Clostridioides difficile in equidae necropsied in Northwestern France, between 2019 and 2021
Sandrine Petry, Jackie Tapprest, Karine Maillard, Frédéric Barbut, Fabien Duquesne, Sofia Kozak, Nathalie Foucher, Maud Bernez-Romand, Ludovic Bridoux, Isabelle Poquet

TL;DR
This study found that 20% of horses in France had harmful C. difficile strains, some of which could spread to humans, highlighting a One Health concern.
Contribution
The study provides new insights into C. difficile prevalence, diversity, and virulence in French equidae, emphasizing zoonotic potential.
Findings
20% of necropsied equidae had toxigenic C. difficile strains, including ribotype 017, which caused fatal infections.
Four animals showed free toxins and signs of CDI, with ribotype 017 being the sole identified cause in two cases.
Common pathogenic ribotypes like 078 and 017 were found, suggesting possible cross-species transmission.
Abstract
Clostridioides difficile, an anaerobic, spore-forming enteropathogen, is less studied in animals than in humans despite its importance in One Health. We evaluated C. difficile occurrence, diversity, circulation, and virulence in French Equidae (n = 100) after their necropsy in northwestern France, from 2019 to 2021. We systematically recovered all cecal contents and any watery intestinal contents. We isolated C. difficile strains, determined their toxin gene profile by PCR, and established their PCR-ribotype according to the WEBRIBO database. We also performed free toxin detection. Twenty-seven Equidae were positive for C. difficile and 20 had a toxigenic strain, including one animal co-colonized by a non-toxigenic strain. Toxigenic isolates belonged to eight already described ribotypes: (i) 078 and 126 (toxin genes of wild-type size: tcdA+ tcdB+ cdt+), (ii) 005, 012, 020, AI-53, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
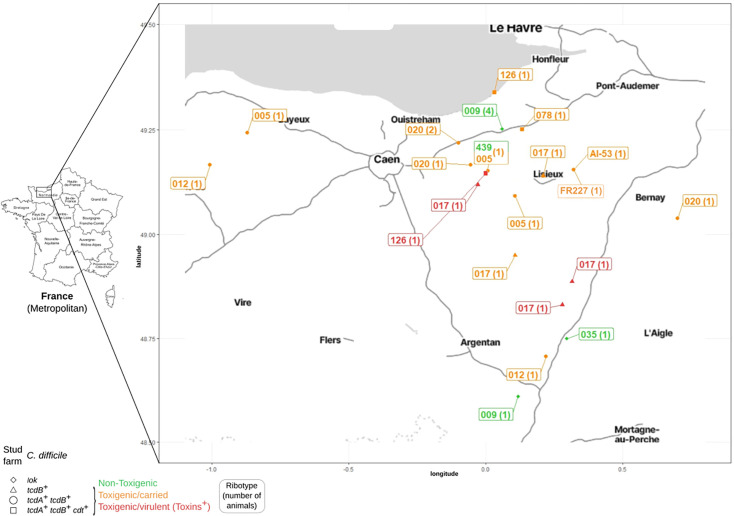 Fig 1
Fig 1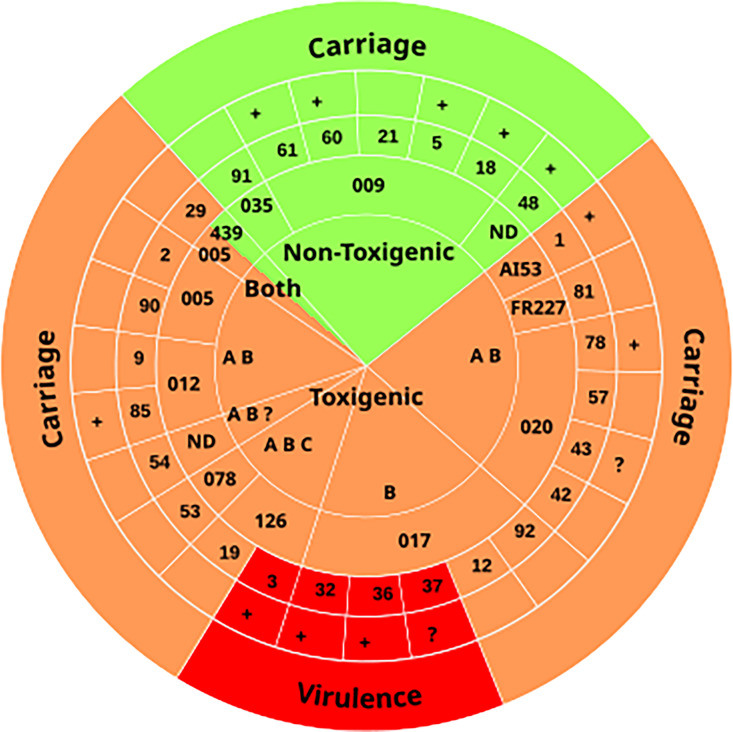 Fig 2
Fig 2|
| Presence | Pathogenicity | Toxin production | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No | Yes | Non-toxigenic | Toxigenic | Both | No | Yes | ||||
|
| ||||||||||
|
| ||||||||||
| 2019 | 32 | 15 | 0.60 | 3 | 11 | 1 | 0.91 | 8 | 4 | 0.30 |
| 2020 | 28 | 8 | 3 | 5 | 0 | 5 | 0 | |||
| 2021 | 13 | 4 | 1 | 3 | 0 | 3 | 0 | |||
|
| ||||||||||
|
| ||||||||||
| Female | 46 | 21 | 0.23 | 7 | 13 | 1 | 0.34 | 11 | 3 | 1.00 |
| Male, gelding | 26 | 6 | 0 | 6 | 0 | 5 | 1 | |||
| |
|
| ||||||||
|
| ||||||||||
| Foal | 33 | 15 | 0.55 | 6 | 9 | 0 | 0.42 | 6 | 3 | 0.79 |
| Yearling | 10 | 1 | 0 | 1 | 0 | 1 | 0 | |||
| Young | 5 | 2 | 0 | 2 | 0 | 2 | 0 | |||
| Adult | 25 | 9 | 1 | 7 | 1 | 7 | 1 | |||
|
| ||||||||||
| Thoroughbred | 30 | 12 | 0.80 | 6 | 6 | 0 | 0.15 | 6 | 0 |
|
| Trotter | 23 | 8 | 1 | 6 | 1 | 3 | 4 | |||
| Saddlebred horse | 9 | 2 | 0 | 2 | 0 | 2 | 0 | |||
| Other breeds | 9 | 5 | 0 | 5 | 0 | 5 | 0 | |||
| |
|
| ||||||||
| Thoroughbred/all other breeds | 30/41 | 12/15 | 1.0 | 6/1 | 6/13 | 0/1 |
| 6/10 | 0/4 | 0.27 |
| Trotter/all other breeds | 23/48 | 8/19 | 1.0 | 1/6 | 6/13 | 1/0 | 0.29 | 3/13 | 4/0 |
|
|
| ||||||||||
| Signs of diarrhea | ||||||||||
| One sample per animal: CAE | 59 | 11 |
| 3 | 8 | 0 | 1.0 | 8 | 0 | 0.12 |
