The cell cycle-regulated cytoplasmic kinase, TgCRCK1, is required for efficient propagation of human protozoan pathogen, Toxoplasma gondii
Dima Hajj Ali, Ramu Anandakrishnan, Frank O. Aylward, Rajshekhar Y. Gaji

TL;DR
This study identifies TgCRCK1, a cell cycle-regulated kinase in Toxoplasma gondii, as important for parasite growth but not for causing disease in mice.
Contribution
The study characterizes TgCRCK1, a previously uncharacterized kinase in Toxoplasma gondii, revealing its role in parasite propagation.
Findings
TgCRCK1 is a cytosolic kinase with cell cycle-dependent expression.
Conditional depletion of TgCRCK1 impairs parasite growth due to defects in division and invasion.
TgCRCK1 is not essential for virulence in a mouse model of toxoplasmosis.
Abstract
Toxoplasma gondii is an obligate intracellular parasite that relies on a complex network of protein kinases to regulate essential processes such as invasion, replication, and egress. The Toxoplasma genome encodes approximately 159 kinases, yet only a small subset has been characterized. Our group is interested in defining the role of cell cycle-regulated kinases important for Toxoplasma fitness. In this study, we investigated the role of an uncharacterized, cell cycle-regulated cytoplasmic kinase, which we named TgCRCK1, in parasite biology and pathogenesis. Using endogenous tagging and immunofluorescence assays, we demonstrated that TgCRCK1 is a cytosolic kinase exhibiting a cell cycle-dependent expression pattern. Conditional depletion of TgCRCK1 via an auxin-inducible degron system impaired parasite growth, primarily due to defects in cell division and invasion. Transcriptomic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
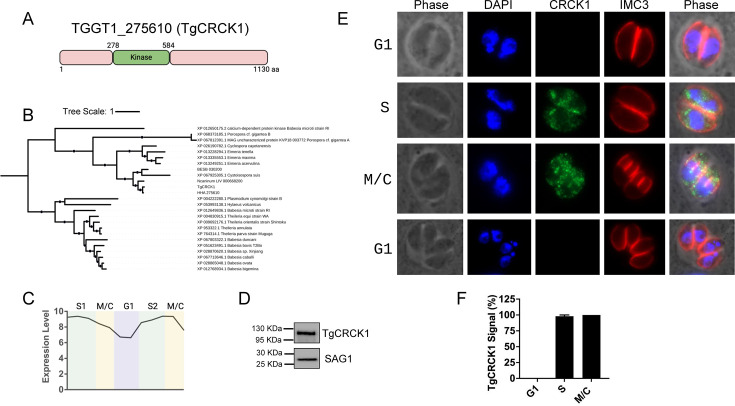 Fig 1
Fig 1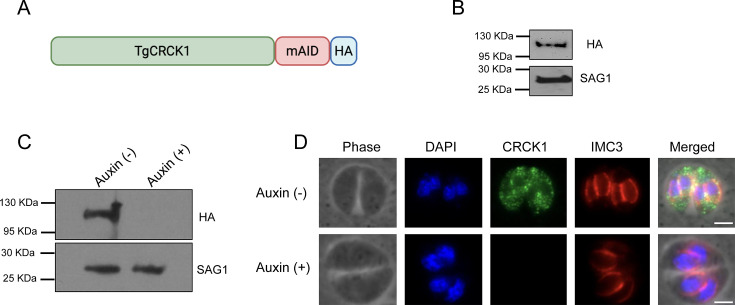 Fig 2
Fig 2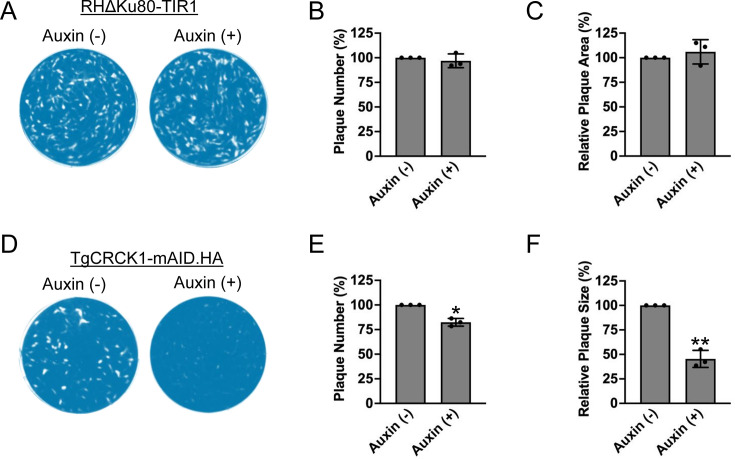 Fig 3
Fig 3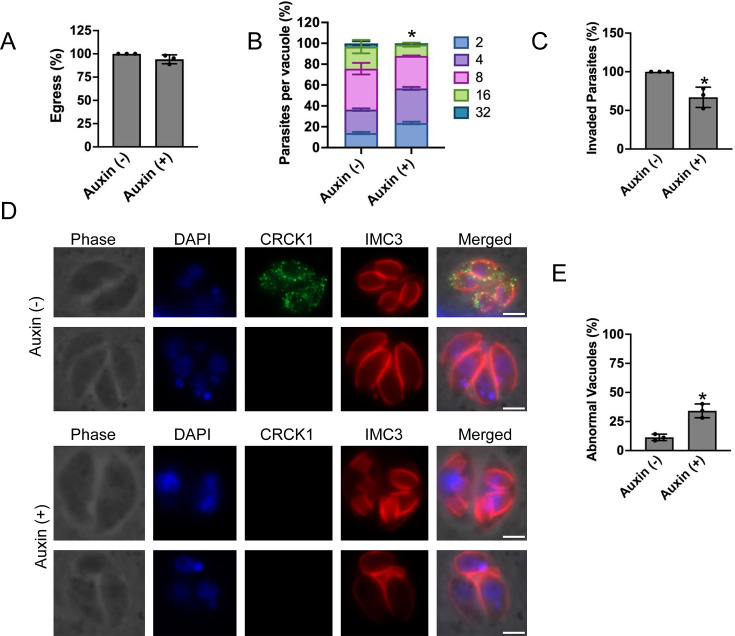 Fig 4
Fig 4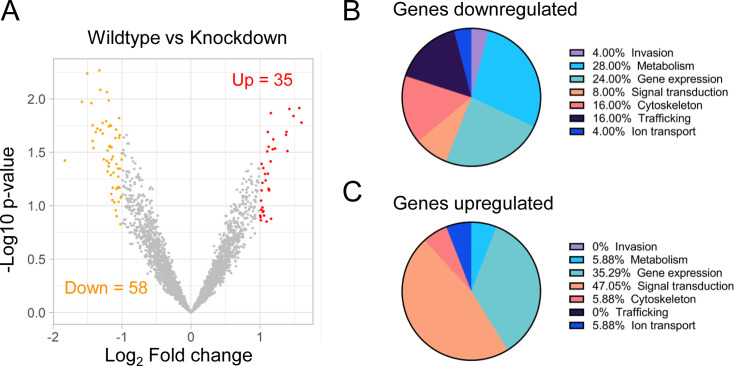 Fig 5
Fig 5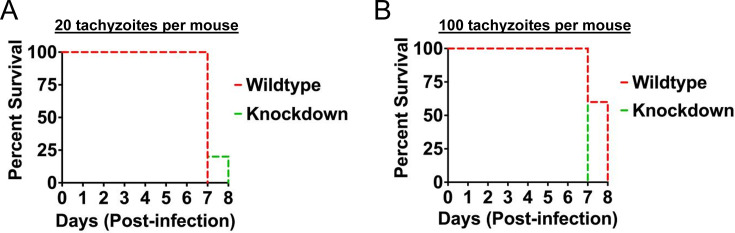 Fig 6
Fig 6- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Trypanosoma species research and implications · Amoebic Infections and Treatments
INTRODUCTION
Toxoplasma gondii, the causative agent of toxoplasmosis in humans, is a unicellular eukaryote classified within the phylum Apicomplexa (1, 2). This phylum includes several notable human and animal pathogens, such as Plasmodium, Cryptosporidium, Babesia, and Sarcocystis spp. (3). Cats act as the definitive hosts of Toxoplasma as the sexual phase of the life cycle of the parasite occurs in the intestinal epithelial cells of this species (4). The parasite is excreted in cat feces as oocysts, which act as the source of infection for intermediate hosts, primarily farm animals. Humans