Single-strain and consortium inoculations with plant-beneficial Pseudomonas spp. promote lettuce growth under field conditions
Adrien Biessy, Mélanie Cadieux, Florence Mc Duff, Arianne Deshaies, Kosal Khun, Joël Lafond-Lapalme, Philippe Vigneault, Martin Filion

TL;DR
Using two beneficial Pseudomonas bacteria together helps lettuce grow better in the field compared to using just one strain.
Contribution
The study shows that a Pseudomonas consortium improves lettuce growth under field conditions and reveals a beneficial interaction between the strains.
Findings
Pseudomonas consortium and P. protegens B21-024 significantly increased lettuce shoot dry weight in the field.
Co-inoculation increased P. protegens B21-024 population in the rhizosphere compared to single-strain inoculation.
Nitrogen-rich medium used for inoculum preparation contributed to plant growth promotion.
Abstract
Plant-beneficial Pseudomonas spp. can promote plant growth by a wealth of mechanisms. Developing microbial inoculants encompassing multiple Pseudomonas strains (consortia) could represent an attractive solution to some of the problems encountered by single-strain inoculants, such as inconsistent rhizosphere colonization under field conditions. In this study, we evaluated the potential of Pseudomonas protegens B21-024 and Pseudomonas putida B21-029, inoculated alone or in combination, to promote lettuce growth under greenhouse and representative commercial field conditions. Plant growth metrics, including shoot fresh and dry weights, were used to evaluate plant growth promotion. In addition, unmanned aerial vehicle imaging was used to monitor plant growth in the field. Strain-specific primer-probe sets were also developed to study, using the quantitative polymerase chain reaction, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
