production against L. monocytogenes, we performed whole genome sequencing to gain genetic knowledge for the isolate obtained in this study. Our cheese board B. safensis isolate, hereafter called CB375, has a genome of approximately 3.8 Mbp with 41.64% GC content, harboring 3,763 genes, encoding 3,692 proteins, 64 tRNA genes, and 6 rRNA genes (Fig. 6A). Of the 3,781 CDS predicted, 1,184 are proteins of unknown function, and 2,579 proteins have functional assignments, including 1,312 proteins with Enzyme Commission (EC) numbers, 1,799 with Gene Ontology (GO) assignments, and 752 proteins that can be mapped to Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. By phylogenetic and average nucleotide identity (ANI) analyses, we confirmed that CB375 is B. safensis and most closely related to strain FUA2118 (Fig. S4; Table S1). Antimicrobial resistance gene detection by the CARD database (29) indicated that B. safensis CB375 harbors a putative chloramphenicol acetyltransferase (CAT), which exhibits 42% identity and 66% similarity to CAT from commonly used bacterial expression plasmids (Fig. S5). In LB broth, B. safensis CB375 was noticeably more chloramphenicol resistant than B. subtilis type strain 168, which does not carry a cat gene, although their minimum inhibitory concentrations were similar: approximately 5 µg/mL for B. safensis and 4.5 µg/mL for B. subtilis (Fig. 6B).
*Genomic analysis of B. safensis strain CB375. (A) The assembled B. safensis CB375 genome with a predicted chloramphenicol acetyltransferase gene (cat86). (B) Chloramphenicol resistance of B. safensis CB375 and B. subtilis 168, grown in LB at 37°C. For each bacterium, growth at each chloramphenicol concentration was normalized to growth in LB only. Statistical analysis was performed by Student’s for the indicated pairs: **, P < 0.001.
Bacillus species produce a wide range of antimicrobial peptides, encoded by biosynthetic gene clusters (BGCs). We therefore mined the B. safensis CB375 genome for BGCs using the antiSMASH database. This analysis revealed 14 BGCs, most of which are predicted to encode non-ribosomal peptide synthases (NPRS) (Table 1; Fig. S6). Four BGCs within B. safensis CB375 are highly similar to those producing known compounds. BGC5 perfectly matches the reference BGC for bacillibactin, a siderophore that has antibacterial and antifungal activities due to iron sequestration (30). BGC2 is 91% similar to plantazolicin, a peptide of the RiPP family (ribosomally synthesized and post-translationally modified peptides) with a narrow spectrum activity towards B. anthracis, B. cereus, and B. thuringiensis (31). BGC6 and BGC12 exhibit 85% similarity to bacilysin and sporulation killing factor, respectively (32–35). Five BGCs have low similarity to fengycin (an antifungal peptide), schizokinen (a siderophore), lichenysin, and sporulation killing factor (36, 37). Although BGC14 is predicted to produce lichenysin, it is divergent from the reference BGC and most likely synthesizes a different compound (Fig. S6). The remaining five BGCs do not match with reference BGCs and are unique to B. safensis and closely related species (Table 1).
B. safensis-derived antimicrobials that inhibit L. monocytogenes are peptides or proteins
Given the presence of many antimicrobial peptide-encoding BGCs within the B. safensis genome, we sought to isolate secreted peptides from B. safensis by ammonium sulfate precipitation of its culture supernatant. These concentrated preparations of secreted compounds were bactericidal toward L. monocytogenes in a dose-dependent manner, with up to ~6 log cfu of killing after 1 h of exposure (Fig. 7A). Furthermore, proteinase K treatment abolished L. monocytogenes inhibition, further verifying that antimicrobial compounds are proteins or peptides in nature (Fig. 7B). Combined with the predicted presence of multiple BGCs, B. safensis CB375 most likely inhibits L. monocytogenes through secreted antimicrobial peptides.