| Two samples per animal: CAE + CDL | 14 | 16 | 4 | 11 | 1 | 8 | 4 | |||
| Signs of endo-enterotoxemia | ||||||||||
| Yes | 7 | 13 |
| 3 | 9 | 1 | 1.0 | 7 | 3 | 0.58 |
| No | 65 | 14 | 4 | 10 | 0 | 9 | 1 | |||
| |
|
| ||||||||
|
| ||||||||||
| Previous antibiotic treatment | ||||||||||
| Yes | 24 | 11 | 0.48 | 5 | 6 | 0 | 0.18 | 3 | 3 |
|
| No | 43 | 14 | 2 | 11 | 1 | 12 | 0 | |||
| |
|
|
|
|
|
|
| |||
| Previous hospitalization | ||||||||||
| Yes | 22 | 11 | 0.47 | 4 | 7 | 0 | 0.65 | 7 | 0 | 0.25 |
| No | 49 | 16 | 3 | 12 | 1 | 9 | 4 | |||
| |
|
|
|
|
|
|
| |||
|
| Equidae | Multiplex PCR | Ribotype | MLST | Vancomycin | Metronidazole | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Identification | Digestive content |
|
|
| lok |
| Profile | (WEBRIBO) | MIC (mg/L) | MIC (mg/L) | ||
| 2019-2021 | |||||||||||||
| 3 | CAE | + | + | + | − | −39 | + |
| 126 | ST11 |
| 0.094 | |
| 3 | CDL | + | + | + | − | −39 | + | 126 | ST11 |
| 0.064 | ||
| 19 | CAE | + | + | + | − | −39 | + | 126 | nd | 0.38 | 0.047 | ||
| 53 | CAE | + | + | + | − | −39 | + | 078 | nd | 0.50 | 0.094 | ||
| 1 | CAE | + | + | + | − | 0 | T |
| AI-53 | nd | 0.25 | 0.125 | |
| 81 | CDL | + | + | + | − | 0 | T | FR227 | nd | 0.38 | 0.19 | ||
| 42 | CAE | + | + | + | − | 0 | T | 020 | nd |
| 0.19 | ||
| 42 | CDL | + | + | + | − | 0 | T | 020 | nd | 0.50 | 0.125 | ||
| 43 | CAE | + | + | + | − | 0 | T | 020 | ST2 | 0.50 | 0.19 | ||
| 43 | CDL | + | + | + | − | 0 | T | 020 | 0.50 |
| |||
| 57 | CAE | + | + | + | − | 0 | T | 020 | nd | 0.50 | 0.125 | ||
| 78 | CAE | + | + | + | − | 0 | T | 020 | nd | 0.50 | 0.19 | ||
| 9 | CAE | + | + | + | − | 0 | T | 012 | ST54 | 0.50 | 0.047 | ||
| 85 | CAE | + | + | + | − | 0 | T | 012 | nd | 0.50 | 0.047 | ||
| 2 | CAE | + | + | + | − | 0 | T | 005 | nd |
| 0.125 | ||
| 29 | CDL | + | + | + | − | 0 | T | 005 | nd |
| 0.125 | ||
| 90 | CAE | + | + | + | − | 0 | T | 005 | nd | 0.25 | 0.047 | ||
| 12 | CAE | + | del | + | − | 0 | − |
| 017 | ST37 | 0.50 | 0.094 | |
| 32 | CAE | + | del | + | − | 0 | − | 017 | ST37 |
|
| ||
| 32 | CDL | + | del | + | − | 0 | − | 017 | ST37 |
| 0.19 | ||
| 36 | CAE | + | del | + | − | 0 | − | 017 | ST37 |
|
| ||
| 36 | CDL | + | del | + | − | 0 | − | 017 | ST37 |
| 0.19 | ||
| 37 | CAE | + | del | + | − | 0 | − | 017 | nd |
|
| ||
| 37 | CDL | + | del | + | − | 0 | − | 017 | ST37 |
|
| ||
| 92 | CAE | + | del | + | − | 0 | − | 017 | nd | 0.50 |
| ||
| 29 | CAE | + | − | − | + | − | − |
| 439 | nd | 0.50 | 0.047 | |
| 91 | CAE | + | − | − | + | − | − | 035 | nd | 0.125 | 0.125 | ||
| 5 | CAE | + | − | − | + | − | − | 009 | ST3 | 0.50 | 0.094 | ||
| 18 | CAE | + | − | − | + | − | − | 009 | ST3 |
| 0.125 | ||
| 18 | CDL | + | − | − | + | − | − | 009 | ST3 | 0.50 | 0.094 | ||
| 21 | CAE | + | − | − | + | − | − | 009 | nd | 0.125 | 0.094 | ||
| 60 | CAE | + | − | − | + | − | − | 009 | nd |
| 0.094 | ||
| 60 | CDL | + | − | − | + | − | − | 009 | nd | 0.50 | 0.064 | ||
| 61 | CDL | + | − | − | + | − | − | 009 | nd |
| 0.064 | ||
| 2018 | |||||||||||||
| 0 | CDL | + | del | + | − | 0 | − |
| 017 | nd | 0.38 | 0.032 | |
| Equidae number | Digestive content | Strain | Enrichment | Pathogenicity | Ribotype | GDH | Free toxins |
|---|---|---|---|---|---|---|---|
|
| CAE | − |
| 126 | + |