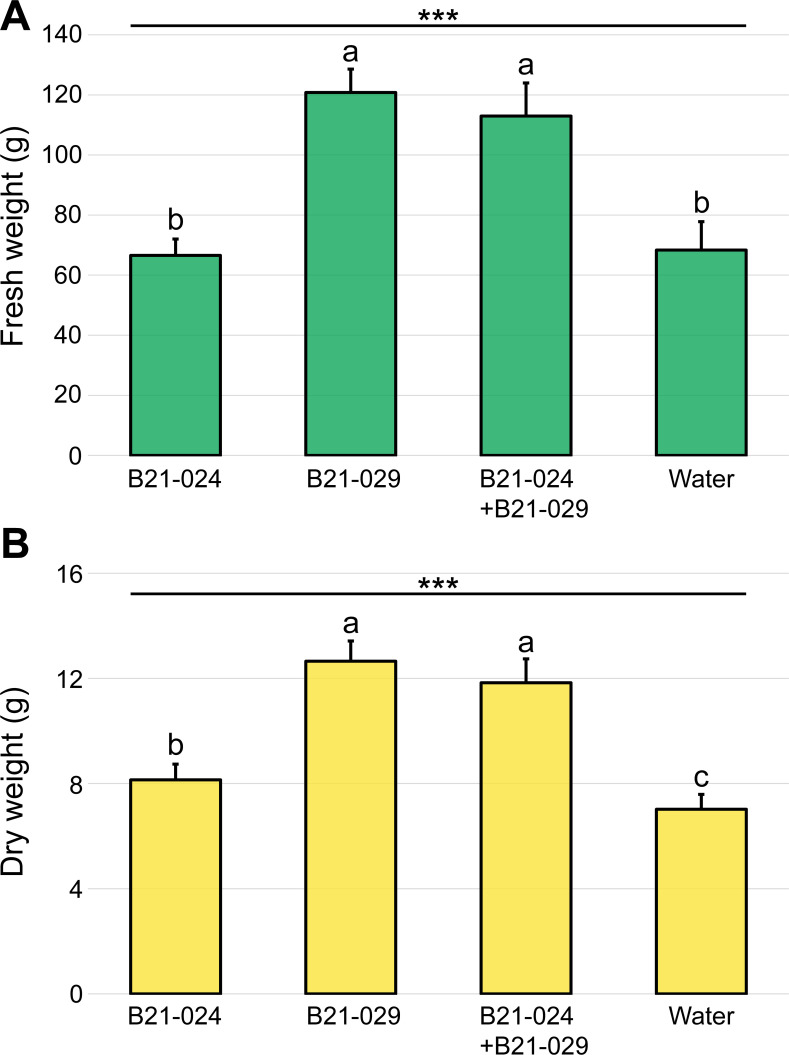 Fig 1
Fig 1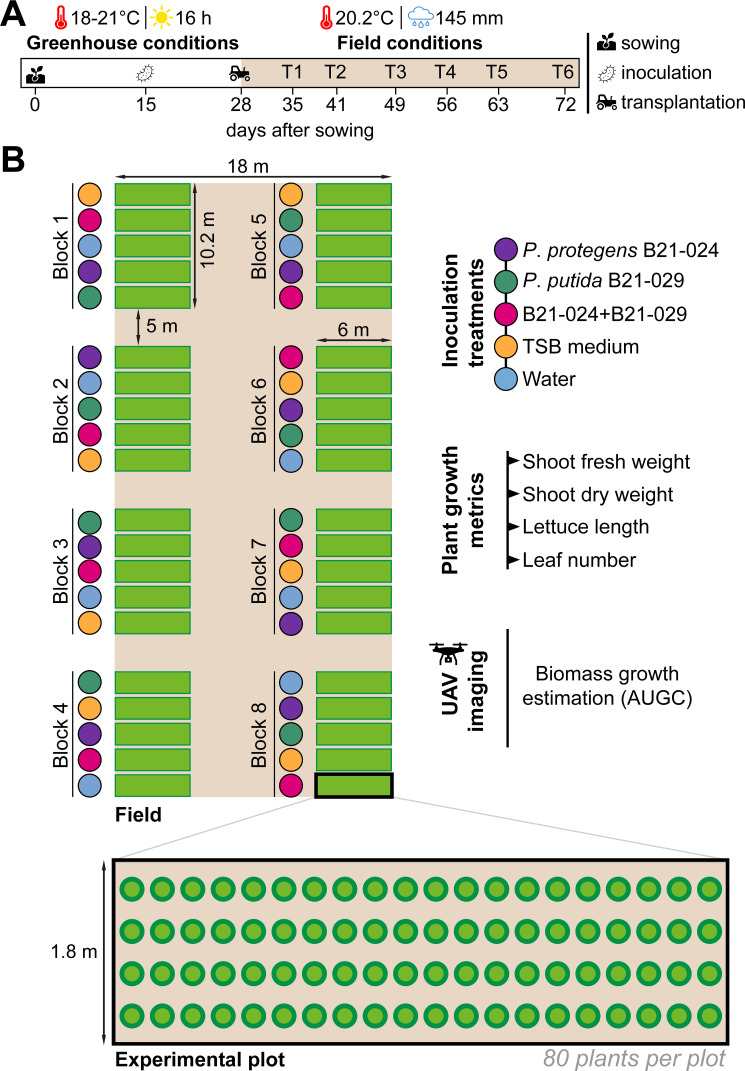 Fig 2
Fig 2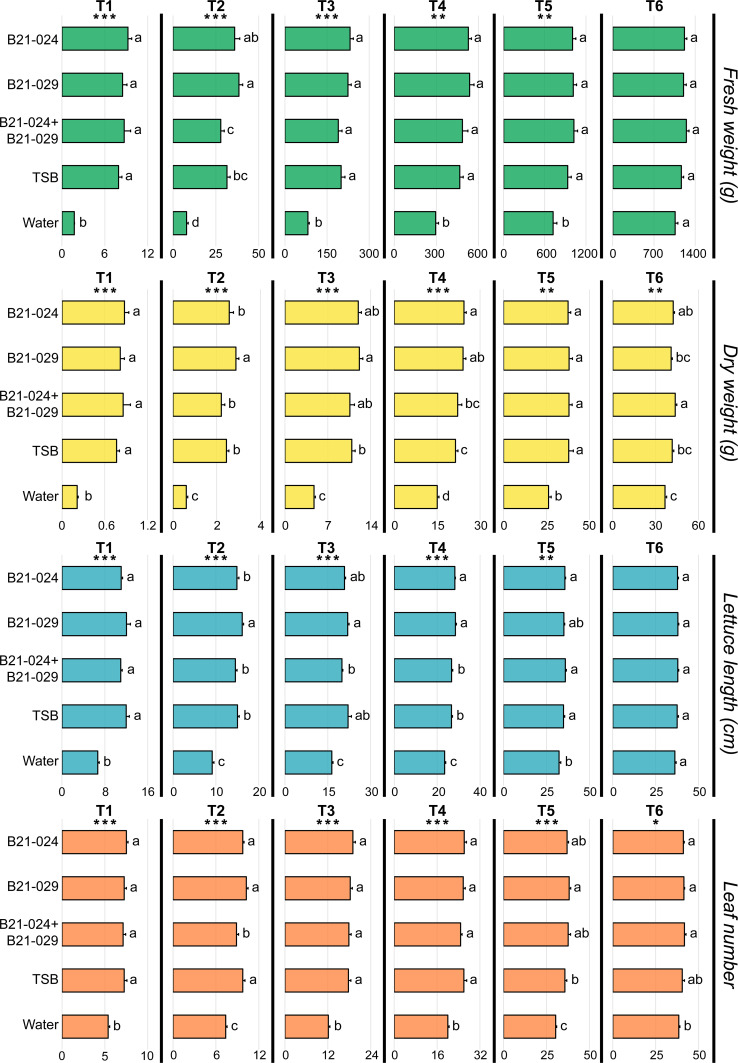 Fig 3
Fig 3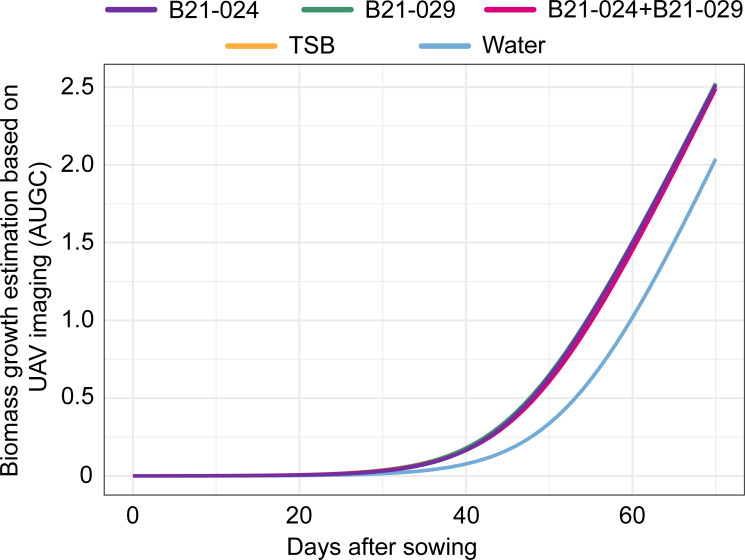 Fig 4
Fig 4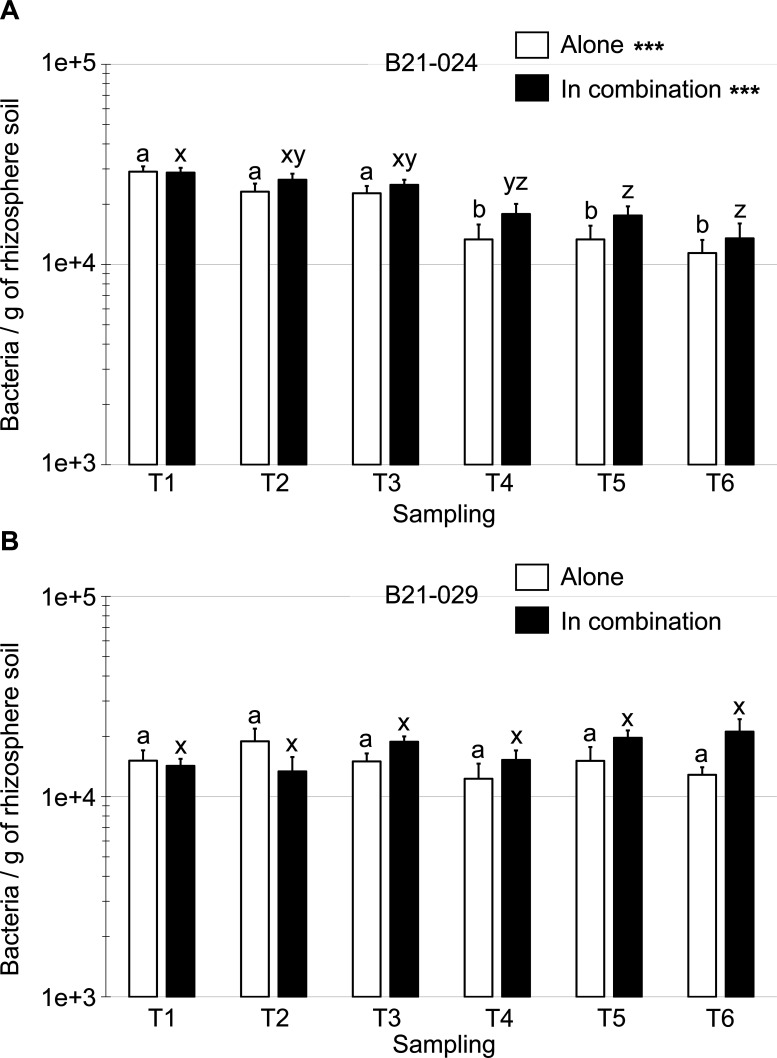 Fig 5
Fig 5| Strain | Target gene | Primer/probe name | Nucleotide sequence (5′ → 3′) | Amplicon size (bp) | Efficiency (%) |
|---|---|---|---|---|---|
| LOY29_14745 | B21-024_F |
| 56 | 104.5 | |
| B21-024_R |
| ||||
| B21-024_P |
| ||||
| LOY24_13695 | B21-029_F |
| 61 | 103.9 | |
| B21-029_R |
| ||||
| B21-029_P |
|
- —Agriculture and Agri-Food Canadahttp://dx.doi.org/10.13039/501100000040
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Pathogenic Bacteria Studies · Bacterial biofilms and quorum sensing
INTRODUCTION
Members of the genus Pseudomonas are rod-shaped Gram-negative Gammaproteobacteria that are found in many environments, including soil, water, and higher organisms (1). This large and diverse genus includes more than 300 species, some of which can cause diseases to plants and animals (2). For example, P. syringae is one of the most devastating bacterial plant pathogens to roam the phyllosphere (3, 4). By contrast, many Pseudomonas species and strains establish mutually beneficial relationships with a myriad of plant species, including many important crops (5–7). Plant-beneficial Pseudomonas spp. aggressively colonize the rhizosphere—the zone under the influence of the roots—and use root exudates as source of carbon and energy (5, 8, 9). In return, these microorganisms often protect the plant root system against soilborne plant pathogens (5, 7, 10), help the plant to cope with abiotic stresses (11), and/or directly promote plant growth (12).