acquire Toxoplasma infection through multiple routes, including ingestion of contaminated meat from infected intermediate hosts, consumption of food or water contaminated with parasite oocysts, or via transplacental transmission from mother to fetus during pregnancy. In individuals with healthy immune systems, Toxoplasma infection is typically asymptomatic. However, the disease can be fatal in immunocompromised individuals, including cancer patients undergoing immunosuppressive therapy and HIV-positive persons. Furthermore, primary exposure to Toxoplasma during pregnancy can result in miscarriage or congenital abnormalities in the newborn (1, 2, 5–7). Currently, no vaccine exists to prevent Toxoplasma infection, and the drugs used to treat the acute form of the disease are associated with adverse side effects (8).
The pathology of toxoplasmosis primarily results from extensive tissue damage caused by the rapid and prolific multiplication of the parasite within infected individuals (8, 9). Specifically, Toxoplasma, an obligate intracellular pathogen, actively invades host cells and resides within a protective parasitophorous vacuole. The parasite then replicates by endodyogeny, a process in which two daughter parasites bud within the mother parasite (10–12). Toxoplasma continues to multiply until the host cell becomes filled with tachyzoites. Approximately 2 days post-infection, the newly formed daughter cells egress, leading to rupture of the infected host cell and subsequent invasion of neighboring cells. Because the infected host cell is destroyed during parasite egress, this intracellular replication process is commonly referred to as the lytic cycle (8, 9). Since the events of the lytic cycle—invasion, replication, and egress—are critical for the efficient propagation and survival of Toxoplasma within the host, identifying and characterizing parasite factors essential for Toxoplasma growth has become a priority for the development of novel therapeutics (8, 13, 14).
Kinases play critical roles in diverse cellular processes within eukaryotic cells, including regulation of gene transcription, translation, metabolism, cell division, motility, and responses to environmental signals. The Toxoplasma genome contains approximately 8,000 genes, of which 159 are predicted to encode kinases, representing about 2% of the genome (15). Members of this kinase family have been shown to be essential for key Toxoplasma processes, such as invasion, motility, cytoskeletal organization, endodyogeny, egress, and gene regulation (16–21). Although significant progress has been made in mining the Toxoplasma genome to understand the roles of kinases in parasite biology, the precise functions of many kinase family members remain largely undetermined.