| |
|
| − |
| 126 | + |
| ||
|
| CAE | − |
| 017 | + | ||
|
| − |
| 017 | + | |||
|
| CAE | − |
| 017 | + |
| |
|
| − |
| 017 | + |
| ||
|
| CAE | − |
| 017 | + |
| |
|
| − |
| 017 | + |
| ||
| 29 | CAE | + | Non-Toxigenic | 439 | + (weak) | − | |
|
| − |
| 005 | + (weak) | − | ||
| 42 | CAE | + |
| 020 | − | − | |
|
| + |
| 020 | − | − | ||
| 43 | CAE | − |
| 020 | + | − | |
|
| − |
| 020 | + | − | ||
| 53 | CAE | + |
| 078 | − | − | |
|
| None | NA | NA | NA | − | − | |
| 54 | CAE | None | NA | NA | NA | − | − |
|
| Not preserved | − |
| ND | − | − | |
| 57 | CAE | − |
| 020 | − | − | |
|
| None | NA | NA | NA | − | − | |
| 78 | CAE | + |
| 020 | − | − | |
|
| None | NA | NA | NA | − | − | |
| 81 | CAE | None | NA | NA | NA | − | − |
|
| + |
| FR227 | − | − | ||
| 1 | CAE | + |
| AI-53 | − | − | |
| 2 | CAE | + |
| 005 | + (weak) | − | |
| 9 | CAE | − |
| 012 | + | − | |
| 12 | CAE | − |
| 017 | − | − | |
| 19 | CAE | − |
| 126 | − | − | |
| 85 | CAE | − |
| 012 | + | − | |
| 90 | CAE | + |
| 005 | − | − | |
| 92 | CAE | − |
| 017 | − | + (weak) | |
| 18 | CAE | − | Non-toxigenic | 009 | + | − | |
|
| − | Non-toxigenic | 009 | + | − | ||
| 48 | CAE | None | NA | NA | NA | − | − |
|
| Not preserved | − | Non-toxigenic | ND | − | − | |
| 60 | CAE | − | Non-toxigenic | 009 | + | − | |
|
| − | Non-toxigenic | 009 | + | − | ||
| 61 | CAE | None | NA | NA | NA | − | − |
|
| + | Non-toxigenic | 009 | + | − | ||
| 5 | CAE | − | Non-toxigenic | 009 | + | − | |
| 21 | CAE | − | Non-toxigenic | 009 | − | − | |
| 91 | CAE | − | Non-toxigenic | 035 | + | − |
- —IFCE
- —IFCE
- —Agence Nationale de Sécurité Sanitaire de l'Alimentation, de l'Environnement et du Travail (ANSES)
- —Institut National de Recherche pour l'Agriculture, l'Alimentation et l'Environnementhttp://dx.doi.org/10.13039/501100022077
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsClostridium difficile and Clostridium perfringens research · Yersinia bacterium, plague, ectoparasites research · Bacillus and Francisella bacterial research
INTRODUCTION
Clostridioides difficile is a spore-forming anaerobe responsible for diarrhea and colitis (1, 2). C. difficile, whose spores are transmitted by the fecal-oral route, is a major cause of healthcare-associated diarrhea and a rising cause of community-acquired infections (1, 2). C. difficile infections (CDIs) can be severe, with complications and recurrences, and life threatening (1, 2). Toxin A (TcdA) and Toxin B (TcdB) are responsible for symptoms, with TcdB alone being sufficient to cause disease. The binary toxin (CDT) has an accessory role (3). Non-toxigenic strains are unable to cause disease, while toxigenic strains can cause disease or be asymptomatically carried (4). Antibiotics, hospitalization, and age increase the risk of developing CDI (1, 2). Antibiotics notably induce microbiota dysbiosis, which favors spore germination, vegetative multiplication, and ultimately, toxin production (1, 2). Healthcare-associated CDIs are treated with fidaxomicin and vancomycin (2, 5), which can contribute to microbiota dysbiosis and favor C. difficile re-emergence, possibly from persisting biofilms (2, 6, 7). In this context, the last resort treatment is fecal microbiota transplant (FMT) (5).