Direct plant growth promotion by plant-beneficial Pseudomonas spp. is mediated by a wealth of mechanisms that result in biomass and/or yield increases. One of the main mechanisms used by plant-beneficial Pseudomonas spp. is the modulation of the plant hormonal balance (13). For example, many Pseudomonas strains produce phytohormones, such as auxins and gibberellins (14–16). Production of the auxin molecule indole-3-acetic acid (IAA) by plant-beneficial Pseudomonas spp. has been shown to increase root biomass and to promote the development of the root system, either by stimulating primary root elongation or by increasing root hair formation (13, 15). Conversely, gibberellic acid production increases shoot length and shoot fresh/dry weight (14). Moreover, many Pseudomonas strains produce the 1-aminocyclopropane-1-carboxylate (ACC) deaminase enzyme that degrades ACC, the precursor of the stress hormone ethylene (17–19). This enables the bacteria to act as a sink for ACC and to modulate endogenous ethylene levels, promoting root elongation and increasing biomass (17). Additional plant growth promotion mechanisms include phosphate solubilization (20, 21), and the production of volatile organic compounds with plant growth promotion activity (22, 23). By promoting the development of the root system, plant-beneficial Pseudomonas spp. improve the ability of the plant to absorb nutrients, thereby accelerating its growth and increasing crop yield.