*B. safensis-secreted antimicrobials against L. monocytogenes are proteins or peptides. (A) Killing of L. monocytogenes by concentrated compounds from B. safensis culture supernatants. B. safensis supernatants were concentrated with ammonium sulfate and resuspended in PBS, then used to treat L. monocytogenes cultures, at approximately 109 cfu/mL, for an hour. Statistical analysis was performed by one-way ANOVA, comparing each treatment with the control. ****, P < 0.0001; **, P < 0.001; ns: non-significant. (B) Concentrated B. safensis supernatants were treated with proteinase K for 2 h at 37°C, 0.5 mM PMSF (phenylmethylsulfonyl fluoride), or mock-treated, and spotted on a L. monocytogenes lawn to inspect for antimicrobial activities.
B. safensis supernatant induces a specific stress response by L. monocytogenes
Although L. monocytogenes was potently inhibited by B. safensis culture supernatants and concentrated peptide preparations, Bacillus species were not abundant in the surface microbiota of wooden boards in our study. We therefore examined the response of L. monocytogenes to a low dose of B. safensis culture supernatant (1.5%) that modestly inhibited L. monocytogenes growth (Fig. S7). Using a stringent fold change cutoff, we found this treatment to significantly upregulate 25 genes by ≥4-fold and significantly downregulate 16 genes by ≥4-fold compared with untreated L. monocytogenes cultures (Fig. 8). Strikingly, all downregulated genes map to the Φ10403S prophage or monocin locus (38). However, comparing two isogenic L. monocytogenes strains with and without the Φ10403S prophage, we found them to be equally sensitive to B. safensis (Fig. S8). The upregulated genes included a drug efflux system (lmo2087-lmo2088), consistent with an antimicrobial stress response. Furthermore, a putative phosphate transport system, homologous to the high-affinity PstSCAB phosphate transporter (39), and an operon of unknown function, lmo2567-lmo2568, were also significantly upregulated.
Transcriptional response of L. monocytogenes to B. safensis cell-free culture supernatant. (A) Volcano plot depicts L. monocytogenes genes that were up- and down-regulated during growth in 1.5% B. safensis supernatant, compared with untreated cultures. (B–E) Notable down- and upregulated genes in treated vs. untreated L. monocytogenes cultures. Log2 fold change in gene expression was calculated by EdgeR, using normalized read counts per million.
DISCUSSION
With a goal to expand our knowledge of wooden cheese board microbiota, we conducted a small survey of seven wooden boards from three different cheese makers in Wisconsin, United States. Although we expected an overlap between the microbiota of cheese rind and wooden boards, we hypothesized that wood harbors native bacteria that are not directly transferred from cheese. To analyze the microbiota that reside on board surfaces during storage, we obtained sanitized boards and native boards at the completion of ripening. Our analysis of total microbial DNA confirms the diversity of bacterial communities on wooden boards in cheese ripening facilities. Of the 93 genera identified on those boards, Staphylococcus, Brevibacterium, and Brachybacterium were universally present and were among the most abundant bacteria in those microbial communities. On clean zones of wooden boards outside of cheese blocks, these three genera comprised up to 36%, 60%, and 32% of the total bacterial abundance, respectively. Their abundance is congruent with two previous surveys of wooden boards in other locations (4, 5). Furthermore, we found these bacteria to also be prominent in board areas where cheese blocks had been placed, consistent with their ubiquitous presence on cheese rinds (7). All boards in our study also presented an abundance of Nocardiopsis, a genus of halotolerant bacteria that is also abundant in cheese rind communities, but not in other food products (7). Together, these observations suggest that Staphylococcus, Brevibacterium, and Brachybacterium are core members of wooden board microbial communities and highlight the reciprocal microbial transfer between wood and cheese surfaces.
Our principal component analysis indicated that bacterial signatures on clean and “cheese” zones were different on most boards, presenting an opportunity to distinguish cheese-associated bacteria from those from dairy processing environments. Upon further inspections, we observed a particularly high abundance of Ruania in clean zones of wooden boards from cheese maker 3 (up to 18% of bacterial communities), although their abundance was rather low in other cheese ripening facilities. Therefore, Ruania is likely associated with the ripening environment of cheese maker 3 and might have been deposited onto wooden boards from aerosols or splashing. More thorough analyses are necessary to distinguish environmental bacteria from those of cheese origins, and their relative contributions to the ripening process.