C. difficile is a One Health issue (8–11). Spores, which are highly resistant, are widely disseminated by fecal spreading in the environment and can contaminate different hosts (8–12). C. difficile strains can be transmitted between host species (11, 13–15). Toxigenic strains can cause CDI in a wide range of wild, farm, and companion animals (9–12, 16, 17). Equidae, which are monogastric mammals, are closely interacting with humans and relevant to zoonotic and anthropo-zoonotic transmission. Importantly, in these animals of high economic value (French Institute for Horse and Riding [IFCE], https://www.ifce.fr), antibiotics treatments and hospitalization are common and increase the risk of developing CDI, as it is the case in humans. In Equidae*, C. difficile* is also responsible for outbreaks and sporadic cases of CDI (18–22). Both Equidae and humans are treated with vancomycin and have long been treated with metronidazole, which is still used in animals but replaced with fidaxomicin for hospitalized patients (2, 5). Like humans, Equidae can also be treated with FMT (23). The main difference between Equidae and humans as *C. difficile-*infected populations is age. While <3-years-old children are mainly asymptomatic carriers (4), CDI is common in foals (18, 19).
Data about the importance of C. difficile in horses (8, 10, 12, 20, 24) remain scarce in France. Here, we studied C. difficile in Equidae necropsied in Normandy, the leading horse breeding region in France (12,600 foals and 145,000 Equidae in 2021 according to IFCE: https://statscartes.ifce.fr/storage/files/1/pdf/IFCE-Depliant-chiffres-cles-2022-WEB.pdf). Necropsy (25) provided the opportunity both to recover digestive contents from a population with diverse clinical pictures and to get access to detailed animal data including the cause of animal death. We included all necropsied Equidae, whatever the cause of death and clinical picture, to test the presence of C. difficile and that of toxins when relevant. Our objective was to post-mortem evaluate C. difficile occurrence, diversity, putative circulation, and virulence in French Equidae.
MATERIALS AND METHODS
Equine necropsy
At the French National Agency for Food, Environmental and Occupational Health & Safety (ANSES), the PhEED Unit in Normandy is in charge of surveillance, prevention, and study of equine diseases. The veterinarian of PhEED is notably in charge of equine necropsies at the request of breeders or veterinarians to determine the cause of animal death. The standardized procedure includes examination of the cadaver, evisceration, and organ examination. Detailed animal data including identification (necropsy date, sex, age, breed, and location), ante-mortem information (treatments, including antibiotics for 2–3 weeks before death, and hospitalization) and post-mortem data (necropsy observations, standard analyses performed in case of signs of infection to identify pathogens, and the cause of animal death) are registered (https://sitesv2.anses.fr/fr/minisite/resumeq/presentation-de-resumeq) (25) (Table S1). Housing style was not recorded.
In 2018, we chose five necropsied Equidae with post-mortem signs of endo-enterotoxaemia and diarrhea and collected their watery intestinal contents for a pilot study. Then, from May 2019 to August 2021, we studied all (n = 100) necropsied animals (except fetus and stillborn foals), irrespective of clinical picture or cause of death. This total number, after 2 years characterized by Covid-19 and lockdowns, was relatively low, matching the previous annual average. Animals originated from Normandy (n = 97), Brittany (n = 1), and Centre Val de Loire (n = 1) (the origin of one animal was not registered) (Table S1). Animal digestive contents were systematically recovered in the cecum (CAE; n = 100), and in case of a watery consistency, in any other intestine segment (CDL; n = 30), notably the colon. Aliquots of all digestive contents except a cecal one (129 samples in 2019–2021) were preserved at 4°C, −20°C, and −70°C.
This study followed institutional guidelines and was approved by the scientific committee of IFCE that funded the project. All data are anonymous.
Standard pathogen testing
In case of post-mortem observed signs of infection, the veterinarian requested routine pathogen testing in relevant samples (Table S1). For intestinal infections, usual testing in digestive contents was as follows. Escherichia coli colonies, after detection on Eosin Methylene Blue plates, were streaked on blood plates to see hemolysis. Salmonella spp. were isolated on XLD plates (Thermo Scientific). Clostridial species were detected under anaerobiosis: C. difficile on ChromID (BioMérieux), Clostridium perfringens and Paeniclostridium sordellii on Columbia agar containing 5% sheep blood (BioMérieux). Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight (MALDI-TOF) Mass Spectrometry (MS) (MBT Smart mass spectrometer, Compass and FlexControl softwares, Bruker Daltonics) confirmed identification. Rotavirus was identified by PCR. Intestinal parasite communities were identified and quantified by visual inspection (26).
C. difficile testing, isolation, and characterization
C. difficile presence was independently tested by streaking a sample of any digestive content onto ChromID or CLO plates (BioMérieux) and incubating for 48 h at 37°C under anaerobiosis. In case of a negative result, a 1 g-aliquot was harvested after sample homogenization, inoculated into a rich, permissive medium: BHI broth supplemented by cysteine (0.1%), yeast extract (5 g/L), taurocholate (1 g/L), D-cycloserine (250 µg/mL), and cefoxitin (8 µg/mL) (OXOID), and finally incubated for 3–7 days (enrichment) before plating onto ChromID. A positive stool was included as a control. Suspicious, irregular, and/or black colonies were screened by PCRs targeting tpi, tcdA, and tcdB genes (27, 28). Identification was confirmed by MALDI-TOF MS. A single C. difficile colony per positive sample was then grown in BHI broth at 37°C under anaerobiosis to be preserved in glycerol (17%) at −70°C, except in two cases: two strains (non-toxigenic in animal #48 and toxigenic in #54) characterized without being preserved, could not be recovered later.
