A new species of Curtobacterium from apple fruit: Curtobacterium pomorum sp. nov. resistant to heavy metals
Kazuki Tsuruga, Kouki Shimamoto, Kenji Okumura, Kohei Ogura, Wataru Hashimoto

TL;DR
A new Curtobacterium species, Curtobacterium pomorum, was discovered in apple fruit and shows resistance to heavy metals like cadmium and copper.
Contribution
The discovery of a novel Curtobacterium species with unique genomic and phenotypic traits, including heavy metal resistance.
Findings
Strain KT1 has low genome sequence identity with known Curtobacterium species.
KT1 possesses a gene island linked to heavy metal resistance.
KT1 grows in cadmium/copper-contaminated soils and host plants.
Abstract
The genus Curtobacterium has been frequently found in soil and plants. In this study, we isolated the bacterial strain designated KT1 from an apple fruit. The 16S rRNA gene sequence of the KT1 was identical to those of Curtobacterium flaccumfaciens and Curtobacterium allii type strains. The average nucleotide identities between the complete genomes of the strain KT1 and the two strains were 89.3% and 89.2%, and the digital DNA–DNA hybridization values were 64.2% and 60.5%, respectively, which were less than the threshold values for species delimitation. Phylogenetic analysis based on orthologs of genus Curtobacterium categorized the strain KT1 into a cluster that contained no representative or type strain genome. The strain KT1 grew at a pH range of 5.0–10.0, while C. flaccumfaciens JCM 1347 (=ATCC 6887) grew at pH 5.0–9.0. KT1 cells had abundant anteiso-heptadecanoic acid. The strain…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
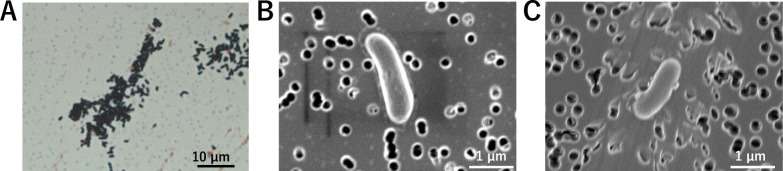 Fig 1
Fig 1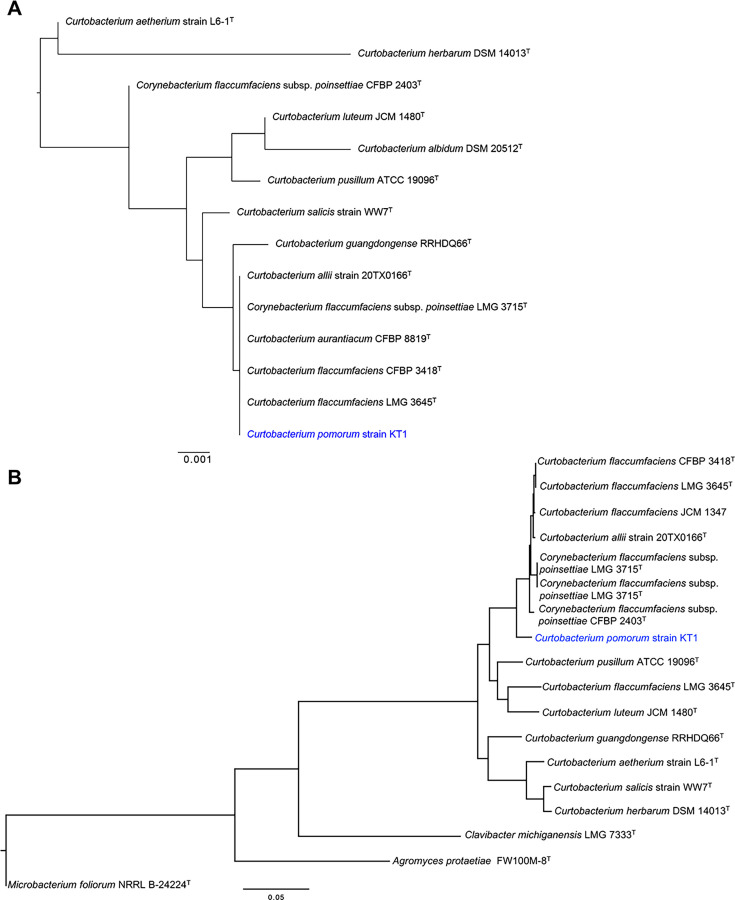 Fig 2
Fig 2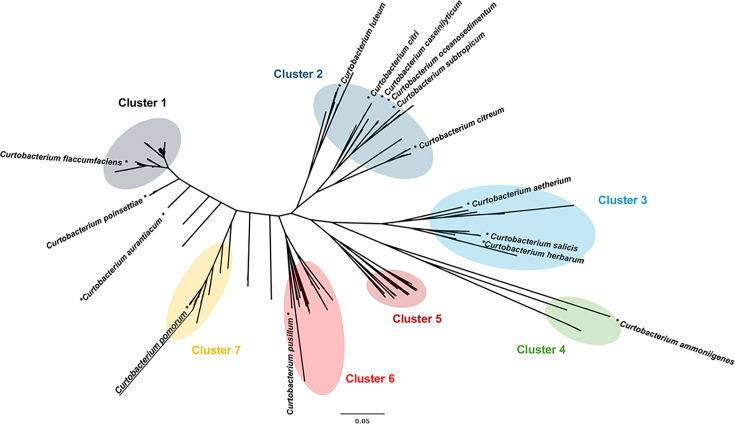 Fig 3
Fig 3 Fig 4
Fig 4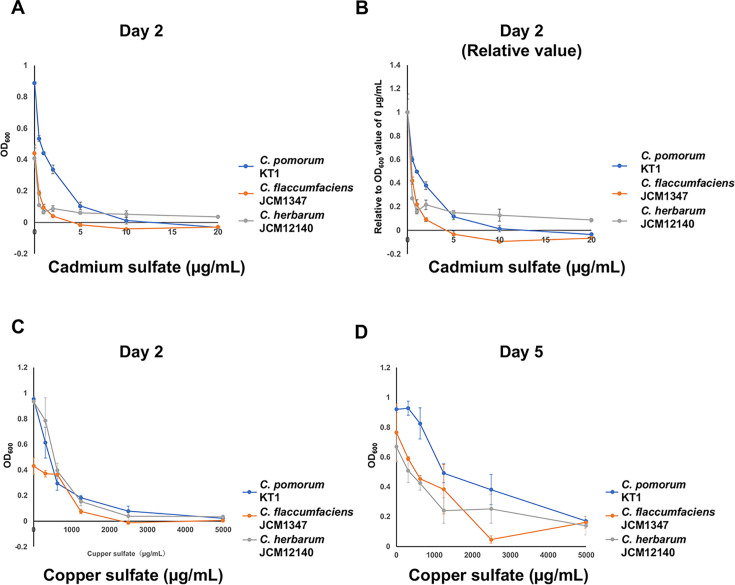 Fig 5
Fig 5| References | Accession no. | ANI (%) | DDH (%) |
|---|---|---|---|
| Genus | |||
| |
| 89.3 | 64.3 |
| |
| 89.3 | 64.2 |
| |
| 89.2 | 59.7 |
| |
| 89.2 | 60 |
| |
| 89.2 | 60.3 |
| |
| 89.2 | 60.5 |
| |
| 86.5 | 49.7 |
| |
| 85.6 | 43.1 |
| |
| 85.1 | 45.7 |
| |
| 85.1 | 45.1 |
| |
| 84.7 | 38.1 |
| |
| 84.6 | 35.8 |
| |
| 84.4 | 38.1 |
| Related strains | |||
| |
| 98.62 | 86.8 |
| |
| 98.45 | 86.6 |
| |
| 98.45 | 86.6 |
| |
| 97.06 | 72.2 |
| Characteristics | ||||||
|---|---|---|---|---|---|---|
| G+C content (mol%) | 70.8 | 68.3 | 70.8 | 70.5 | 71.5 | 71.4 |
| Motility | − | − | − | + | + | + |