Analyses of microbial DNA from wood shavings offer a culture-independent, comprehensive survey of board microbiota. However, such an approach likely reports the cumulative microbial history, including nonviable microorganisms, and might not accurately reflect the live microorganisms that stably reside on board surfaces. Indeed, we frequently isolated Bacillus and Lactococcus species despite their extremely low abundances in the total bacterial DNA footprints on wooden boards and rarely encountered Brevibacterium that was expected to be abundant. Therefore, we also performed a preliminary survey of bacterial communities that can be recovered from wooden boards under laboratory conditions.
The live, culturable bacteria on those boards harbored many bacteria that were abundant in the microbiota composition of wood grindings, consistent with a previous observation that cheese rind bacteria are mostly culturable (7, 22). Although our recovery method of live bacteria was not exhaustive, we identified many live bacteria that were not detected in analyses of wood shavings, likely due to their low abundances in the total microbial footprints. As an example, Bacillus species were consistently present on boards, although they are not typically associated with cheese rind. A more thorough recovery approach will likely uncover more environmental bacteria that colonize wooden board surfaces, and those bacteria might explain variations in flavor profiles among cheese production facilities.
We compared the bacterial compositions on native and sanitized boards as an additional approach to identify native bacterial communities that stably reside on board surfaces. As expected, sanitized boards in our study were not sterile, but they interestingly harbored many bacteria that were also abundant on native, unsanitized boards. This observation suggests that cheese ripening facilities and wooden boards are also their natural habitats. Larger surveys are necessary to establish the origins and stability of resident board communities.
Our study also sought to assess L. monocytogenes survival on wooden cheese boards, with three technical improvements compared with a previous study (19). First, we applied grinding, a disruptive method, to maximize L. monocytogenes recovery from wood following surface inoculation (40). Second, recognizing that even the most effective method only recovers less than 30% of inoculated bacteria on wood, we normalized L. monocytogenes burdens to native board bacteria recovered from wooden boards. Raw L. monocytogenes counts on each board suggested that L. monocytogenes was inhibited. However, normalized burdens revealed one instance of an increase in relative abundance of L. monocytogenes, by 2-log in the “cheese” zone of board A. Therefore, internal standards are necessary controls for the quantification of pathogens from wood. Finally, our study tracked L. monocytogenes burdens for 20 days following surface inoculation, much longer than a previous study (19), thereby providing safety data for longer storage of wooden boards.
Although L. monocytogenes grew within the cheese zone of board A, our aggregated data suggest that board wood inhibits L. monocytogenes. First, L. monocytogenes burdens declined in the clean area of board A, outside of the cheese block. Second, a reduction in L. monocytogenes burdens, by 0.5- to 2-log cfu, also occurred on other boards, both in the cheese and clean zones. Of note, board A was used for ripening of a soft cheese, which might explain its permissiveness for L. monocytogenes growth where there was leftover cheese. Combining the stability of native bacterial populations and the overall decline in L. monocytogenes counts, our data suggest that wooden boards are resilient to pathogen contamination over long-term storage. Analyses of additional boards over extended periods will be necessary to identify the core antimicrobial communities on wooden boards.
The inhibition of L. monocytogenes on wooden boards can be attributed to antimicrobial activities of both wood and the resident microbiota. Trees naturally produce antimicrobial compounds as a defense against pathogens, and essential oils or polyphenolic compounds from wood have been demonstrated to effectively kill foodborne pathogens (20). However, since those studies mainly examined pathogen survival in highly concentrated antimicrobial extracts, the extent and kinetics of pathogen killing on cheeseboard wood remain to be determined. Within the wooden board microbiota, several prominent bacteria are known to produce antimicrobial compounds. For instance, L. monocytogenes is inhibited by various bacteriocins from lactic acid bacteria (23), phenazines and antimicrobial peptides from Brevibacterium linens (41, 42), and micrococcin P1 from Staphylococcus equorum (24). By systematically screening culturable bacteria from wooden cheese boards, we identified additional antimicrobial-producing bacteria whose abundances were low, such as Serratia marcescens and Bacillus species.