All preserved isolates (N = 34 from n = 25 Equidae in 2019–2021 and N = 1 in 2018) were characterized by multiplex PCR targeting: (i) tpi gene, (ii) wild-type or truncated tcd and cdt genes, and (iii) lok fragment characterizing non-toxigenic strains, followed by capillary gel electrophoresis (29). For ribotyping, PCR profiles after capillary gel-based electrophoresis (30) were analyzed with GeneMapper software (Thermo Fischer Scientific, Villebon-sur-Yvette, France) and ribotypes assigned according to the WEBRIBO database (https://webribo.ages.at/). The first isolates were characterized by Multi Locus Sequence Typing (MLST) (31).
Susceptibility to vancomycin and to metronidazole
The Minimum Inhibitory Concentration (MIC) was determined using Etest (BioMérieux) according to the EUCAST and CA-SFM recommendations when the experiments were performed (https://www.sfm-microbiologie.org/wp-content/uploads/2021/04/CASFM2021__V1.0.AVRIL_2021.pdf). Briefly, a suspension of 1.0 MacFarland was prepared in Schaedler broth and spread on a pre-reduced Brucella blood agar (Becton Dickinson) supplemented with vitamin K1 (10 mg/L), hemin (5 mg/L), and defibrinated horse blood (5%) (OXOID). Of note, Brucella blood agar medium was recently published to be suboptimal for Metronidazole Etests (32). A strip, in which antibiotic concentration ranged from 0.0016 to 256 mg/L, was laid on the plates, which were then incubated for 24h (vancomycin) or 48h (metronidazole) at 37°C under anaerobiosis (using Thermo Scientific Oxoid AnaeroGen Compact sachets). We included C. difficile reference strain ATCC 700057 as a control.
Toxin detection
C. difficile toxin A, toxin B, and glutamate dehydrogenase (GDH) were detected in digestive contents using an Enzymatic Immuno-Assay (Quick Check Complete, Alere) according to manufacturer’s instructions. For each newly opened kit, the supplied positive control was tested. For each series of experiments, we also included an aliquot of a human stool known to be positive and stored at −80°C. A second independent observation confirmed any weak positive result.
Statistical analysis
We used a Fisher’s exact test to analyze the equine population composition, after excluding unknown values, and a Mann-Whitney test to compare strain MICs.
RESULTS
Necropsied equidae
We studied all Equidae (n = 100) necropsied at ANSES in Normandy from May 2019 to August 2021, without any selection (Table S1). They belonged to 78 premises, all located in Normandy except three (Table S1). Most were females (67%) and <1-year-old animals (59%), with foals (< 6 months) representing half of the population (48%) (Table 1). Nine breeds were present, mostly Thoroughbred (42%), Trotters (31%), and Saddlebred horses (11%) (Table 1). Animals had died from very diverse (> 30) causes including infectious or non-infectious diseases and trauma (Table S1). The first and second most prevalent causes of death were (i) endo-enterotoxaemia (12%), an enteric disease caused by toxin-producing bacteria like Salmonellae and Clostridia (21), and (ii) infection by Rhodococcus equi (9%), common in foals (33).
C. difficile prevalence
Animal cecal contents (99 CAE) and watery intestinal contents (30 CDL) were tested for C. difficile, directly or after enrichment. After screening by standard PCRs (27, 28) and confirmation by MALDI-TOF, twenty-seven animals were positive for C. difficile, in their two samples in nine cases (Table S1). C. difficile presence in Equidae was independent of animal sex, age, breed, antibiotic treatment or hospitalization, but significantly correlated to signs of diarrhea or endo-enterotoxaemia (Table 1, Fig. S1).
Pathogenicity
Thirty-four C. difficile isolates were preserved from twenty-five Equidae (Table 2). They form the first French library of C. difficile strains originating from Equidae, CloDifEqui (Table 2). Their pathogenicity profile was characterized by multiplex PCR, which revealed non-toxigenic isolates (N = 9) and toxigenic ones (N = 25) with three profiles (tcdA^+^ tcdB^+^ cdt^+^, tcdA^+^ tcdB^+^ or tcdB^+^) (Table 2). The former isolates originated from seven animals, including six Thoroughbred, and the latter from nineteen animals of six breeds (Table S1). One animal (#29) was co-colonized by a non-toxigenic strain (CAE) and a toxigenic one (CDL). In the eight other animals with two isolates, both were either non-toxigenic (#18, #60) or toxigenic with the same profile (#3, #32, #36, #37, #42, #43) (Table 2).
Ribotype diversity
CloDifEqui isolates were assigned to 11 ribotypes and 4 clades (1, 2, 4, and 5) (Table 2; Table S2) (30). Toxigenic isolates were of eight ribotypes: 126 and 078 (tcdA^+^ tcdB^+^ cdt^+^ and the same tcdC truncation), 020, 005, 012, AI-53, and FR227 (tcdA^+^ tcdB^+^), and 017 (tcdB^+^). Non-toxigenic isolates belonged to ribotypes 009, 035, and 439. The two most prevalent ribotypes (n = 5 animals) were 017 (N = 8 isolates) and 009 (N = 7).
The two strains of animal #29, which were toxigenic and non-toxigenic, belonged to ribotype 005 and 439. In all other pairs of isolates originating from the same animal, both were of the same ribotype (Table 2).