| Assimilation of | ||||||
| | + | + | + | + | + | − |
| | + | + | + | w | − | − |
| | + | − | w | N.D. | N.D. | N.D. |
| Bromo-succinic acid | − | w | − | − | − | N.D. |
| Citric acid | − | w | + | − | − | N.D. |
| | + | + | + | + | + | + |
| | + | − | + | N.D. | N.D. | − |
| | w | + | + | − | + | − |
| | w | w | + | − | − | − |
| Gelatin | + | − | + | N.D. | N.D. | − |
| Glucuronamide | + | + | + | − | − | + |
| | + | + | + | − | − | + |
| Inositol | + | + | + | − | − | + |
| α- | + | + | + | − | + | + |
| | + | + | + | + | − | − |
| | + | + | + | − | + | + |
| | + | + | + | + | − | − |
| | + | + | + | − | − | − |
| | + | + | + | + | − | + |
| Tween 40 | w | + | + | − | − | N.D. |
| Fatty acid | ||||||
|---|---|---|---|---|---|---|
| C14:0 | 1.0 | 0.3 | 0.4 | N.D. | N.D. | N.D. |
| Iso-C14:0 | 0.4 | 0.5 | 0.3 | N.D. | 3.2 | 0.4 |
| Iso-C15:0 | 3.2 | 5.8 | 6.5 | 2.8 | 7.9 | 1.22 |
| Anteiso-C15:0 | 32.8 | 48.0 | 52.9 | 19.6 | 39.5 | 43.73 |
| C16:0 | 5.4 | 0.8 | 1.0 | 0.4 | 0.4 | 1.57 |
| Iso-C16:0 | 7.9 | 5.4 | 3.5 | 3.9 | 24.0 | 9.5 |
| Iso-C17:0 | 1.3 | 1.4 | 1.1 | 1.9 | 2.8 | 1.06 |
| Anteiso-C17:0 | 40.5 | 32.1 | 28.5 | 18.6 | 18.4 | 41.43 |
| C18:0 | 0.5 | 0.3 | 0.1 | N.D. | 0.4 | No data |
| Summed feature 8 | 7.0 | 3.6 | 3.8 | No data | No data | No data |
- —Toyo Institute of Food Technologyhttp://dx.doi.org/10.13039/100018134
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Metabolism and Applications · Plant Pathogenic Bacteria Studies
INTRODUCTION
Curtobacterium, a Gram-positive rod-shaped bacterium belonging to the family Microbacteriaceae, was first isolated by Yamada and Komagata in 1972 (1) and recategorized from the genus Corynebacterium. At the time of writing this paper (June 2025), 11 valid scientific names have been published for the genus Curtobacterium in the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH database (2). Curtobacterium strains are strictly aerobic and found in plant-surrounding environments, such as soil, waterweeds, grasses, and citrus leaves (3–5). As of November 2025, 13 species have been registered as validly published ones in the list of prokaryotic names with standing in nomenclature (https://lpsn.dsmz.de/genus/curtobacterium) (2), whereas 14 reference genomes were registered in the NCBI Genome database. Genomes of genus Curtobacterium are rich in G+C with contents of approximately 70%–72%. Previous reports have shown that genus Curtobacterium strains produced unsaturated menaquinones