We focused on characterizing a wooden board B. safensis isolate, CB375, as a novel candidate for antimicrobial discovery against L. monocytogenes. B. safensis is a soil-dwelling bacterium that can colonize plant roots and stimulate plant growth (43–45), and some B. safensis strains have potent antifungal activity (26). Here, we found that B. safensis CB375 inhibits L. monocytogenes via secreted compounds that can be easily obtained and concentrated in the culture supernatant. Our bioinformatics analysis detected 14 biosynthetic gene clusters that encode antimicrobial peptides within the B. safensis CB375 genome. Six of those BGCs have no known homologs and likely encode novel compounds for future antimicrobial discovery efforts. Seven BGCs are highly similar, both in gene cluster organization and sequences, to those for known compounds, including fengycins, plantazolicin, schizokinen, bacillibactin, bacilysin, lichenysin, and sporulation killing factor. Fengycins are amphiphilic cyclic lipopeptides that are fungicidal against filamentous fungi through membrane permeabilization (36, 46). Plantazolicin has a narrow-spectrum activity toward B. anthracis, B. cereus, and B. thuringiensis and is therefore unlikely to inhibit L. monocytogenes (31). Schizokinen and bacillibactin are siderophores that can help Bacillus species in competitions with other bacteria (47), such as the inhibition of Pseudomonas by bacillibactin (30, 48), but their effect against L. monocytogenes has not been tested. Bacilysin from B. subtilis is important for cellular differentiation and can inhibit other bacteria, such as Campylobacter jejuni (34, 49), but its antimicrobial activity has not been broadly characterized. Lichenysin is a lipopeptide biosurfactant that can disperse bacterial biofilm, but there is yet no evidence for direct antibacterial activities (50, 51). Similarly, sporulation killing factors lyse kin cells within a Bacillus species population as a survival strategy under carbon source starvation and are unlikely to act against other species (36, 37). Finally, BGC13 within B. safensis CB375 harbors several genes similar to a surfactin-encoding reference BGC, but with substantial differences in genetic organization. Therefore, BGC13 likely encodes a different surfactin homolog. Surfactins, widely produced by many Bacillus species, are membrane-permeabilizing cyclic lipopeptides that can inhibit different bacteria (52, 53). However, more genetic analyses are needed to establish if their activities are physiologically significant in microbial communities (53). A recent study identified safencin E as a novel class II bacteriocin from a B. safensis isolate from bees’ gut (54). Although class II bacteriocins generally have broad-spectrum activity (55), the antimicrobial effect of safencin E toward L. monocytogenes remains to be tested. Nevertheless, the B. safensis CB375 genome does not harbor a homologous BGC for safencin E.
L. monocytogenes strain 10403S encodes two phage-like elements, a Φ10403S prophage and a monocin, that are predicted to attack other bacteria (38). Both elements were significantly downregulated in response to a sub-inhibitory concentration of B. safensis supernatant. As for other bacteria, prophage and monocin gene expression in L. monocytogenes is induced by the SOS response and activated upon treatment with DNA-damaging reagents (38). The expression of these loci is also inhibited by prophage-encoded AriS, which suppresses the SOS response, and derepressed by monocin-encoded MpaR (38, 56). Therefore, the repression of prophage and monocin suggests that B. safensis-derived antimicrobial peptides might activate AriS, inhibit MpaR, or suppress SOS response by other unknown mechanisms. Among upregulated genes, an increased expression of drug efflux is consistent with an antimicrobial stress response. The upregulated lmo2567-lmo2568 operon is homologous with the PstSCAB phosphate transporter (39), suggesting that a phosphate starvation response might be induced. However, the role of Lmo2567-Lmo2568 in phosphate transport and the impact of phosphate on L. monocytogenes antimicrobial response both need to be experimentally determined.
In summary, our study expands our knowledge of the microbial ecology on wooden boards used in cheese ripening. Our findings indicate an inhibitory effect of clean wooden boards against surface-inoculated L. monocytogenes. The relative contributions of wood and the resident microbiota to pathogen inhibition are unknown, but our identification of several bacteria that inhibit L. monocytogenes highlights wooden board microbiota as a source for antimicrobial discovery.