Susceptibility to metronidazole and to vancomycin
All strains were susceptible to the antibiotics used to treat CDI in Equidae, metronidazole and vancomycin (MICs lower than EUCAST breakpoints: 2 mg/L in both cases; https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_15.0_Breakpoint_Tables.pdf) (Table 2). For metronidazole, strain MICs ranged from 0.032 to 0.38 mg/L, with a median value of 0.13 mg/L. For vancomycin, strain MICs ranged from 0.13 to 1 mg/L, with a median value of 0.5 mg/L. Strains belonging to ribotypes 017 and 126 showed the highest MIC for metronidazole (0.38 mg/L) and for vancomycin (1.0 mg/L), respectively.
Co-location of Equidae sharing the same ribotype
We cross-analyzed Equidae location and C. difficile ribotypes (Fig. 1; Table S1). In two premises between Caen and Pont-Audemer, some animals shared the same ribotype: (i) 020, for two co-located Equidae necropsied in 2019 (#42 and #43) and (ii) 009 for four co-located Equidae necropsied in 2019 (#5, #18, and #21) and 2020 (#61). This suggested (i) that in each premises, isolates of the same ribotype might be clonal and (ii) that transmission between animals or from a common environmental source might have occurred.
Geographical location of C. difficile-positive Equidae. All C. difficile-positive Equidae are shown on a map of Normandy according to the position of their stud farm (Table S1). The premises of each animal is indicated by a symbol depending on C. difficile toxin gene profile (multiplex PCR): squares for tcdA+ tcdB+ cdt+, circles for tcdA+ tcdB+, triangles for tcdB+, and diamonds for non-toxigenic strains (Table 2). C. difficile ribotype (Table 2) is indicated in a box, and when several animals harbored this ribotype in the same premises (Table S1), their number appears between brackets. C. difficile pathogenicity and virulence are represented using a color code: (i) green for non-toxigenic, (ii) orange for toxigenic and carried, and (iii) red for toxigenic and virulent (having produced toxins). The map was generated in RStudio using the ggmap package, using underlying geographic data from OpenStreetMap.
Toxin production
We tested the presence of free toxins in equine digestive contents using a standard enzymatic immuno-assay, initially developed for human fecal materials, but also used in animals, notably foals (34). To check the assay specificity when applied to equine digestive contents, we included samples expected to be toxin-negative: (i) all 10 samples displaying a non-toxigenic C. difficile (Table 3) and (ii) 27 C. difficile-negative samples (15 CDL and all samples from animals with endo-enterotoxaemia signs) (Table 3; Table S1 and Table S3). No toxins could be detected in any of them, while GDH was positive in seven of the former (Table 3; Table S3).
Free toxins and GDH were detected in the two samples of four animals (#3, #32, #36, and #37), indicating C. difficile virulence (Table 3). These animals displayed signs of diarrhea (CDL in the colon) and a toxigenic C. difficile (ribotype 126 or 017) was identified as the only enteropathogen (Table 3, Fig. 2). These data were highly suggestive of animal CDI. The 16 remaining Equidae with a toxigenic C. difficile, in which no toxin was detected (Table 3), were carriers. Half of them showed no sign of diarrhea (#1, #2, #9, #12, #19, #85, #90, and #92), suggesting asymptomatic carriage. For almost all others (except #57), signs of diarrhea might be due to another identified intestinal pathogen (C. perfringens in #43, C. perfringens or P. sordellii in #29 and #42, C. perfringens, P. sordellii or a rotavirus in #53, and a parasite in #78) or to the non-infectious intestinal disease having caused death (torsion of the large colon in #54 and intestinal lymphoma in #81) (Table S1).
C. difficile virulence or carriage in C. difficile-positive Equidae. All C. difficile-positive animals are shown as a wedge of a pie chart. For each positive animal, the pathogenicity, ribotype, and virulence of C. difficile are indicated in four concentric rings. First (core): C. difficile pathogenicity (Table 2). Non-toxigenic and toxigenic strains are indicated on a green or orange background, respectively. “Both” is indicated when a non-toxigenic strain and a toxigenic strain were co-colonizing the same animal (#29, bi-colored wedge). The toxin gene profile of toxigenic strains is specified using the following abbreviations: A for tcdA+, B for tcdB+, C for cdt+ and “?” when the presence of cdt genes has not been tested. Second: C. difficile ribotype (according to the WEBRIBO database) (Table 2). Third: Animal identification number. Fourth: Antibiotic treatment in the 2–3 weeks before death (“+”) or not. Fifth (outer ring): Virulence (Toxin production, Table 3) or carriage. Virulence, carriage of a toxigenic strain, and carriage of a non-toxigenic strain are on a red, orange, and green background, respectively.
Among Equidae with a toxigenic C. difficile, we compared those in which toxins were detected to the others. These two subpopulations were similar in terms of sex, age, and non-pathognomonic signs of endo-enterotoxaemia or diarrhea, but they significantly differed in terms of breed and antibiotic treatment (Table 1). Three out of 4 toxin-positive animals (all Trotters) compared to 3 out of 16 carriers of a toxigenic C. difficile had received antibiotics (Table 1, Fig. 2; Table S1). In the subpopulation with a toxigenic C. difficile, toxins were, therefore, produced in half of the antibiotic-treated animals (3/6), but in none of the untreated ones (0/12) (Table 1, Fig. 2). Antibiotic treatments could, therefore, have increased the risk for Equidae to develop CDI.