In Toxoplasma, kinases have been found in different organelles, including the parasite nucleus, cytoplasm, plasma membrane, rhoptries, apicoplast, dense granules, pellicle, and the mitochondrion (15, 16, 20, 22–29). Although a large number of these kinases are constitutively expressed, some of these exhibit cell cycle regulation (20). A notable group of cell cycle-regulated kinases belongs to the Cdk-related kinase (CRK) family, which localizes to the parasite nucleus and plays a critical role in cell cycle progression (30). Other Toxoplasma kinases with periodic expression include TgMAPK1, TgMAPK2, TgARK1-3, TgNEK1, and TgTKL4; interestingly, many of these have been shown to be involved in parasite cell division (18, 31–36).
Our group aims to understand the role of cell cycle-regulated kinases that may be important for Toxoplasma growth and pathogenesis. In this study, we provide initial insights into the function of Toxoplasma gondii Cell-cycle Regulated Cytoplasmic Kinase 1 (TgCRCK1), a kinase predicted to be regulated during the cell cycle and important for parasite fitness (20). We show that the protein localizes to the parasite cytoplasm and is temporally regulated with exclusive expression in S and M/C phases. Phenotypic analysis of TgCRCK1 showed that its absence results in impaired parasite growth that is caused by defects in endodyogeny and reduced invasion competence. Furthermore, transcriptomic analysis suggested that loss of TgCRK1 results in dysregulation of gene expression profile in the parasite. However, despite its critical requirement for efficient parasite growth in vitro, TgCRCK1 depletion did not attenuate the parasite’s virulence in a murine infection model. Collectively, these findings provide novel insights into the role of this cell cycle-regulated kinase in Toxoplasma biology.
RESULTS
TGGT1_275610 is a cytoplasmic kinase expressed during the S and M/C phases of tachyzoite cell cycle
TGGT1_275610 is a Toxoplasma gene encoding a 1,130-amino acid protein with a kinase domain located in the distal region of the proximal half of the protein (Fig. 1A). Phylogenetic analysis (37) indicated that this kinase is highly conserved among members of the Eimeriidae family of apicomplexan parasites (Hammondia hammondi, Besnoitia besnoiti, Cystoisospora suis, and Neospora caninum) and exhibits only weak homology to piroplasmids, such as Plasmodium, Babesia, and Theileria (38). Notably, within the Eimeriidae, the kinase shows the greatest orthology with Hammondia species (Fig. 1B).
TgCRCK1 is a cell cycle-dependent cytoplasmic kinase. (A) Schematic representation of TgCRCK1 domain architecture, highlighting the relative position of the kinase domain. (B) Phylogenetic analysis of TgCRCK1 among related apicomplexan parasites. The alignments were generated using Muscle 5.1, and IQ-TREE v. 2.2.2.7 was used to produce the tree using the LG+F+R10 model (37). (C) Transcriptomic profile of TgCRCK1 across different cell cycle stages (39). (D) Western blot analysis of TgCRCK1.HA strain using an anti-HA antibody. TgSAG1 is used as a loading control. (E) Immunofluorescence analysis showing the localization of TgCRCK1 in intracellular parasites at various cell cycle stages using anti-HA antibody. TgIMC3 (red), a marker for the pellicle, was used to identify different cell cycle stages. DAPI was used to stain the nuclei. Scale bar, 2 μm. (F) Quantification of parasitophorous vacuoles in different cell cycle stages that showed TgCRCK1 signal. Data are presented as mean ± standard deviations from three independent experiments.
A previous transcriptomic study suggested that TGGT1_275610 is a cell cycle-regulated kinase, with peak expression during the S and M/C phases (40) (Fig. 1C). To determine its localization, we endogenously tagged TGGT1_275610 at the C-terminus with a hemagglutinin (HA) epitope using CRISPR/Cas9 technology (41). Western blot analysis with an anti-HA antibody detected a single band of the expected size (~118 kDa) in the tagged clone (Fig. 1D).
Next, we wanted to determine the localization of this kinase in the parasite, and hence, we performed immunofluorescence analysis of intracellular parasites. The results revealed that TgGT1_275610 localizes to parasite cytoplasm. Interestingly, the protein was not observed during the G1 stage of the parasite cell cycle. However, the protein makes an entrance in the S phase of the cell cycle, persists during M/C stages, and once parasites return to G1 phase, the protein again disappears (Fig. 1E). This stage-specific pattern was further supported by quantification of vacuoles exhibiting a TgTCRCK1 signal (Fig. 1F). We also observed that during the M/C phases, some fraction of the protein showed co-localization with the inner membrane complex (IMC) regions of the newly forming daughter cells (Fig. 1D).
Altogether, these results suggest that TGGT1_275610 is indeed a temporally regulated cytoplasmic kinase and its expression is confined to the S and M/C phases during tachyzoite cell cycle. Since this is a developmentally regulated kinase found in the parasite cytoplasm, we named this protein TgCRCK1.