MATERIALS AND METHODS
Wooden boards
For microbiota analyses, seven wooden boards were obtained from three cheese makers in Wisconsin (United States), including native boards at the completion of cheese ripening or sanitized boards. Boards were sanitized in four steps: washing in a diluted detergent at ~75°C, rinsing with fresh water, dipping in a sanitizing reagent (peracetic acid), and air drying. Boards were numbered in this report as follows. All boards from cheese maker 1 were made of cedar: board 1 (sanitized), board 2 (native, used for a smear-ripened cheese), and board 3 (native, used for a hard cheese). Boards from cheese maker 2 were made of pine: board 4 (native, used for a hard cheese) and board 5 (sanitized). Boards from cheese maker 3 were made of pine: board 6 (native) and board 7 (used for a hard cheese).
For L. monocytogenes survival analyses, boards A–D were obtained from cheese maker 1. Board A was equivalent to board 2 (native board from the same smear-ripened cheese), board B was equivalent to board 3 (native board from the same hard cheese), and boards C and D were equivalent to board 1 (sanitized).
Extraction of genomic DNA and sequencing of bacterial amplicons
Genomic DNA (gDNA) extraction of microbial community on the wooden board surfaces was performed based on a published procedure (4). Wood dust, obtained by drilling and shaving, was thoroughly resuspended in phosphate-buffered saline (PBS) by gentle rotation, pelleted by centrifugation, and mechanically disrupted by bead beating, with the addition of phenol and 20% sodium dodecyl sulfate. Lysates were then centrifuged, and DNA was extracted and purified using phenol:chloroform:isoamyl alcohol (25:24:1, pH 8) (Thermo Fisher Scientific), precipitated, and washed with ethanol. DNA samples were quantified with a Qubit fluorometer (Invitrogen).
Total genomic DNA samples were PCR amplified for the 16S rRNA V4 region using universal primers as previously published (57). Briefly, 50 ng of DNA was used as template in each reaction, amplified with HotStart Ready Mix (KAPA Biosystems). PCR products were gel-purified, quantified with a Qubit fluorometer, pooled in equimolar ratios, and sequenced on an Illumina MiSeq using a v2 kit to generate paired-end 250 bp reads with custom sequencing primers.
Sequences were demultiplexed on the Illumina MiSeq system and quality filtered using the q2-demux plugin, followed by denoising with DADA2 (58). Subsequent processing was performed using QIIME 2 (59). Amplicon sequence variants were aligned using mafft (via q2-alignment) (60) and used to construct a phylogeny with fasttree2 (via q2-phylogeny) (61). Taxonomy was assigned to ASVs using the q2-feature classifier. Relative abundances of the top 20 taxa among all the samples were plotted using phyloseq and fantaxtic R packages (62, 63).
Recovery of bacteria from wooden boards and taxonomic identification
PBS suspensions of wood shavings obtained above were diluted and plated onto tryptic soy agar and PCAMS agar (22) and incubated at room temperature for up to 5 days. For each board, bacteria were pooled from all agar plates, and DNA was extracted using phenol:chloroform:isoamyl alcohol (25:24:1, pH 8) (Thermo Fisher Scientific), precipitated, and washed with ethanol. DNA samples were quantified with a Qubit fluorometer (Invitrogen).
Long-read Nanopore-based sequencing was used to characterize live microbes to the species level and thereby provide a more accurate census of their identities. Sequencing libraries were prepared from the gel-extracted 16S rRNA amplicon bands using the Rapid Barcoding Kit 96 V14 (Oxford Nanopore Technologies) and sequenced using PromethION flow cells (R10.4.1) on a P2 Solo device attached to a GridION (ONT) with MinKnow v 22.08.9. Super accurate base calling mode was selected along with barcode trimming. All other parameters were set to the default. Samples were base-called on the machine with the most up-to-date Dorado base caller.
For the full-length 16S rRNA gene amplicons, adapters were removed using Porechop (64), and sequences were filtered by size using Nanofilt (65), selecting for reads between 1,000 and 3,000 bp. Filtered fastq files were taxonomically classified using SINA (66), with the NR 99 138.2 SILVA database (67) as reference.