with nine isoprene units (menaquinone-9) (6, 7). Curtobacterium flaccumfaciens is the most extensively studied species within the genus Curtobacterium. C. flaccumfaciens, a phytopathogenic species causing bacterial wilt in various plants, is categorized into five pathovars, viz., C. flaccumfaciens pv. flaccumfaciens (infecting leguminous plants) (8), pv. beticola (infecting sugar beet) (9), pv. poinsettiae (infecting poinsettia) (10), pv. oortii (infecting tulips) (11), and pv. ilicis (infecting American holly) (12). In addition to C. flaccumfaciens, a recent study has reported that Curtobacterium allii 20TX0166^T^ exhibits pathogenicity to onions (13). While Curtobacterium citreum is known as a yellow pigment-producing bacterium, Curtobacterium caseinilyticum, a novel species producing yellow pigments, has been registered recently (5). In addition, Scouten et al. have reported Curtobacterium aetherium, which displays high levels of tolerance to desiccation and UV radiation (14).
Although Curtobacterium species are often isolated from soil, plant surfaces, and phyllosphere environments, their ecological functions (e.g., nutrient cycling, plant–microbe interactions, survival strategies) remain largely uncharacterized. In addition, the phylogenetic relationships among the known species are still under revision, with possible cryptic or misclassified species. In this study, we isolated a new bacterial species of the genus Curtobacterium from an apple fruit, termed Curtobacterium pomorum strain KT1, and characterized it using a polyphasic approach.
RESULTS AND DISCUSSION
Isolation of Curtobacterium strain from apple fruits
Apples were mashed, squeezed, and spread onto bacterial culture plates. After incubation at 30°C aerobically for 2 days, colonies were collected for further analysis. Based on 16S rRNA sequences, one of the obtained colonies was identified as genus Curtobacterium and named strain KT1. Other than the Curtobacterium strain, we also isolated Bacillus sp. (species undetermined by its 16S rRNA gene sequence), Luteibacter sp. (species undetermined), Pseudomonas graminis, Pseudomonas koreensis, Priestia aryabhattai, Priestia megaterium, and Stenotrophomonas nematodicola. The colony of strain KT1 was yellow, rounded, and viscous. Strain KT1 was Gram-positive and non-spore-forming (Fig. 1A). The KOH test (15) also showed Gram-positive results. Strain KT1 is an irregular, short rod-shaped bacterium without flagella (Fig. 1B and C).