Generation of conditional knockdown strain of TgCRCK1
Next, we wanted to determine the role of TgCRCK1 in Toxoplasma propagation. Since a previous study that conducted genome-wide analysis to identify genes important for Toxoplasma fitness suggested that TgCRCK1 is an essential gene with a mean phenotype score of −4.25 (40), we wanted to generate a conditional knockdown mutant of this gene. Toward this goal, we used the recently developed auxin-inducible degron (AID) system (42). This system involves tagging the C-terminus of TgCRCK1 with a mini-AID (mAID) domain followed by an HA epitope in a parasite line stably expressing the auxin receptor TIR1 (42). In the absence of auxin, the mAID-tagged TgCRCK1 functions normally; however, upon auxin addition, the mAID tag targets the protein for ubiquitination and subsequent proteasomal degradation.
Accordingly, we introduced the mAID-HA tag at the C terminus of TgCRCK1 by CRISPR-Cas9 technology to generate the TgCRCK1.mAID.HA strain as described in the Materials and Methods (Fig. 2A). Immunoblotting using anti-HA antibody revealed a single band of expected size for TgCRCK1 protein (Fig. 2B). Immunofluorescence analysis (IFA) confirmed that TgCRCK1.mAID.HA localizes to the parasite cytoplasm, consistent with our previous observation using the HA tag alone (Fig. 2D, top panel). To assess downregulation, parasites were treated with vehicle control or auxin, followed by immunoblotting. The results showed that TgCRCK1 completely disappears as early as 2 h in the presence of auxin (Fig. 2C). We also performed IFA analysis of TgCRCK1.mAID.HA strain using anti-HA antibody. We observed complete loss of TgCRCK1 protein in the parasites treated with auxin (Fig. 2D, bottom panel), thus suggesting successful establishment of conditional knockdown strain of TgCRCK1 protein.
Establishment of TgCRCK1 conditional knockdown strain using the AID system. (A) Schematic representation of TgCRCK1 with the mAID-HA tag at the C terminus. (B) Immunoblot analysis of TgCRCK1.mAID.HA strain using anti-HA antibody. TgSAG1 is used as the loading control. (C) Western blot analysis of intracellular TgCRCK1.mAID.HA strain (treated with either vehicle control or auxin for 2 h) using an anti-HA antibody. TgSAG1 serves as the loading control. (D) Immunofluorescence analysis of TgCRCK1.mAID.HA parasites (treated with either vehicle control or auxin for 2 h) using anti-HA antibody. TgIMC3 used as an IMC marker. Scale bar, 2 μm.
TgCRCK1 is important for parasite growth in vitro
To assess the impact of TgCRCK1 depletion on parasite replication and lytic cycle progression, we performed plaque assays on confluent human foreskin fibroblast (HFF) monolayers (Fig. 3A). Since the downregulation of TgCRCK1 involves culturing parasites in the presence of auxin, we first wanted to test if auxin treatment has any adverse effect on Toxoplasma growth. Towards this goal, we performed plaque assays for the parental strain (RHΔKu80-TIR1) in the presence or absence of auxin. The results showed that there was no significant difference in the number and size of the plaques formed by parental strain in the presence or absence of auxin (Fig. 3A through C). These findings suggested that auxin treatment does not affect Toxoplasma propagation in vitro. Next, to determine the contribution of TgCRCK1 to parasite growth, we performed plaque assays using the TgCRCK1.mAID.HA strain in the presence or absence of auxin. Importantly, we observed a significant reduction in the number (~18%) as well as the size (~55%) of the plaques with parasites lacking TgCRCK1 compared with the untreated parasites (Fig. 3D, E and F). These findings suggest that TgCRCK1 protein is indeed critical for efficient Toxoplasma growth in vitro.
*TgCRCK1 is critical for parasite growth in vitro. (A) Plaque assays were performed to examine the growth of the parental strain (RHΔku80-TIR1) in the presence or absence of auxin. Plaques are visible as clear zones on the background of crystal violet-stained HFF monolayer. Quantification of plaque numbers (B) and plaque size (C) for parental strain grown in the presence and absence of auxin. Data are presented as mean ± standard deviations from three independent experiments. ns, not significant. Unpaired two-tailed t-test. (D) Plaque assay showing the growth of TgCRCK1.mAID.HA strain cultured in presence or absence of auxin. Plaques are visible as clear zones on the background of a crystal violet-stained HFF monolayer. Quantification of plaque numbers (E) and plaque size (F) for TgCRCK1.mAID.HA parasites grown in the presence and absence of auxin. Data are presented as mean ± standard deviations from three independent experiments. *, P < 0.05 unpaired two-tailed t-test. *, P < 0.01 unpaired two tailed t-test.
TgCRCK1-deficient parasites show a defect in host-cell invasion and parasite division
An impairment of plaque formation can be caused by defects in one or more steps of the parasite lytic cycle, including host-cell invasion, egress, or cell division. Therefore, we next sought to determine which aspect of the lytic cycle was impaired in parasites lacking TgCRCK1. We first performed egress assays using calcium ionophore A23187. However, we did not see any significant difference in egress capability of parasites treated either with vehicle control or auxin, thus indicating that TgCRCK1 does not appear to play any role in this event (Fig. 4A). We next assessed parasite replication using standard doubling assays (23), and we observed that 24 h post-infection, auxin treatment resulted in fewer numbers of parasites per vacuole compared with wild-type parasites (Fig. 4B). These results suggest that TgCRCK1 is required for efficient intracellular growth of Toxoplasma. Further, we also performed parasite invasion assay into host cells, and we found there is moderate reduction (~33%) in the number of invaded parasites with auxin-treated parasites compared with the control (Fig. 4C). These results suggest that TgCRCK1 is important for the Toxoplasma host-cell invasion process.