Survival of L. monocytogenes on the wooden board surface
Wooden boards were cut into 24 blocks of 2 × 2 inches or 3 × 3 inches. Blocks from clean areas were separated from those inside the round area where cheese had been placed during ripening. L. monocytogenes 10403S (streptomycin resistant) was grown in Brain Heart Infusion broth, washed, resuspended in PBS, and spread evenly on each wooden board block. The blocks were placed in a closed container, maintained at 85% relative humidity using a saturated K_2_SO_4_ solution. All blocks were incubated at 12°C, and relative humidity was monitored using a digital hygrometer. For L. monocytogenes recovery at each time point, the surface was shaved off wooden blocks and ground into small bits inside a clean beaker. Bacterial counts on day 0 were obtained 30 min after surface inoculation to allow L. monocytogenes impregnation on wood. Wood shavings and bits were thoroughly resuspended in PBS, with gentle disruption using sterile glass beads. Serial dilutions were plated on BHI with 200 µg streptomycin to count for L. monocytogenes cfu, or BHI without antibiotics for total bacteria. Native board-associated bacterial counts were calculated as total bacterial counts subtracted by L. monocytogenes counts.
Isolation and identification of wooden board bacteria that inhibit L. monocytogenes
Wooden boards were shaved and ground to obtain wood bits, which were thoroughly resuspended in PBS by gentle vortexing. The PBS slurries were plated onto the following agar media: BHI, TSB (Tryptic Soy Broth), PCAMS (22), and MRS. BHI, TSB, and PCAMS agar were incubated aerobically at 30°C. MRS agar was incubated at 37°C in a hypoxic chamber (4% CO_2_). Bacterial isolates were purified based on colony morphologies and spotted onto a lawn of 10^5^ cfu L. monocytogenes. Bacterial isolates that produced zones of clearance were purified and tested for at least four more rounds. Initial identification was achieved using a MALDI-TOF Bruker Biotyper at the Wisconsin Veterinary Diagnostic Laboratory. For further identification, bacterial DNA was PCR-amplified for the V4-V9 regions using primers 515F and 1492R as previously published (68). Bacterial species were identified using BLAST, with sequencing reads as the query.
Whole genome sequencing and genomic analyses
Genomic DNA was extracted from B. safensis using phenol-chloroform (pH 8.0), precipitated and washed with ethanol, and resuspended in water. Illumina whole genome sequencing was performed at SeqCenter (Pittsburgh, PA, USA) to obtain 2 × 150 bp reads at a depth of 1.33 million reads per sample. Quality control and adapter trimming were performed with FastQC. Contig and genome assembly were performed de novo by SPAdes (69), with quality assessment by QUAST (70). BV-BRC web resources were used to confirm taxonomic identification, find similar genomes (about 100 genomes), and construct a phylogenetic tree. B. safensis CB375 and closely related genomes, revealed by the phylogenetic tree, were calculated for average nucleotide identity using KBase. The genome was annotated using Prokka (71), antimicrobial resistance genes were identified using CARD (29), and prophages were identified using Phigaro and VirSorter2 (72, 73). The genome map was generated with Proksee (74). Biosynthetic gene clusters (BGCs) were predicted using antiSMASH 6.0 (75) and zol (76).
Inhibition and killing of L. monocytogenes by B. safensis
For inhibition on BHI agar, B. safensis was streaked on half of the petri dishes and pre-incubated for 0–4 days prior to spotting of serial dilutions of a L. monocytogenes culture in the other half of the petri dish. BHI agar was further incubated for another 16 h at 37°C to assess L. monocytogenes growth.
Cell-free culture supernatants were obtained from B. safensis cultures grown in BHI broth at 30°C for 16–20 h and filtered through a 0.22 µm PES membrane. As a control, cell-free culture supernatants were also obtained from L. monocytogenes cultures grown at 37°C in BHI broth. Inhibition of L. monocytogenes in BHI broth was assessed in 96-well plates containing BHI only or BHI containing indicated concentrations of cell-free culture supernatants. Bacterial growth was measured by OD_600_ at 37°C with intermittent shaking for 14 h in a plate reader. For each strain, growth rates at each supernatant concentration were normalized to the growth rate in BHI only of the same strain.