Microscopic images of the novel strain C. pomorum strain KT1. (A) Gram staining result showed that the C. pomorum strain KT1 cells were stained a bluish-purple color, indicating that they are Gram-positive. The scale bar measures 10 µm. (B) Scanning electron microscopy (SEM) observation result using NanoSuit shows that C. pomorum strain KT1 is a short rod-shaped bacterium without flagella. The scale bar measures 1 µm. (C) SEM observation result using classical fixation protocol with formaldehyde and osmium tetroxide shows the same as NanoSuit. The scale bar measures 1 µm.
It is estimated that approximately 30 to 40 genes are required for complete flagellar assembly and function (16). Although the expression of flagella-related genes can vary depending on the culture conditions, the genome of this bacterium lacks the gene encoding the cap protein FliD, a key component of the flagellar structure, suggesting that this bacterium is unable to form functional flagella.
Complete genome sequence
The isolate exhibited 100% similarity to C. flaccumfaciens LMG3645^T^ and C. allii 20TX0166^T^ based on the 16S rRNA sequences (Fig. 2A). The other species with high sequence similarity of the 16S rRNA gene were Curtobacterium pusillum DSM 20527^T^ (99.52%), Curtobacterium luteum DSM 20542^T^ (99.31%), and C. citreum DSM 20528^T^ (99.31%), all of which had a coverage rate of ≥99%. Using long and short reads, the complete genome of strain KT1 was determined. The genome size of the Curtobacterium isolate was 3,901,239 bp with a G+C content of 70.8%. Annotation using the DFAST web tool (17) detected 3,730 CDSs. Assessment of genome completeness showed 99.6% of BUSCOs classified as complete. Of these, 99.4% were complete and single-copy, while 0.2% were complete but duplicated. Additionally, 0.3% were fragmented, and only 0.1% were missing (18).
Phylogenetic analysis of strain KT1 and Curtobacterium type strains. The phylogenetic trees were constructed based on (A) 16S rRNA gene and (B) whole-genome sequences, and visualized using FigTree (http://tree.bio.ed.ac.uk/software/figtree/). The phylogenetic tree based on whole-genome sequences includes three outgroup species: Agromyces protaetiae FW100M-8T, Clavibacter michiganensis LMG 7333T, and Microbacterium foliorum NRRL B-24224T. Additionally, C. flaccumfaciens JCM 1347, which was analyzed in this study, is also part of the tree.
The strain KT1 was similar to C. flaccumfaciens CFBP 3418^T^ with an average nucleotide identity (ANI) value of 89.3% and digital DNA–DNA hybridization (dDDH) value of 64.3%, both of which were less than the threshold for the identical species (ANI ≧ 95% and dDDH ≧ 70%) (19, 20) (Table 1). The strain KT1 was also similar to C. flaccumfaciens LMG3645^T^ and C. allii strain 20TX0166^T^ with ANI values of 89.3% and 89.2%, respectively, whereas the dDDH values were 64.2% and 60.5%. These results showed that the strain KT1 is a novel species belonging to the genus Curtobacterium. Phylogenetic analysis using conserved bacterial marker genes (21) showed that strain KT1 is relatively close to C. flaccumfaciens CFBP 2403^T^ and C. allii strain 20TX0166^T^, yet it is situated on a distinct phylogenetic branch (Fig. 2B). Therefore, we named it as C. pomorum strain KT1.
Characteristics of C. pomorum strain KT1
For comparison, we utilized a non-type strain C. flaccumfaciens JCM 1347 (=ATCC 6887 =DSM 20129), which shows ANI values of 97.6%, 97.6%, and 96.0% with C. flaccumfaciens LMG 3645^T^, C. flaccumfaciens pv. flaccumfaciens CFBP 3418^T^, and C. allii strain 20TX0166^T^, respectively. While C. flaccumfaciens JCM 1347 grew at pH 5.0–9.0 with pH 7.0 as the optimal condition, C. pomorum strain KT1 grew at pH 5.0–10.0 with the same optimal pH. Regarding temperature tolerance, C. flaccumfaciens JCM 1347 grew at 15°C–37°C (optimal 28°C–30°C), whereas C. pomorum strain KT1 grew at 10°C–35°C (optimal 28°C–30°C).
C. pomorum strain KT1 grew with N-acetyl-d-glucosamine, d-arabitol, d-aspartic acid, d-fructose, d-fucose, gelatin, glucuronamide, d-glucuronic acid, inositol, α-d-lactose, d-melibiose, d-raffinose, l-rhamnose, and d-sorbitol as nutrients but not bromosuccinic acid and citric acid (Table 2). Compared with that in five closely related species, the assimilation of carbon source was found to differ in several compounds, such as d-aspartic acid and gelatin.