*TgCRCK1-deficient parasites exhibit defects in host cell invasion and endodyogeny. TgCRCK1 parasites were subjected to lytic cycle assays, including induced egress (A), replication (B), and host-cell invasion (C) assays in the presence or absence of auxin. Data represent the mean and standard deviation results from three independent experiments, each performed with technical triplicates. ns, not significant. *, P < 0.05 Unpaired two-tailed t test. (D) Immunofluorescence examination of intracellular TgCRCK1.mAID.HA parasites treated with vehicle control (top two panels), or auxin (bottom two panels) followed by staining with anti-HA antibody and IMC3. scale bar, 2 μm. (E) Quantification of abnormal vacuoles in intracellular parasites treated with either vehicle control or auxin. Data are presented as mean ± standard deviations from three independent experiments. , P < 0.05, Unpaired two-tailed t-test.
We also examined cell division in parasites treated with auxin or the vehicle control. We noticed that untreated parasites were able to undergo endodyogeny leading to the formation of normal daughter cells as expected (Fig. 4D). However, in parasites treated with auxin, we observed a significant increase in the number of vacuoles that showed abnormal features (Fig. 4D and E). This included multiple buddings in a single mother parasite as well as an odd number of parasites per vacuole (Fig. 4D and E). Additionally, TgCRCK1-deficient parasites showed defects in centrosome division, characterized by discrepancies between the number of centrosomes and parasite nuclei (Fig. S1A). We also observed abnormal apical cap formation in daughter cells lacking TgCRCK1, which may contribute to the irregular parasite numbers per vacuole (Fig. S1B). Collectively, these findings indicate that TgCRCK1 plays a critical role in Toxoplasma endodyogeny.
Depletion of TgCRCK1 alters global gene expression in T. gondii
To determine whether loss of TgCRCK1 affects gene expression in Toxoplasma, RNA sequencing (RNA-seq) analysis was performed on intracellular parasites. We purified RNA from TgCRCK1.mAID-HA parasites treated with vehicle control or auxin and performed transcriptomic analysis to assess the effects of loss of TgCRCK1 on gene expression globally. The RNA sequencing studies revealed that there were a total of 93 genes that were differentially expressed in parasites lacking TgCRCK1. Of these, 58 genes were downregulated, while 35 genes were upregulated (Fig. 5A and Table S2). To gain functional insights, we manually categorized the differentially expressed genes according to their known or predicted functions and also their cell cycle expression profile (43, 44). Among the regulated genes, a large percentage were related to parasite metabolism, while the upregulated gene set contained a major percentage of genes involved in gene expression (Fig. 5B and C and Table S2). Within the downregulated data set, 32 genes are constitutively expressed, 13 genes show cell cycle regulation, and there are no data available for the remaining 13 genes. Among the upregulated genes, 24 genes show constitutive expression, 10 genes exhibit temporal expression profile, and no data are available for one gene (Table S2). Together, these findings indicate that loss of TgCRCK1 does cause transcriptional changes in the parasite.
Loss of TGGT1_275610 results in dysregulation of gene expression in Toxoplasma. (A) Volcano plot illustrating the statistical significance (−log10 P-value) versus fold change (log2) in the gene expression of intracellular parasites treated with vehicle control or auxin. Differentially expressed genes (log2 fold change ≥ 2) are shown, with downregulated genes (n = 58) indicated in red on the left and upregulated genes (n = 35) in red on the right. (B) and (C) Pie charts showing the functional classification of downregulated (B) and upregulated (C) genes. Gene classifications were based on known or putative functions, derived from conserved domains.
Loss of TgCRCK1 does not attenuate the parasite’s virulence in vivo
Given the importance of TgCRCK1 for parasite fitness in vitro, we next sought to determine its role in Toxoplasma pathogenesis in vivo. To assess virulence, female BALB/c mice were inoculated either with 20 or 100 tachyzoites of TgCRCK1.mAID-HA strain per mouse via intra-peritoneal route (n = 5 mice/group). Auxin was administered orally and also through daily intraperitoneal injections to maintain continuous protein depletion in the knockdown groups (45). Strikingly, with both doses of parasite inoculum (including those treated with auxin or vehicle control), we observed animals succumbing to parasite infection in the first 8 days (Fig. 6A and B). These findings suggest that loss of TgCRCK1 does not attenuate Toxoplasma virulence in this animal model.
Loss of TgCRCK1 does not attenuate parasite virulence in vivo. Survival curves for female mice injected intraperitoneally with either (A) 20 or (B) 100 tachyzoites of TgCRCK1.mAID.HA strain. Five female BALB/c mice were included in each group. The animals were subsequently treated with either vehicle control or auxin orally and also intraperitoneally. ns, not significant. Log-rank (Mantel-Cox) test (GraphPad Prism).