Lettuce (Lactuca sativa L., Asteraceae) is an important horticultural crop that is grown in most areas worldwide. Growers are interested in new sustainable methods to increase yield and enhance lettuce nutritional quality. Many Pseudomonas strains have been shown to enhance lettuce growth under both controlled and field conditions (24–28). For example, Cipriano et al. reported that single-strain inoculations with Pseudomonas spp. promoted lettuce growth under field conditions (24). Over the years, the development of microbial inoculants encompassing multiple strains (microbial consortia) has been increasingly recognized as an attractive solution to some of the problems of single-strain inoculants, namely insufficient rhizosphere colonization and inconsistent plant growth promotion/disease suppression (10, 29–31). However, Pseudomonas spp. are known for their ability to produce a wide diversity of secondary metabolites with antimicrobial activity (32), which complexifies their successful association with other bacteria. For example, Pseudomonas strains producing the antibiotic 2,4-diacetylphloroglucinol (DAPG) inhibit the growth of bacteria from two major plant-beneficial genera, Bacillus and Azospirillum (28, 33, 34). Combining two or more Pseudomonas strains could potentially reduce the risk of incompatibility between bacterial strains during co-inoculation.
In a previous study, we showed that inoculation with the plant-beneficial strain Pseudomonas protegens B21-024 promoted lettuce growth under field conditions (27). This strain significantly increased lettuce shoot fresh/dry weight under optimal nitrogen conditions. In addition, inoculation with P. protegens B21-024 compensated for most of the yield losses caused by reduced nitrogen fertilizer application. Based on these promising results, co-inoculating P. protegens B21-024 with other Pseudomonas strains appears as an attractive strategy that could further improve lettuce growth.
In this study, we evaluated the ability of two Pseudomonas strains, P. protegens B21-024 and Pseudomonas putida B21-029, to promote lettuce growth and yield. First, we validated the plant growth promotion ability of the two strains, alone and in combination, under greenhouse conditions. Second, we carried out a field experiment to validate the plant growth promotion activity of the two strains, alone and in combination, under representative commercial field conditions. We used a combination of weekly manual plant sampling and biweekly unmanned aerial vehicle (UAV) flights to monitor lettuce growth. Finally, we designed two strain-specific primer-probe sets to quantify, using quantitative polymerase chain reaction (qPCR), the abundance of the two bacterial strains in the rhizosphere soil surrounding lettuce plants throughout the growing season.
RESULTS
Lettuce growth promotion under greenhouse conditions
As a first step to evaluating the plant growth promotion potential of P. protegens B21-024 and P. putida B21-029, alone and in combination, a greenhouse experiment was carried out over a 55-day period. The inoculation treatments included i) P. protegens B21-024, ii) P. putida B21-029, iii) the two Pseudomonas strains in combination, and iv) a water control. P. putida B21-029 and the Pseudomonas consortium (B21−024+B21-29) significantly increased lettuce aboveground fresh biomass by 77% and 65%, respectively, when compared to the water-inoculated lettuce plants (Fig. 1). In addition, both treatments significantly increased shoot dry weight, with increases ranging from 69% to 80%. While inoculation with P. protegens B21-024 alone did not significantly increase aboveground fresh biomass (Fig. 1), this treatment significantly increased shoot dry weight by 16%.
Plant growth promotion under greenhouse conditions. Lettuce seedlings were inoculated 13 days after sowing with four different treatments: three bacterial treatments (P. protegens B21-024, P. putida B21-029, and both Pseudomonas strains in combination) and a water control. Fifty-five days after sowing, the plants were harvested, and the shoot fresh weight (A) and shoot dry weight (B) were measured. The values are presented as means + standard errors. The three asterisks indicate a P value inferior to 0.001 (Friedman test). Treatments with different letters are significantly different according to Fisher’s LSD post hoc test (α = 0.05).
Lettuce growth promotion under field conditions
A field experiment was carried out in the summer of 2024 to test the plant growth promotion effect of P. protegens B21-024 and P. putida B21-029, alone and in combination, under representative commercial field conditions (Fig. 2). Five treatments were used: three bacterial treatments (P. protegens B21-024, P. putida B21-029, and a consortium of these two strains), a tryptic soy broth (TSB) treatment, and a water control treatment. The TSB treatment was added as the two Pseudomonas strains were initially grown in TSB medium, and fresh TSB medium was added during the preparation of the inocula (one-third of the total volume) to support the establishment of the strains in the rhizosphere. In addition, this treatment previously promoted the growth of field-grown lettuce plants under limited nitrogen availability conditions (27). Therefore, the use of this treatment enabled us to monitor whether the TSB medium promotes plant growth and determine to what extent. Lettuce plants were harvested at six different sampling times during the growing season: 35 (T1), 41 (T2), 49 (T3), 56 (T4), 63 (T5), and 72 (T6) days after sowing. One randomly selected lettuce plant per plot was sampled during the five intermediate samplings (T1:T5; 8 per treatment), whereas five lettuce plants per plot were harvested at the final sampling (T6; 40 per treatment). Four plant growth metrics were measured: shoot fresh weight, shoot dry weight, lettuce length, and leaf number.