The major (>10%) cellular fatty acids of strain KT1 were anteiso-C_17:0_ (40.5%), anteiso-C_15:0_ (32.8%), iso-C_16:0_ (7.9%), and summed feature8 (C_18:1_ ω7c and/or C_18:1_ ω6c, 7.0%). The levels of omega fatty acids were higher than those of related species. Moreover, C_16:0_ fatty acids were relatively abundant (5.4%).
Phylogenetic analysis of Curtobacterium genomes
Phylogenetic diversity in the 334 Curtobacterium genomes was examined based on single-copy orthologs (defined as orthogroups) within the genus using OrthoFinder (22). The Curtobacterium genomes were categorized into seven clusters (Fig. 3). The strain KT1 and its related species were categorized into cluster 7.
Phylogenetic trees of 334 Curtobacterium genomes. The tree was constructed with concatenated alignment of single-copy orthogroups detected by OrthoFinder (22). The detected clusters (1–7) were shown by colors. The tree was constructed using FigTree (http://tree.bio.ed.ac.uk/software/figtree/).
Cluster 1 contained C. flaccumfaciens and C. alii strains. Consistent with a previous report (23), some strains assigned as C. flaccumfaciens belonged to clusters not 1 but 2, 6, and 7. Osdaghi et al. proposed that C. flaccumfaciens pv. beticola ATCC BAA-143, which is categorized into cluster 2, should be renamed as C. citreum pv. basellae based on its genome sequence (24). Evseev et al. proposed that C. flaccumfaciens JUb65 (cluster 6) and its related species need to be assigned to distinct species (23). C. citreum was also detected in various clusters: clusters 1, 2, and 7. These results showed that 16S rRNA gene sequences are insufficient for species typing in genus Curtobacterium.
Closely related species
We found four isolates with ANI values of more than 95% to C. pomorum strain KT1: Curtobacterium sp. JUb34 (no information on isolated site) (98.6%), C. citreum strain RIT_BL8 isolated from Marcgravia umbellata (98.5%), C. citreum strain RIT_GXS8 from grape vine xylem sap (98.4%), and Curtobacterium sp. strain TC1 from a seed coat of Medicago sativa (97.1%). The genome sequences of C. citreum strains RIT_BL8 and RIT_GXS8 were distinct from the genome of C. citreum DSM 20512^T^ with ANI values of 85.6%, showing that the two strains should be typed as not C. citreum but C. pomorum.
Flagellum-related genes
C. pomorum strain KT1 showed no motility on an R2A soft agar (0.25%) plate. However, this strain harbors a gene cluster comprising 16 flagellum-related genes (Fig. S1). BLAST search showed the region (1,952,051 to 1,970,065) is conserved in C. flaccumfaciens strains, which are motile with one to three lateral or polar flagella (25). Krimi et al. reported that C. flaccumfaciens strain EHF3 showed a moderate swimming and swarming motility (26). These reports indicated that C. pomorum strain KT1 may also exhibit motility under conditions other than those provided by the R2A medium culture plate.
Genomic island
Islandviewer 4 (27) detected several genomic islands in C. pomorum strain KT1, while PHASTEST (28) showed this strain possessed no prophage region. The approximately 60 kbp genomic island spanning positions 1,283,758 to 1,344,261 was conserved exclusively in strain KT1 and absent in cluster 7 and other Curtobacterium species (Fig. 4) and showed partial sequence similarity with Brevibacterium sp., Gordonia sp., Micrococcus luteus, and related taxa. However, no significant BLAST hits were found for the full-length sequence, except in strain KT1. This island contained a gene cluster encoding heavy metal-related proteins, such as putative CueP family metal-binding proteins (Locus tags, ACGH48_06220 and ACGH48_06415 in Genbank ID CP172409.1), heavy metal translocating P-type ATPases (ACGH48_06235 and ACGH48_06350), metal-sensitive transcriptional regulator (ACGH48_06385), and metalloregulator ArsR/SmtB family transcription factor (ACGH48_06245), assuming that strain KT1 exhibits some characteristics in heavy metal transport.
Genomic island in strain KT1 genome. Locus tags were listed in Genbank (ID CP172409.1). Functions of the encoded proteins were predicted in DFAST web tool (17). The illustration was created using the drawGeneArrows3 tool, developed by Dr. Yoshiyuki Ohtsubo from Tohoku University, Japan (https://www.ige.tohoku.ac.jp/joho/).