DISCUSSION
T. gondii relies on a complex array of regulatory mechanisms to invade host cells, replicate, and propagate within its host (8, 9, 46, 47), and protein kinases play central roles in many of these events (16). In this study, we characterized TgCRCK1, a temporally regulated kinase, and demonstrated its importance in the in vitro lytic cycle, linking its contribution to cell division, host cell invasion, and gene regulation.
We found that TgCRCK1 localizes to the parasite cytoplasm and is exclusively expressed in the S and M/C phases of the cell cycle. Our findings that TgCRCK1 is a developmentally regulated cytoplasmic kinase are indeed in agreement with a previous study (20). Thus far, the precise mechanism involved in tight regulation of expression of cell cycle-regulated proteins in Toxoplasma has not been defined. However, since the protein expression pattern closely follows the transcriptomic expression profile (39), it is quite likely that the TgCRCK1 appearance in the parasite is controlled at the transcriptional level. Hence, investigating the promoter-proximal elements responsible for this regulation would be an interesting avenue for future research.
The genome-wide screen indicated that TgCRCK1 may be essential for Toxoplasma viability, as evidenced by a phenotype score of −4.25 (40). However, our plaque assays demonstrated that although TgCRCK1 plays a significant role in parasite growth, it is not indispensable for Toxoplasma fitness. This observation is not uncommon, as many proteins with negative phenotypic scores have been demonstrated to be non-essential for parasite propagation (43, 48, 49).
The timing of expression of TgCRCK1 is confined to the S and M/C phases during Toxoplasma cell cycle. This temporal regulation of expression of this kinase provided an initial inkling that TgCRCK1 could be playing a role in endodyogeny. Additionally, a small pool of this protein was found to be associated with the pellicle region of the newly forming daughter parasites. Accordingly, we did see that the loss of TgCRCK1 is associated with defects in endodyogeny that included vacuoles containing odd numbers of parasites as well as abnormal daughter cell buddings. To assess whether TgCRCK1 plays a role in egress, we examined the egress efficiency of TgCRCK1-depleted parasites using the calcium ionophore A23187. Although we did not observe a significant reduction in ionophore-induced egress, it remains possible that TgCRCK1 contributes to other egress pathways (50). Future studies using alternative egress stimuli or monitoring natural egress dynamics may help clarify whether TgCRCK1 is involved in this process.
In addition to defects in replication, we noticed some moderate defect in host-cell invasion capability of parasites lacking TgCRCK1. This invasion defect is likely an indirect consequence of TgCRCK1 deficiency, as the protein is not expressed during the G1 phase. The invasion defect seen during TgCRCK1 deficiency could be due to multiple reasons. Firstly, studies have shown that although a large percentage of the extracellular parasite population is in G1 phase of the cell cycle, there is a significant proportion of parasites in the S and M/C phases, as well (51). Hence, it is feasible that in those extracellular parasites in the S and M/C phases, lack of TgCRCK1 could be affecting parasite fitness leading to reduced invasion capacity. Second, it is also feasible that defects arising during endodyogeny due to absence of TgCRCK1 could have downstream effects that impair parasite invasion fitness.
Loss of TgCRCK1 also leads to dysregulation of gene expression in the parasite, although the number of affected genes and the magnitude of fold changes are relatively modest. Given that TgCRCK1 is a kinase, its primary role in parasite biology is likely mediated through phosphorylation of substrate proteins rather than direct transcriptional regulation. Therefore, the relatively subtle changes observed at the transcriptome level are not unexpected. To better understand the impact of TgCRCK1 on phosphorylation events, quantitative phosphoproteomics experiments will be essential (52). Combining phosphoproteomics data with techniques such as proximity labeling (53, 54) could help identify TgCRCK1 substrates and clarify the precise mechanisms by which this kinase contributes to Toxoplasma propagation.
Despite the modest transcriptomic effects, several downregulated genes are of particular interest. Notably, one of the most significantly downregulated genes encodes a kelch repeat-containing protein (TGGT1_229290). In the related apicomplexan Plasmodium, kelch domain proteins have been implicated in cytoskeletal organization, motility, and host cell invasion (55). Another downregulated gene, TGGT1_209970, encodes a member of the Spc97/Spc98 family, which is known to play a critical role in microtubule organization (56). Additionally, several genes involved in gene expression (TGGT1_254140, TGGT1_262750, TGGT1_203170, and TGGT1_258990) and metabolism (TGGT1_255200, TGGT1_231000, TGGT1_219630, and TGGT1_257480) are also downregulated. Many of the affected genes are annotated as hypothetical proteins, including TGGT1_220060A, TGGT1_225890, and TGGT1_254530, some of which are predicted to be essential for Toxoplasma fitness (40). Given the functional importance of these genes, it is not surprising that TgCRCK1 deficiency leads to a marked reduction in parasite propagation.
Although TgCRCK1 is critical for Toxoplasma growth in vitro, its loss did not affect parasite virulence in a murine infection model. This discrepancy could reflect incomplete TgCRCK1 knockdown in vivo following auxin treatment. However, because our findings are consistent with a CRISPR-based genome-wide screen that identified genes involved in acute toxoplasmosis in mice (57), this explanation is unlikely. A more plausible interpretation is that functional redundancy among parasite kinases may compensate for the absence of TgCRCK1. Alternatively, TgCRCK1 may primarily function during lytic cycle events, with limited involvement in host immune modulation. Nonetheless, these results do not exclude a potential role for TgCRCK1 in parasite pathogenesis. Notably, TgCRCK1 is also expressed in bradyzoites (58), suggesting it may contribute to chronic stages of infection. Future studies investigating its function in bradyzoite biology may uncover additional roles for this kinase in the Toxoplasma life cycle.