Timeline and experimental design of the field trial. (A) Romaine lettuce seeds were germinated and grown under greenhouse conditions until field transplantation. The seedlings were inoculated 15 days after sowing, and the plants were transplanted into the field 28 days after sowing. During the 44 days the lettuce plants were in the field, the average temperature was 20.2°C and total precipitation amounted to 145 mm. Lettuce plants were harvested at six different sampling times during the growing season: 35 (T1), 41 (T2), 49 (T3), 56 (T4), 63 (T5), and 72 (T6) days after sowing. (B) A randomized complete block design was used, with a total of eight blocks. The five inoculation treatments were randomized within each block, and each experimental plot was inoculated with the same treatment. The plots included a total of 80 lettuce plants each. One randomly selected lettuce plant per plot was sampled during the five intermediate samplings (T1, T2, T3, T4, and T5), whereas five lettuce plants per plot were harvested at the final sampling (T6). Four plant growth metrics were collected in this study: shoot fresh weight, shoot dry weight, lettuce length, leaf number. In addition, the growth of each individual lettuce was monitored throughout the growing season thanks to biweekly flights of an UAV mounted with imaging sensors. The area under the growth curve (AUGC) of the surface area metric was chosen to estimate aboveground biomass accumulation.
During most of the growing season (T1–T5), lettuce plants inoculated with one of the three bacterial treatments or with the TSB treatment had a higher aboveground biomass (fresh and dry weight) and a larger number of leaves than water-inoculated lettuce plants (Fig. 3). Most treatments also increased lettuce length when compared to the water control at the five intermediate samplings (T1–T5). The plant growth promotion effect of these four treatments was remarkably high at the first sampling times but gradually decreased as the growing season progressed (Fig. 3; Fig. S1). For example, at the first sampling time (35 days after sowing), the fresh biomass of lettuce plants inoculated with one of the three bacterial treatments or with the TSB treatment was, on average, more than four times greater than the fresh biomass of water-inoculated plants. Sixty-one days after sowing (T5), the differences between these four treatments and the water treatment ranged from 29% to 42%. While similar plant growth promotion was achieved by the three bacterial treatments and the TSB treatment (Fig. 3; Fig. S1), there were some differences between these four treatments at some of the five intermediate samplings. For example, 41 days after sowing (T2), lettuce plants inoculated with P. putida B21-024 had a higher dry weight and a higher lettuce length than the four other treatments. By contrast, 41 days after sowing (T2), lettuce plants inoculated with the Pseudomonas consortium treatment had, on average, fewer leaves than lettuce plants inoculated with the TSB treatment or with the other two bacterial treatments.
Plant growth promotion under field conditions. Lettuce plants were grown from seeds in a greenhouse and inoculated with five different treatments: P. protegens B21-024, P. putida B21-029, a combination of both Pseudomonas strains, TSB medium, and water. The plants were transplanted into the field 28 days after sowing. Lettuce plants were harvested at six different sampling times throughout the growing season: 35 (T1), 41 (T2), 49 (T3), 56 (T4), 63 (T5), and 72 (T6) days after sowing. Four plant growth metrics were measured: shoot fresh weight, shoot dry weight, lettuce length, and leaf number. The values are presented as means + standard errors. The five treatments were compared to each other at each sampling time independently. One, two, or three asterisks refer to P values inferior to 0.05, 0.01, and 0.001, respectively (Friedman test). Treatments with different letters are significantly different from each other according to Fisher’s LSD post hoc test (α = 0.05).
At harvest (T6), no significant differences in fresh weight were observed between the five treatments (Fig. 3). Even though the differences were not significant, lettuce plants inoculated with the Pseudomonas consortium treatment displayed, on average, an 18% higher fresh weight (1,250 g) than the water-inoculated plants (1,059 g), and 7% higher fresh weight than TSB-inoculated plants (1,164 g). In addition, there were no significant differences between the five treatments regarding lettuce length. Nevertheless, lettuce plants inoculated with P. protegens B21-024 or with the Pseudomonas consortium displayed a significantly higher dry weight than the water-inoculated lettuce plants (Fig. 3). The Pseudomonas consortium treatment led to the highest dry weight increase (+20%). In addition, it was the only treatment significantly different from the TSB treatment for shoot dry weight. Lettuce plants inoculated with one of the three bacterial treatments also had a significantly higher number of leaves than the water-inoculated plants at harvest (Fig. 3). At harvest (T6), the TSB treatment was not significantly different from the water treatment for any of the four plant growth metrics monitored in this study (Fig. 3).
In addition to plant growth metrics obtained from weekly manual sampling, biweekly UAV flights mounted with imaging sensors allowed us to closely monitor the growth of each individual lettuce throughout the growing season. UAV imaging is increasingly used to monitor the growth of agricultural crops under field conditions (35) and has been previously used to evaluate the effect of plant growth-promoting rhizobacteria on plant growth (27, 36). Three morphological metrics were calculated for more than 600 lettuce plants per treatment: height, surface area, and volume. The area under the growth curve (AUGC) of the surface area metric was previously chosen as the best morphological metric to monitor biomass accumulation in field-grown lettuce plants (27). The same choice was made in the present study. The AUGC values calculated for each treatment strongly correlated with shoot fresh weight (r^2^ = 0.97) and shoot dry weight values (r^2^ = 0.95) throughout the growing season (Fig. S2). When comparing AUGC values and shoot fresh/dry weight values for individual lettuce plants at harvest, the correlation was smaller (Fig. S3). The AUGC curves clearly demonstrate the plant growth-promoting effect of the four treatments (B21-024, B21-029, B21-024+B21-029, and TSB) when compared to the water control (Fig. 4). At harvest, plants inoculated with one of the three bacterial treatments or with the TSB treatment had significantly higher AUGC values than water-inoculated plants (P value = 0.0017). However, the AUGC values did not allow for discriminating the three bacterial treatments and the TSB treatment from each other.