Response to heavy metal
To assess resistance to toxic heavy metals, C. pomorum strain KT1, C. flaccumfaciens JCM 1347, and C. herbarum JCM 12140 (DSM 14013) were incubated in the presence of cadmium sulfate or copper sulfate. The growth of C. flaccumfaciens JCM 1347 was inhibited at 2 µg/mL cadmium sulfate (Fig. 5A), and no growth was observed at 5 µg/mL. C. herbarum JCM 12140 showed growth inhibition at concentrations of 1 µg/mL or higher. C. pomorum strain KT1 exhibited partial growth at both 1 and 2 µg/mL cadmium sulfate.
Resistance against toxic heavy metals. (A) Growth after 2 days of incubation in the presence of the indicated concentrations of cadmium sulfate. (B) Relative OD600 values compared to those observed without cadmium. (C and D) Growth after (C) 2 days and (D) 5 days of incubation in the presence of the indicated concentrations of copper sulfate.
To quantitatively compare the degree of growth inhibition, the optical density at 600 nm (OD_600_) of cultures grown without cadmium sulfate was normalized to 1, and relative growth was calculated accordingly (Fig. 5B). At concentrations of 0.5, 1.0, and 2.0 µg/mL cadmium sulfate, C. pomorum strain KT1 displayed significantly higher relative growth than C. flaccumfaciens and C. herbarum (Student’s t-test following ANOVA, P < 0.0167).
Next, exposure to high concentrations of copper sulfate inhibited the growth of all three Curtobacterium strains after 2 days of incubation (Fig. 5C). However, after 5 days, C. pomorum strain KT1 exhibited substantial growth at a copper sulfate concentration of 300 µg/mL (Fig. 5D). These results suggest that the genomic island present in strain KT1 may contribute to its enhanced resistance to heavy metals.
There are three types of efflux transporters involved in Cd^2+^ resistance: P-type ATPase, CBA transporters, and CDF transporter (29, 30). While strain KT1 possessed two genes encoding heavy metal translocating P-type ATPases (ACGH48_06235 and ACGH48_06350), these genes were not conserved in the other Curtobacterium species but in Micrococcus luteus, Brevibacterium sp., and Gordonia polyisoprenivorans. Next, in Salmonella enterica, CueP protein lowers copper concentrations in the cytoplasm, conferring resistance against high concentrations of copper (31, 32). Similar to P-type ATPases, C. pomorum strain KT1, but not other Curtobacterium species, possessed CueP proteins. Although the C. pomorum strain KT1 possessed two cueP genes in the island (ACGH48_06220 and ACGH48_06415), the identity of the genes was lower than the threshold of nucleotide BLAST. Identity of the two CueP proteins was moderate (44.0%). Notably, the two cueP genes showed no identity with the other bacterial genomes registered on the NCBI website by nucleotide BLAST. The amino acid sequence of one CueP protein (ACGH48_06220) showed identity with that of Microbacterium sp. (56.7% identity), the other (ACGH48_06415) resembles those found in Kocuria (78.8%) and Corynebacterium (75.9%) species. In addition, the putative metalloregulator ArsR/SmtB family transcription factor (ACGH48_06245) gene showed identity with that from Kocuria sp. (86.5%) and Brevibacterium sp. (85.3%), whereas the encoded protein showed 94.0% and 90.7% identity with Cd(II)/Pb(II)-sensing metalloregulatory transcriptional regulator CmtR from Microbacterium and Arsenicicoccus species, respectively. It is known that ArsR/SmtB family transcription factors respond to various metalloids and heavy metals, such as zinc, arsenic, and cadmium (33). Our findings indicated that the genes in strain KT1 were acquired through horizontal gene transfer from distinct bacterial lineages and subsequently mutated to acquire different functions. Further analysis is required to clarify the involvement of the genomic island in environmental adaptation.
Due to the antimicrobial properties of copper ions against bacteria and fungi by disrupting microbial cell membranes and inhibiting enzyme activity (34), copper-containing pesticides have been widely utilized in organic farming. Cadmium, a nonessential element, is toxic to host plants and microorganisms. Zhuang et al. reported accumulation of cadmium in roots, stems, and leaves of apple trees in China, while this study showed no data on apple fruits (35). Our study indicated that the acquisition of the genomic island by the strain KT1 gave an advantage for survival in environments containing these heavy metals, leading to colonization of apple fruits.
Conclusion
In this study, we isolated a new species named Curtobacterium pomorum. Genus Curtobacterium needs to be categorized based on sequences of the whole genome, but not 16S rRNA genes, because 16S rRNA gene sequencing frequently mistypes. Curtobacterium pomorum strain KT1, isolated in this study, possessed a genomic island containing genes that are suggested to export toxic metals, such as cadmium and copper. The strain KT1 was registered in Leibniz Institute DSM Z (DSM 118677) and RIKEN BRC (JCM 37513).