In summary, we have identified a cell cycle-regulated kinase, TgCRCK1, which is essential for Toxoplasma growth in vitro. Future studies aimed at elucidating the TgCRCK1 interactome and characterizing its candidate substrate proteins will provide deeper insights into the kinase’s mechanism of action and the signaling pathways it regulates.
MATERIALS AND METHODS
Host cells and parasite culture
T. gondii tachyzoites were maintained through continuous passaging in HFF cells within a humidified incubator set at 37°C with 5% CO_2_. The culture medium used was Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% fetal bovine serum (FBS), L-glutamine (2 mM), and penicillin-streptomycin (50 µg/mL). Standard procedures were followed for parasite maintenance, harvesting, and purification (43, 44, 59).
Endogenous tagging of TgCRCK1 and generation of TgCRCK1.mAID.HA strain
The endogenous tagging of TgCRCK1 at the C-terminus was performed according to previously published protocols (42, 60, 61). Briefly, a single guide RNA (sgRNA) plasmid containing a protospacer against the 3′ untranslated region (UTR) of TgCRCK1 downstream of the stop codon was generated through site-directed mutagenesis. The homology-directed repair (HDR) templates were PCR amplified using the vectors, p3XHA.LIC-DHFR and pmAID3xHA.LIC-HPT that contain HA and mAID.HA epitope tags, respectively. Additionally, both the plasmids contain a selection cassette that provides resistance against pyrimethamine (DHFR) and MPA-Xanthine (HXGPRT) (62, 63). The 60-bp primers used in generating the repair template include 40 bp of homology immediately upstream of the stop codon or 40 bp of homology within the 3′ UTR downstream of the CRISPR/Cas9 cut site. All primers that were used for pU6-Universal plasmids and HDR templates are listed in Table S1. The sgRNA plasmid and the HDR templates were then transfected into RHΔKu80 or RHΔKu80.TIR1 strain using nucleofector (35). Transfected parasites were then cultured in the presence of either pyrimethamine or MPA/xanthine to select stably transformed parasites that were cloned by limiting dilution. The clones were screened and validated by PCR and sequencing.
Immunofluorescence microscopy
Immunofluorescence staining of the intracellular parasites was carried out as described previously (35, 64, 65). Primary antibodies used were mouse anti-HA (Cell Signaling Technology, Inc., 6E2, 1:250), rabbit anti-IMC3 (1:500) (35, 43), TgCentrin1 (1:250) (66) and TgISP1 (1:500) (67). Secondary antibodies were Alexa Fluor 594-conjugated goat anti-rabbit or Alexa Fluor 488-conjugated goat anti-mouse (35, 43) (Molecular Probes, 1:1,000). Imaging was performed using a Zeiss Axio Observer seven microscope (Carl Zeiss). Digital images were acquired utilizing Axiocam 506 mono charge-coupled device (CCD) camera using Axiovision software.
Plaque assays
Plaque assays were performed as described previously with some modifications (44, 68). Intracellular parasites were harvested, syringe filtered, and added onto a confluent monolayer of HFF cells in a 12-well plate containing medium (DMEM with 10% FBS) with vehicle control or auxin (500 μM IAA). The plates were then incubated at 37°C for 6 days without any movement. The plates were then washed with PBS, methanol fixed, and stained with 2% crystal violet to visualize regions of host cell disruption. Plaques were imaged using the Molecular Imager Gel Doc XR system (Bio-Rad) and analyzed using Image Lab software. ImageJ was used to outline and quantify plaque areas. Experiments were conducted in triplicate to determine average plaque number and sizes.
Replication assay
To assess the parasite doubling time, freshly egressed parasites were inoculated into confluent HFF monolayers in 12-well plates and allowed to invade for 2 h. The monolayers were then washed three times with medium (DMEM with 10% FBS) to remove uninvaded parasites and incubated at 37°C in DMEM with 10% FBS containing vehicle control or presence of 500 μM IAA. At 24 h post-infection, the cells were fixed with methanol and stained using Diff-Quik (Dade-Behring) according to the manufacturer’s instructions. For each treatment, at least 100 vacuoles from three biological replicates were assessed for the number of parasites per vacuole.
Ionophore-induced egress assay
Egress efficiency following calcium ionophore treatment was assessed following established protocols (44, 69, 70). Freshly harvested parasites were added to confluent HFF monolayers in 24-well plates at a multiplicity of infection (MOI) of 1 and then incubated at 37°C for 28 h. The cultures were then treated with vehicle control or auxin for 4 h, and egress was induced using calcium ionophore A23187 (1 µM) in Hanks’ Balanced Salt Solution (HBSS) at 37°C for 2 minutes. Cultures were then fixed with methanol and stained using Diff-Quik (Dade-Behring). Egress percentage was calculated by dividing the number of lysed vacuoles by the total vacuole count per sample.