Secreted compounds from B. safensis were concentrated by adding a saturated ammonium sulfate solution (at a final 80%) to cell-free culture supernatants. Precipitates were separated by centrifugation and resuspended in PBS. Protein concentration was quantified by Bradford assay (Bio-Rad), using bovine serum albumin as a standard. For proteinase K treatment, proteinase K was added to concentrated protein/peptide preparations at a final concentration of 1 mg/mL, incubated at 37°C for 2 h, and 0.2 mM PMSF was added to inactivate proteinase K. As a control, 0.2 mM PMSF was added to another aliquot of concentrated proteins/peptides without proteinase K. For mock treatment, protein preparations were incubated at 37°C for 2 h.
For the assessment of L. monocytogenes killing, concentrated protein preparations from B. safensis culture supernatant were diluted to 4 µg/mL and added to L. monocytogenes cell suspensions in PBS. L. monocytogenes cfu were assessed at 0 and 1 h post-treatment, by plating serial dilutions on BHI agar + 200 µg/mL streptomycin.
RNA-seq and data analysis
L. monocytogenes was grown in BHI to mid-log phase (OD_600_ ~0.5) without or with 1.5% B. safensis cell-free culture supernatant. RNA was extracted, and contaminated DNA was removed as previously described. RNA-seq was performed at SeqCenter (Pittsburgh, PA) to achieve 12 million 2 × 150 bp reads per sample. Reads were mapped to the L. monocytogenes 10403S genome (NCBI:txid393133). Differentially expressed gene analysis was performed by EdgeR (77).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lortal S, Licitra G, Valence F. 2014. Wooden tools: reservoirs of microbial biodiversity in traditional cheesemaking. Microbiol Spectr 2:CM-0008-2012. doi:10.1128/microbiolspec.CM-0008-2012 · doi ↗
- 2Mallia S, Carpino S, Corallo L, Tuminello L, Gelsomino R, Licitra G. 2005. Effects of aroma profiles of Piacentinu and Ricotta cheese using different tool materials during cheesemaking, Vol. 300, p 23. Special Publication - Royal Society of Chemistry.
- 3Licitra G, Caccamo M, Valence F, Lortal S. 2017. Traditional wooden equipment used for cheesemaking and their effect on quality. In Papademas P, Bintsis T (ed), Global cheesemaking technology: cheese quality and characteristics. John Wiley & Sons, Inc.
- 4Wadhawan K, Steinberger AJ, Rankin SA, Suen G, Czuprynski CJ. 2021. Characterizing the microbiota of wooden boards used for cheese ripening. JDS Commun 2:171–176. doi:10.3168/jdsc.2020-001436338451 PMC 9623629 · doi ↗ · pubmed ↗
- 5Settanni L, Busetta G, Puccio V, Licitra G, Franciosi E, Botta L, Di Gerlando R, Todaro M, Gaglio R. 2021. In-depth investigation of the safety of wooden shelves used for traditional cheese ripening. Appl Environ Microbiol 87:e 01524-21. doi:10.1128/AEM.01524-2134550766 PMC 8579974 · doi ↗ · pubmed ↗
- 6Mariani C, Briandet R, Chamba J-F, Notz E, Carnet-Pantiez A, Eyoug RN, Oulahal N. 2007. Biofilm ecology of wooden shelves used in ripening the French raw milk smear cheese Reblochon de Savoie. J Dairy Sci 90:1653–1661. doi:10.3168/jds.2006-19017369205 · doi ↗ · pubmed ↗
- 7Wolfe BE, Button JE, Santarelli M, Dutton RJ. 2014. Cheese rind communities provide tractable systems for in situ and in vitro studies of microbial diversity. Cell 158:422–433. doi:10.1016/j.cell.2014.05.04125036636 PMC 4222527 · doi ↗ · pubmed ↗
- 8Irlinger F, Layec S, Hélinck S, Dugat-Bony E. 2015. Cheese rind microbial communities: diversity, composition and origin. FEMS Microbiol Lett 362:1–11. doi:10.1093/femsle/fnu 015 · doi ↗