MATERIALS AND METHODS
Isolation of bacterium
Apple fruits cultivated in Aomori Prefecture (Japan) were purchased from a supermarket in Kyoto Prefecture (Japan). The apple must (mashed apple) was prepared by squeezing them with a pestle in sterile bottles, followed by spreading onto bacterial culture plates containing 0.5% wt/vol glucose (Nakalai Tesque, Kyoto, Japan), 1% wt/vol tryptone (Nakalai Tesque), 1% wt/vol beef extract (Becton, Dickinson and Company, NJ, USA), 0.2% wt/vol yeast extract (Nacalai Tesque), 1% wt/vol NaCl (Nakalai Tesque), and 1.5% wt/vol agar (Nakalai Tesque), with pH adjusted to 7.2 ± 0.1. Colonies were obtained after incubating the culture plates at 30°C aerobically for 2 days. The colonies were purified by streaking them onto new culture plates.
16S rRNA gene sequence analysis
16S rRNA gene was amplified using the 27F and 1492R universal primers (36). Amplicons were purified using the AMPure XP Reagent (Beckman Coulter, Brea, CA, USA) and sequenced using a 3730xl DNA Analyzer (Applied Biosystems, MA, USA).
Genome sequencing
For short-read sequencing, the genomic DNA of the Curtobacterium isolate was extracted using the DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany). Short-read sequencing (paired-end 150 bp) was performed using MGI DNBSEQ (Shenzhen, China). For long-read sequencing, genomic DNA samples were extracted from the Genomic DNA Buffer Set (QIAGEN) and Genomic tip 20/G (QIAGEN), followed by sequencing using Revio (PacBio, CA, USA). Sequencing was performed at the Bioengineering Lab. Co., Ltd. (Kanagawa, Japan). Draft genome was obtained from the short-read data by de novo assembly SPAdes (v3.15.5) (37). Complete genome sequencing was performed using Unicycler (v0.4.1) (38) using both the short- and long-read data. The genome assembly and annotation completeness were assessed by BUSCO (v5.8.3) (18). We have confirmed that the 16S rRNA gene sequences we obtained are identical to those acquired through PCR and Sanger sequencing techniques.
Calculation of ANI and DDH
ANI values were calculated using fastANI (39). A total of 335 Curtobacterium genomes were obtained from the NCBI genome database in March 2025 (Table 1; Table S1). dDDH values were calculated based on the length of all high-scoring segment pairs divided by total genome length using Genome-to-Genome Distance Calculator (GGDC 3.0) (TYGS webtool) (40) (Table 1).
Phylogenetic analysis
GTDB-Tk was employed to construct a phylogenetic tree (Fig. 2B) that includes strain KT1, the 13 Curtobacterium type strains, C. flaccumfaciens JCM 1347, and three strains belonging to the family Microbacteriaceae: Agromyces protaetiae FW100M-8^T^ (GCF_004135405.1), Clavibacter michiganensis LMG7333^T^ (GCF_021216655.1), and Microbacterium foliorum NRRL B-24224^T^ (GCF_003367705.1) (21). The tree was rooted by identifying the leaf corresponding to Microbacterium foliorum and designating it as the outgroup. Another phylogenetic tree (Fig. 3) was constructed using OrthoFinder using 33
4 Curtobacterium genomes (22). The assembly data of Curtobacterium sp. strain S6 (GCF_000710345.2) were excluded from the analysis because of its low ANI of 76.7% with strain KT1.
Morphology
Bartholomew and Mittwer-modified Gram staining (41) was performed using the Barmii-M kit (Muto Pure Chemical, Tokyo, Japan). Cells were treated with NanoSuit Solution Type I (NanoSuit Inc., Shizuoka, Japan) according to the manufacturer’s instructions, followed by SEM analysis using Hitachi SU8230 (Hitachi High-Tech, Tokyo, Japan). Additionally, SEM observation was performed on bacterial cells fixed using the conventional fixation protocol involving formaldehyde and osmium tetroxide.
Growth conditions
The strain was incubated on the above-described bacterial culture plate or a Reasoner’s 2A (R2A) (0.05% wt/vol yeast extract, 0.05% proteose peptone, 0.05% casamino acid, 0.05% glucose, 0.05% soluble starch, 0.03% sodium pyruvate, 0.03% K_2_HPO_4_, 0.05% MgSO_4_·7H_2_O) agar plate under anaerobic conditions prepared by AnaeroPack (Mitsubishi Gas Chemical, Tokyo, Japan). No growth was detected on either plate. For the salt-tolerance test, the strain was incubated for 7 days at 30°C with the addition of 0%–10.0% (wt/vol) NaCl, and growth was detected in the range of 0%–9.0%.