Aboveground biomass estimation based on UAV imaging. The area under the growth curve (AUGC) was obtained by calculating the area under the logistic growth curve modeled from the surface area morphological metric.
Rhizosphere soil population dynamics of P. protegens B21-024 and P. putida B21-029
To quantify the abundance of the two Pseudomonas strains in rhizosphere soil throughout the growing season (field experiment), two novel strain-specific primer-probe sets were designed. The specificity of the two primer-probe sets was first validated with the DNA obtained from 20 Pseudomonas strains belonging to the P. fluorescens and P. putida groups (Table S1). qPCR amplifications were only observed from the DNA samples corresponding to P. protegens B21-024 and P. putida B21-29. In addition, no specific amplification was detected in the rhizosphere soil samples collected from lettuce plants inoculated with either TSB or water only. With the specificity of the two primer-probe sets validated, we studied the seasonal dynamics of both strains in rhizosphere soil. In addition, we compared the population dynamics of the Pseudomonas strains inoculated alone and in combination.
First, we studied the rhizosphere population dynamics of both strains when inoculated alone (thus excluding for now the rhizosphere soil samples from lettuce plants inoculated with the Pseudomonas consortium treatment). P. protegens B21-024 was detected in the rhizosphere soil at a population density of 2.9 × 10^4^ bacteria per gram of dry soil at the first sampling time (T1). The population of P. protegens B21-024 slightly decreased throughout the growing season, reaching a population of 1.1 × 10^4^ bacteria per gram of dry soil at the end of the growing season (T6). There were significant differences between the sampling times, and the population size of this strain was significantly higher at the first three sampling times. By contrast, the population of P. putida B21-029 was extremely stable throughout the growing season, and there were no significant differences between the different sampling times (Fig. 5). The rhizosphere soil population of P. putida B21-029 was 1.5 × 10^4^ bacteria per gram of dry soil at the first sampling time (T1). At the end of the growing season (T6), the equivalent of 1.3 × 10^4^ bacteria per gram of dry soil was detected by qPCR.
Abundance of P. protegens B21-024 (A) and P. putida B21-029 (B) in the rhizosphere of lettuce plants throughout the growing season. Each Pseudomonas strain was either inoculated alone (white histogram bar) or in combination with the other strain (black histogram bar). Rhizosphere soil samples were collected at six different times during the growing season: 35 (T1), 41 (T2), 49 (T3), 56 (T4), 63 (T5), and 72 (T6) days after sowing. Soil DNA was extracted, and the target sequences were amplified by quantitative PCR using strain-specific primer-probe sets. As each target sequence is present only in one copy in the genome of the bacteria under study, the abundance of the bacteria can be expressed as bacteria per gram of dry rhizosphere soil. Values are presented as means + standard errors. Three asterisks signal P values inferior to 0.001 (one-way ANOVA). Treatments with the same letter are not significantly different from each other according to Tukey’s HSD post hoc test (α = 0.05).
We then compared the abundance of the two Pseudomonas strains in the rhizosphere of lettuce plants when inoculated alone or in combination with the other Pseudomonas strain (Fig. 5, Fig. S4). When P. protegens B21-024 was inoculated in combination with P. putida B21-029, its rhizosphere soil population followed a similar trajectory to when the strain was inoculated alone. Its population size slightly decreased from 2.9 × 10^4^ (T1) to 1.4 × 10^4^ (T6) bacteria per gram of dry soil (Fig. 5). While the dynamic was similar, the population size of P. protegens B21-024 when inoculated in combination with P. putida B21-029 was significantly higher (P value = 0.026) than when this strain was inoculated alone (Fig. S4). When P. putida B21-029 was inoculated in combination with P. protegens B21-024, its population size was similar to when inoculated alone, with a rhizosphere population size ranging from 1.7 × 10^4^ to 2.1 × 10^4^ bacteria per gram of dry soil. There was also no difference between the population sizes of P. putida B21-029, whether the strain was inoculated alone and in combination (Fig. S4).
DISCUSSION
In this study, we explored the potential of two plant-beneficial Pseudomonas strains, P. protegens B21-024 and P. putida B21-029, to enhance lettuce growth under representative commercial field conditions. The goal was to develop an effective Pseudomonas consortium that would efficiently colonize the rhizosphere and synergistically increase lettuce yield.
To favor the establishment of the two Pseudomonas strains in the lettuce rhizosphere, fresh TSB medium was added during the preparation of the bacterial inocula. In addition, the bacteria were grown in TSB medium, and while most nutrients were likely used by the bacteria to grow, it is possible that a significant proportion remained. Therefore, a TSB treatment was included in the field experiment to monitor the potential effect of this nitrogen-rich medium on plant growth. It was previously demonstrated that the same TSB treatment promoted the growth of lettuce plants grown under nitrogen-limited conditions but not when using the recommended nitrogen application dose (27). In this study, the TSB treatment promoted the growth of lettuce plants during most of the growing season (T1–T5). Notably, this treatment was significantly different from the water treatment during the first five samplings across all four plant growth metrics (Fig. 3). In addition, biomass estimation based on UAV imaging also identified a plant growth promotion effect associated with this treatment throughout the growing season (Fig. 4).