Invasion assays
Invasion assays were performed in eight-well chamber slides as described previously with the following modifications (71, 72). Briefly, parasites were pretreated with vehicle control or 500 μM IAA (to deplete the mAID-3HA tagged TgCRCK1) in DMEM with 10%FBS for 4 h. Purified tachyzoites were then added onto HFF monolayers (2 × 10^6^ parasites well) and incubated at 37°C for 30 min in the presence or absence of auxin to allow invasion. Slides were then washed three times to remove non-invaded parasites, fixed, blocked, and stained with mouse anti-SAG1 without permeabilization. After 1 h, slides were washed, permeabilized with 0.01% TX-100 and stained with rabbit anti-M2AP antibody. The slides were further washed and stained with secondary antibodies, Alexa Fluor-594-conjugated goat anti-mouse (Molecular Probes) and Alexa Fluor-488-conjugated goat anti-rabbit (Molecular Probes). After 1 h, slides were washed and mounted using Vectashield (with DAPI). Parasites that were both red and green were identified as extracellular (attached), whereas those that were green but not red were identified as intracellular (invaded). Images of 10 random fields of view within each well were captured at 600× magnification, and the total number of intracellular parasites and host cell nuclei were enumerated.
RNA sequencing and differential gene expression analysis
RNA sequencing was performed according to previously published protocols with some modifications (35, 73). Total RNA from intracellular TgCRCK1.mAID.HA parasites treated with 500 μM IAA or vehicle control for 4 h was isolated using the RNeasy kit (Qiagen). RNA samples were obtained from three independent experiments. The quality of total RNA samples was verified by using a BioAnalyzer (Agilent), followed by digestion with DNAse I (NEB). Ribosomal RNA was removed using the Ribo-Zero rRNA removal kit (human/mouse/rat, Illumina). Sequencing libraries were then generated using the TruSeq RNA Sample Prep Kit (v2, Illumina) according to manufacturer’s protocol. Libraries were amplified using the TruSeq Cluster Kit (v3, Illumina), and subjected to 50-bp single-end sequencing with the Illumina HiSeq 2000 system. Sequencing reads were aligned to the Toxoplasma GT1 reference genome (ToxoDB v.53, https://toxodb.org/toxo/app) using the STAR software package (v.2.7.1a , with default settings) (74). Filtered and normalized gene expression levels were calculated from the aligned reads using HTSeq v.0.13.5 (75). Differentially expressed genes were identified by linear modeling and Bayesian statistics using the limma package for R v.3.49.1, (76).
In vivo virulence assay
The virulence assays were performed according to previously published studies (77). All laboratory animal work in this study was conducted in compliance with guidelines from the Virginia Tech Committee on the Use and Care of Animals (IACUC) (protocol no. 23-163). Female 6-week-old BALB/c mice (Jackson Laboratories) were intraperitoneally injected with TgCRCK1.mAID-HA parasites (20 or 100 tachyzoites per mouse). For the knockdown groups, auxin was provided in the drinking water (500 mg/L) and given intraperitoneally (150 mg/kg) every day throughout the experiment (77). Parasite viability was confirmed immediately post-infection through plaque assays from the same preparation used for mouse injections. Mice were monitored throughout the experiment for disease symptoms, and survival was recorded.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Weiss LM, Dubey JP. 2009. Toxoplasmosis: a history of clinical observations. Int J Parasitol 39:895–901. doi:10.1016/j.ijpara.2009.02.00419217908 PMC 2704023 · doi ↗ · pubmed ↗
- 2Halonen SK, Weiss LM. 2013. Toxoplasmosis. Handb Clin Neurol 114:125–145. doi:10.1016/B 978-0-444-53490-3.00008-X 23829904 PMC 4157368 · doi ↗ · pubmed ↗
- 3Dubey JP. 2002. A review of toxoplasmosis in wild birds. Vet Parasitol (Amst) 106:121–153. doi:10.1016/S 0304-4017(02)00034-1 · doi ↗
- 4Dubey JP. 1998. Advances in the life cycle of Toxoplasma gondii. Int J Parasitol 28:1019–1024. doi:10.1016/s 0020-7519(98)00023-x 9724872 · doi ↗ · pubmed ↗
- 5Smith JL. 1997. Long-term consequences of foodborne toxoplasmosis: effects on the unborn, the immunocompromised, the elderly, and the immunocompetent. J Food Prot 60:1595–1611. doi:10.4315/0362-028X-60.12.159531207758 · doi ↗ · pubmed ↗
- 6Luft BJ, Remington JS. 1992. Toxoplasmic encephalitis in AIDS. Clin Infect Dis 15:211–222. doi:10.1093/clinids/15.2.2111520757 · doi ↗ · pubmed ↗
- 7Jones JL, Lopez A, Wilson M, Schulkin J, Gibbs R. 2001. Congenital toxoplasmosis: a review. Obstet Gynecol Surv 56:296–305. doi:10.1097/00006254-200105000-0002511333376 · doi ↗ · pubmed ↗
- 8Blader IJ, Coleman BI, Chen C-T, Gubbels M-J. 2015. Lytic Cycle of Toxoplasma gondii: 15 years later. Annu Rev Microbiol 69:463–485. doi:10.1146/annurev-micro-091014-10410026332089 PMC 4659696 · doi ↗ · pubmed ↗