Carbon sources
The utilization of carbon sources was evaluated using Biolog GEN III MicroPlate according to the manufacturer’s protocol (Table 2).
Fatty acid profiles
Strain KT1 was grown in tryptic soy broth (BD Difco), freeze-dried, and subjected to gas chromatography using the Sherlock Microbial Identification System (Version 6.2) (MIDI, DE, USA) to calculate the fatty acid composition based on the TSBA6 calculation method and TSBA6 library database (Table 3). The analysis was conducted by TechnoSuruga Laboratory Co., Ltd. (Shizuoka, Japan).
Heavy metal resistance
C. pomorum strain KT1, C. flaccumfaciens JCM 1347, and C. herbarum JCM 12140 were precultured in trypticase soy broth (TSB) (BD Bioscience) media at 30°C for 2 days, and suspended in fresh TSB media containing the indicated concentrations of cadmium sulfate (0, 0.5, 1.0, 2.0, 5.0, 10, 20 µg/mL) with the initial OD_600_ = 0.01 in 96-well plates. After incubation at 30°C for 2 days, OD_600_ was measured in TECAN Infinite M Nano (TECAN, Männedorf, Switzerland). C. flaccumfaciens JCM 1347 and C. herbarum 12140 were provided by Japan Collection of Microorganisms, RIKEN BRC, which is participating in the National BioResource Project of the Ministry of Education, Culture, Sports, Science and Technology, Japan. Since some of the OD_600_ values were below the detection limit, statistical analysis using the area under the curve could not be performed. A post hoc Student’s t-test was performed following the ANOVA, with a significance threshold of P < 0.0167 to account for multiple comparisons.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yamada K, Komagata K. 1972. Taxonomic studies on coryneform bacteria. J Gen Appl Microbiol 18:417–431. doi:10.2323/jgam.18.417 · doi ↗
- 2Parte AC, Sardà Carbasse J, Meier-Kolthoff JP, Reimer LC, Göker M. 2020. List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int J Syst Evol Microbiol 70:5607–5612. doi:10.1099/ijsem.0.00433232701423 PMC 7723251 · doi ↗ · pubmed ↗
- 3Aizawa T, Ve NB, Kimoto K-I, Iwabuchi N, Sumida H, Hasegawa I, Sasaki S, Tamura T, Kudo T, Suzuki K-I, Nakajima M, Sunairi M. 2007. Curtobacterium ammoniigenes sp. nov., an ammonia-producing bacterium isolated from plants inhabiting acidic swamps in actual acid sulfate soil areas of Vietnam. Int J Syst Evol Microbiol 57:1447–1452. doi:10.1099/ijs.0.64731-017625173 · doi ↗ · pubmed ↗
- 4Behrendt U, Ulrich A, Schumann P, Naumann D, Suzuki K-I. 2002. Diversity of grass-associated Microbacteriaceae isolated from the phyllosphere and litter layer after mulching the sward; polyphasic characterization of Subtercola pratensis sp. nov., Curtobacterium herbarum sp. nov. and Plantibacter flavus gen. nov., sp. nov. Int J Syst Evol Microbiol 52:1441–1454. doi:10.1099/00207713-52-5-144112361245 · doi ↗ · pubmed ↗
- 5Feng G-D, Li J, Yang S, Zhang J, Zhu H. 2023. Curtobacterium caseinilyticum sp. nov., Curtobacterium subtropicum sp. nov. and Curtobacterium citri sp. nov., isolated from citrus phyllosphere. Int J Syst Evol Microbiol 73:006152. doi:10.1099/ijsem.0.006152 · doi ↗
- 6Collins MD, Goodfellow M, Minnikin DE. 1979. Isoprenoid quinones in the classification of coryneform and related bacteria. J Gen Microbiol 110:127–136. doi:10.1099/00221287-110-1-127107269 · doi ↗ · pubmed ↗
- 7Collins MD, Goodfellow M, Minnikin DE. 1980. Fatty acid, isoprenoid quinone and polar lipid composition in the classification of Curtobacterium and related taxa. J Gen Microbiol 118:29–37. doi:10.1099/00221287-118-1-296775048 · doi ↗ · pubmed ↗
- 8Gonçalves RM, Schipanski CA, Koguishi L, Soman JM, Sakate RK, Júnior TAFS, Maringoni AC. 2017. Alternative hosts of Curtobacterium flaccumfaciens pv. flaccumfaciens, causal agent of bean bacterial wilt. Eur J Plant Pathol 148:357–365. doi:10.1007/s 10658-016-1094-4 · doi ↗