TSB medium is mainly composed of protein hydrolysates, namely pancreatic digest of casein and papain digest of soybean meal, which are a well-studied class of plant biostimulants (37, 38). Different protein hydrolysates were previously shown to promote the growth (39, 40) and reduce salinity stress in lettuce plants (41). In this study, it is likely that the plant growth promotion effect displayed by TSB originates from its high content in amino acids and peptides, which can be taken up by plant roots as a source of nitrogen (42, 43). Alternatively, the use of TSB might favor the recruitment of indigenous plant-beneficial rhizobacteria, as previously reported in another study (44). Finally, it is also possible that the L-tryptophan amino acids present in casein and soy proteins (45) could be used by indigenous rhizobacteria to produce IAA, as this amino acid is the precursor of IAA in most bacterial biosynthetic pathways (46).
Despite its plant growth-promoting effect, the use of TSB alone could not have accounted for the superior plant growth promotion associated with some bacterial inoculation treatments used in this study. Some bacterial treatments achieved superior plant growth promotion at most of the five intermediate sampling times when compared to the TSB treatment (and the water control) (Fig. 3). Moreover, the TSB treatment was not significantly different from the water control for any of the four plant growth metrics at harvest (T6), while the consortium treatment was significantly superior to the TSB treatment (and the water treatment), and resulted in a significant increase in shoot dry weight (+20%) compared to the water control (Fig. 3). In that regard, the Pseudomonas consortium treatment was superior to the P. protegens B21-024 inoculation treatment, which was also significantly superior to the water control at harvest (T6) but not significantly different than the TSB treatment. Even though the Pseudomonas consortium did not significantly increase lettuce fresh weight at harvest, dry weight is usually considered to be more accurate than fresh weight when evaluating plant growth promotion by plant-beneficial rhizobacteria, mainly because variations in tissue water content between treatments can cause significant data distortion (47).
Single-strain inoculation with P. protegens B21-024 previously promoted the growth of lettuce plants under varying nitrogen fertilizer conditions, with dry weight increases ranging from 29.7% to 39.8% depending on the nitrogen fertilization regime (27). In the present study, lettuce plant inoculation with P. protegens B21-024 significantly increased shoot dry weight at harvest (T6) by 16% when compared with water-inoculated lettuce plants. However, this treatment was not significantly different from the TSB treatment for shoot dry weight. Several factors could explain this lower performance. First and foremost, the lettuce cultivars used were different, and the one used in the present study “Sun Valley” might be less susceptible to plant growth promotion by rhizobacteria. Several studies have identified a cultivar-specific variation in plant growth promotion by rhizobacteria (48, 49). Environmental conditions between the two growing seasons were also slightly different (the 2024 season was warmer), providing conditions that could potentially be less favorable for the establishment of Pseudomonas spp. in 2024. Despite a lower plant growth promotion, this is the second demonstration that inoculation with the DAPG-producing strain P. protegens B21-024 significantly increased lettuce shoot dry weight (when compared to the water treatment) under representative commercial field conditions.
Insufficient rhizosphere colonization by plant-beneficial Pseudomonas spp. has often been suggested as one of the main reasons why disease suppression and/or plant growth promotion fail under field conditions (8, 50). In this study, the population dynamic of both strains in rhizosphere soil was monitored throughout the growing season to confirm that they were able to maintain moderate-to-high population levels. To this end, strain-specific qPCR primers and probes were successfully developed using a comparative genomic-driven approach. The populations of both Pseudomonas strains remained fairly stable throughout the growing season (Fig. 5), with population sizes ranging from 1.1 × 10^4^ to 2.9 × 10^4^ bacteria per gram of dry rhizosphere soil. It is difficult to determine whether such population densities are sufficient to actively promote plant growth under field conditions. By comparison, a population threshold of 10^5^ CFU per gram of fresh root was found to be a prerequisite for the biocontrol of take-all of wheat and Fusarium wilt of radish by plant-beneficial Pseudomonas spp. (51, 52). Such population thresholds are often associated with the accumulation of secondary metabolites in the rhizosphere, which directly inhibit pathogen growth (53, 54). To our knowledge, no study has established such population threshold for direct plant growth promotion by Pseudomonas spp. In our study, it is highly probable that the populations of both strains were much higher in the peat-based growing medium prior to field transplantation, which enabled a maximum plant growth promotion effect at the beginning of the lettuce growth cycle. In addition, we quantified only the rhizosphere soil populations of P. protegens B21-024 and P. putida B21-029, and we did not specifically quantify their presence on the root surface (rhizoplane). Therefore, it is likely that the population sizes reported in this study underestimate the total population of both bacteria found in the vicinity of the plant root system. In any case, the population levels attained in this study confirm that both strains successfully colonized the lettuce root system under field conditions over an entire production cycle when inoculated only once before transplantation. Repeated inoculations with P. protegens B21-024 and P. putida B21-029 should be studied in future experiments to determine if rhizosphere populations may be increased and/or plant-growth promoting effects improved.