C. difficile as the most probable cause of animal death
We examined whether C. difficile could have caused the death of the four toxin-positive animals, by analyzing available data (Table S1). The fulminant death of animal #3, a 15-day-old foal, was not primarily due to C. difficile but to small intestine intussusception (Table S1). Animal #37, an adult, died from multiple infections (respiratory, musculoskeletal and digestive infections, including endo-enterotoxaemia), probably due to Klebsiella pneumoniae, which was identified in lungs, lung abscess and hock muscle, and/or C. difficile (Table S1).
For the last two animals, #32 and #36, both foals, death was due to endo-enterotoxemia (Table S1 and Fig. S2). C. difficile was the only identified enteropathogen (Table S1) and considering the presence of toxins (Table 3), the most probable cause of death. The resulting prevalence of C. difficile as the cause of animal death (2%) was of the same range as the previously measured ones (0%–5%). Of note, ribotype 017 was implicated in both cases and likely also in one case of the 2018 pilot study: animal #0, which died from endo-enterotoxaemia following antibiotic treatment, displayed signs of diarrhea and a C. difficile of ribotype 017 as the only enteropathogen (Table 2; Table S1).
Discussion
C. difficile prevalence, diversity, and epidemiology
Here, we evaluated the presence of C. difficile in Equidae necropsied in France for the first time. C. difficile prevalence in these Equidae having died from diverse, mainly non-intestinal diseases (Table S1), was 27%. It was similar to, and sometimes lower than, the prevalence reported in live horses in Europe (4%–33% for healthy animals and 5%–63% for sick animals with intestinal illnesses) (10) or in Australia (31% for animals with diarrhea or not) (35). The prevalence of toxigenic strains was also similar in French Equidae (20%) and Australian horses (18%) (35).
C. difficile strains from Equidae belonged to eleven already described ribotypes (10, 36). Toxigenic strains were of eight ribotypes. The historical epidemic ribotype 027 (1), described in horses (20), was not identified here, probably because of the changing C. difficile epidemiology and 027 prevalence decrease (36–38). Most ribotypes of CloDifEqui toxigenic strains are common in humans: ribotypes 020, 078, 126, and 005 were in the Top 10 ribotypes in European patients in 2013–2014, and ribotypes 017 and 012 in the Top 20 (36). In Czech horses (n > 200), toxigenic strains were of only five ribotypes (24), including one (012) in the Top 20 (36). In horses in Australia (n ~ 400), toxigenic strains belonged to 40 ribotypes, including 18 internationally known ribotypes like 012, 014/020, and 087, locally identified and novel ribotypes (35). Finally, our study revealed that French Equidae harbored diverse, already described, and mostly clinically important ribotypes.
Most ribotypes of CloDifEqui toxigenic strains are also common in animals (10, 12). Ribotypes 126, 020, and 012 had been found in farm animals (10). Ribotype 078, known as prevalent in animals, notably pigs (10), was relatively uncommon here (N = 1 isolate), but the phylogenetically close ribotype 126 was represented (N = 3) (Table 2) (39). Of note, the ribotypes of most CloDifEqui toxigenic strains have been detected in very diverse sources in Europe, including mollusks (005, 012, 017, 020, 078, 126, and AI-53) (40), potatoes, and vegetables (012, 020, 078, and 126) (12, 41).
The diversity and ubiquity of ribotypes identified here supported a role for Equidae in the cycle of C. difficile transmission in France, which is important in a One Health perspective (11, 20). Many different environmental, animal or human sources could have contaminated French Equidae during their life. In turn, they could have contributed to C. difficile dissemination and transmission to other hosts of the same or other species including humans. The cross-analysis of Equidae location and C. difficile ribotypes revealed that in two cases, animals of the same premises displayed the same ribotype, indicating strain phylogenetic proximity and raising the possibility of a single clonal lineage. Each of the two premises might represent a cluster of contamination and of transmission between Equidae, either directly or via a common source. Genome-wide investigations are under progress to analyze the phylogeny of C. difficile isolates originating from Equidae (CloDifEqui collection) and from patients of the same region during the same period. This should help understand C. difficile circulation in Normandy and evaluate zoonotic or anthropo-zoonotic transmission, an important One Health issue.
Toxin production
After PCR amplification of toxin genes, a method of high sensitivity and negative prediction value, we performed toxin detection, a method of high specificity and positive prediction value, as recommended for CDI diagnosis in European patients (4), but rarely performed in animals including Equidae (42–45). Four animals with both signs of diarrhea and a toxigenic C. difficile were positive for toxins, suggesting in vivo (ante-mortem) virulence and CDI in 4% of Equidae. Carriers of a toxigenic C. difficile represented 16% of the population (Table 3, Fig. 2), a high and previously overlooked level.
In Equidae, C. difficile presence (Table 1; Table S1) was significantly correlated to animal signs of diarrhea or endo-enterotoxemia. In C. difficile carriers, in the absence of toxins, these signs were unexpectedly more frequent than in negative animals (12/23 vs 15/75). However, the former, compared to the latter, also more frequently harbored another enteropathogen (bacterium, virus, parasite) (7/23 vs 9/73) (Table S1), which might be responsible for diarrhea. The most prevalent was C. perfringens in four animals, three of which also harbored P. sordelli (Table S1). Two carriers of a toxigenic C. difficile died from endo-enterotoxaemia, which might have been caused by (i) C. perfringens (#43) or (ii) C. perfringens and/or P. sordellii (#42) (Table S1 and Fig. S2). Host co-colonization or co-infection by C. difficile and C. perfringens have been described in horses (46, 47), dogs (48), pigs (49), and humans (50).
In this study, the most complex case was animal #29, which displayed three clostridial enteropathogens and two C. difficile strains, one toxigenic and the other non-toxigenic (Table S1; Table 2). Co-colonization by a toxigenic strain and a non-toxigenic one, notably of ribotype 009, has been reported in companion animals (51). Non-toxigenic strains are able to prevent CDI or reduce its severity in animal models (52) and a ribotype 009 strain can protect both model and commercial piglets (49). Whether CloDifEqui non-toxigenic strains, especially of ribotype 009, could provide protection against a toxigenic strain would require further investigations.
C. difficile had produced toxins in four Trotters and at least three had received antibiotics (Table 1; Table S1). Of note, among the eight C. difficile carriers having received antibiotics, five displayed non-toxigenic strains unable to cause disease (Fig. 2). Antibiotic treatments are known to favor microbiota dysbiosis, C. difficile outgrowth and ultimately, virulence (2). Antibiotic treatments alter the metabolic functions of the microbiota, resulting in the loss of colonization resistance by several mechanisms. One of them is the accumulation of primary bile salts, which favor spore germination and do not inhibit vegetative multiplication (53).
The C. difficile strains that had produced toxins in Equidae were of ribotypes 126 and 017. Ribotype 126 was the sixth most prevalent ribotype in European patients in 2013–2014 (36). It is also, together with its close relative 078, prevalent in production animals, notably pigs (10, 54), and in biogas plants using animal manure in another French region, Brittany (55). Ribotype 017 is prevalent in patients in Asia (56), common in Europe (36), notably in Greece (2), but less frequent in France, where it was undetected in 2022 (36, 38). Ribotype 017 is also described as relatively uncommon in animals, notably compared to ribotype 078 (10, 56). For example, in Japan, in 34 horses with CDI, ribotype 017 was clearly less prevalent than ribotype 078 (< 10% vs 35%) (57). As ribotype 017 was not detected in biogas plants in Brittany (55), its prevalence in French Equidae (5% of the whole population and 25% of the subpopulation with a toxigenic C. difficile) was noticeable. Moreover, its virulence in three close animals (Fig. 1) within a short period (Table S1) raised the possibility of a local infection cluster.
In conclusion, CDI likely affected 4% of the French Equidae under study before their death, in most cases after an antibiotic treatment. Ribotype 017, which was unexpectedly the most prevalent ribotype, was also the most virulent and should be carefully monitored in the future. Moreover, as many as 20% of animals were able to disseminate toxigenic C. difficile. French Equidae, therefore, represented an underestimated reservoir of clinically important C. difficile, with a risk of zoonotic transmission and contribution to community-acquired infections. Our findings highlight the importance of a One Health perspective in the epidemiological surveillance of C. difficile.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Smits WK, Lyras D, Lacy DB, Wilcox MH, Kuijper EJ. 2016. Clostridium difficile infection. Nat Rev Dis Primers 2:16020. doi:10.1038/nrdp.2016.2027158839 PMC 5453186 · doi ↗ · pubmed ↗
- 2Chilton CH, Viprey V, Normington C, Moura IB, Buckley AM, Freeman J, Davies K, Wilcox MH. 2025. Clostridioides difficile pathogenesis and control. Nat Rev Microbiol. doi:10.1038/s 41579-025-01242-2 · doi ↗
- 3Kordus SL, Thomas AK, Lacy DB. 2022. Clostridioides difficile toxins: mechanisms of action and antitoxin therapeutics. Nat Rev Microbiol 20:285–298. doi:10.1038/s 41579-021-00660-234837014 PMC 9018519 · doi ↗ · pubmed ↗
- 4Crobach MJT, Planche T, Eckert C, Barbut F, Terveer EM, Dekkers OM, Wilcox MH, Kuijper EJ. 2016. European society of clinical microbiology and infectious diseases: update of the diagnostic guidance document for Clostridium difficile infection. Clin Microbiol Infect 22 Suppl 4:S 63–81. doi:10.1016/j.cmi.2016.03.01027460910 · doi ↗ · pubmed ↗
- 5van Prehn J, Reigadas E, Vogelzang EH, Bouza E, Hristea A, Guery B, Krutova M, Norén T, Allerberger F, Coia JE, Goorhuis A, van Rossen TM, Ooijevaar RE, Burns K, Scharvik Olesen BR, Tschudin-Sutter S, Wilcox MH, Vehreschild MJGT, Fitzpatrick F, Kuijper EJ, Guideline Committee of the European Study Group on Clostridioides difficile. 2021. European society of clinical microbiology and infectious diseases: 2021 update on the treatment guidance document for Clostridioides difficile infection in adul · doi ↗
- 6Frost LR, Cheng JKJ, Unnikrishnan M. 2021. Clostridioides difficile biofilms: a mechanism of persistence in the gut? P Lo S Pathog 17:e 1009348. doi:10.1371/journal.ppat.100934833705497 PMC 7951904 · doi ↗ · pubmed ↗
- 7Poquet I, Saujet L, Canette A, Monot M, Mihajlovic J, Ghigo JM, Soutourina O, Briandet R, Martin-Verstraete I, Dupuy B. 2018. Clostridium difficile biofilm: remodeling metabolism and cell surface to build a sparse and heterogeneously aggregated architecture. Front Microbiol 9:2084. doi:10.3389/fmicb.2018.0208430258415 PMC 6143707 · doi ↗ · pubmed ↗
- 8Lim SC, Knight DR, Riley TV. 2020. Clostridium difficile and one health. Clin Microbiol Infect 26:857–863. doi:10.1016/j.cmi.2019.10.02331682985 · doi ↗ · pubmed ↗