One of the most interesting findings of this study was the fact that the rhizosphere population of P. protegens B21-024 was significantly higher when co-inoculated with P. putida B21-029 than when inoculated alone (Fig. S4). This is surprising since each Pseudomonas strain was inoculated at a lower concentration (5 × 10^8^ CFU/mL) in the consortium treatment than when inoculated alone (1 × 10^9^ CFU/mL). Consequently, lower rhizosphere populations were expected. Based on these results, and the absence of inhibition observed between these two strains under in vitro conditions (results not shown), it appears that the two Pseudomonas strains under study are compatible. Moreover, it is possible that P. putida B21-029 could favor the establishment and growth of P. protegens B21-024 in the rhizosphere by yet unidentified mechanisms. In previous studies, it was shown that some Pseudomonas strains were able to alter the composition of root exudates in favor of microbial growth (55, 56). Analysis of the composition of lettuce root exudates revealed an abundance of amino acids, sugar/sugar alcohols, and organic acids (57). It is possible that P. putida B21-029 could degrade some of these compounds into metabolites readily usable by P. protegens B21-024. In addition, P. putida B21-029 carries a complete type III secretion system (T3SS) cluster in its genome (which is not the case for P. protegens B21-024). The role of T3SS in plant-beneficial Pseudomonas spp. is not completely understood, but it is hypothesized that this system could help suppress plant immunity and enhance bacterial survival in the soil environment (58, 59). By reducing the plant immune responses, the T3SS of P. putida B21-029 could perhaps help P. protegens B21-024 to better colonize the lettuce rhizosphere.
In conclusion, we successfully developed a Pseudomonas consortium to promote lettuce growth under representative commercial field conditions. Taken together, the results obtained in this study suggest not only that P. protegens B21-024 and P. putida B21-029 are compatible, but also that they display synergistic activity. In the future, it would be interesting to further study the interaction between both strains to determine whether they directly interact in the rhizosphere, for example, by forming multi-species biofilm, and/or whether they colonize the same niches. This study demonstrates the potential and feasibility of using Pseudomonas consortia for leafy greens growth promotion under representative field conditions.
MATERIALS AND METHODS
Bacterial strains
Two Pseudomonas strains were used in this study, P. protegens B21-024 and P. putida B21-029. P. protegens B21-024 was previously shown to promote the growth of field-grown lettuce plants under varying nitrogen conditions (27). P. putida B21-029 promoted the growth of lettuce plants in a preliminary greenhouse experiment (data not shown). Both strains were originally isolated from agricultural soils collected in Southern Quebec (Canada), and their genomes were sequenced in a previous study (60). The strains were routinely grown in TSB medium (BD Difco, Franklin Lakes, NJ) at 25°C for 24 h under agitation (150 rpm).
Lettuce growth promotion under greenhouse conditions
Romaine lettuce seeds (Lactuca sativa L. cv. Sun Valley) were purchased from Norseco (Laval, QC, Canada) and germinated in 72-cell trays filled with a peat-based growing medium (PRO-MIX BX, Premier Tech, Rivière-du-Loup, QC, Canada). Vermiculite was added as a top coating for seed germination. The plants were grown in a climate-controlled greenhouse under a 16-h photoperiod (16 h of light at 21°C followed by 8 h of darkness at 18°C). The seedlings were watered daily and fertilized two times (20 and 24 days after sowing) with 10 mL of a nutrient solution (N100) containing 0.575 g/L of 6-11-31 fertilizer (Master Plant-Prod Inc., Brampton, ON, Canada), 0.425 g/L of 15.5-0-0 fertilizer (Yara International, Oslo, Norway), and 0.13 g/L of Epsom salt (K+S AG, Kassel, Germany). Thirteen days after sowing, each seedling was inoculated by pipetting 10 mL of a bacterial suspension (or water control) at the base of the plant. The following treatments were used: (i) P. protegens B21-024 (3 × 10^9^ CFU/mL), (ii) P. putida B21-029 (3 × 10^9^ CFU/mL), (iii) combination of B21-024 and B21-029 (5 mL of each bacterial suspension at a concentration of 3 × 10^9^ CFU/mL), and (iv) distilled water (control). The plants were transplanted 27 days after sowing in 2-L plastic pots filled with the same growing medium. The pots were organized in a randomized complete block design with eight replicates per treatment. The plants were fertilized three times (5, 12, and 19 days after transplantation), with 100 mL of a nutrient solution (N200) containing 1.15 g/L of 6-11-31 fertilizer (Master Plant-Prod Inc.), 0.85 g/L of 15.5-0-0 fertilizer (Yara International), and 0.26 g/L of Epsom salt (K+S AG). Fifty-five days after sowing, the plants were harvested, and the aboveground biomass (shoot fresh weight) was measured using a precision scale. The shoots were oven-dried at 65°C for one week (or until completely dried), after which the dry weight was measured.
Lettuce growth promotion under field conditions
Plant material, treatments, and inoculation
Romaine lettuce seeds (Lactuca sativa L. cv. Sun Valley) were germinated in 128-cell trays filled with a peat-based growing medium (PRO-MIX BX, Premier Tech). Vermiculite was added as a top coating for seed germination. The seedlings were grown under the same greenhouse conditions as described above. The plants were watered daily and fertilized two times (at 7 and 22 days after sowing), first with the N100 solution and then with the N200 solution. Fifteen days after sowing, the seedling trays were inoculated with one of the five treatments: (i) P. protegens B21-024, (ii) P. putida B21-029, (iii) combination of B21-024 and B21-029, (iv) fresh TSB medium, and (v) water. The inocula were prepared as previously described (27). Briefly, the bacterial suspensions were first adjusted to an optical density equivalent to 3 × 10^9^ CFU/mL with distilled water. One volume of each bacterial suspension was then mixed with one volume of fresh TSB medium and one volume of distilled water. For the treatment containing the two bacterial strains (B21-024+B21-029), one volume of each bacterial suspension (two volumes in total) was mixed with two volumes of fresh TSB and two volumes of distilled water. The fresh TSB treatment was prepared by mixing two volumes of fresh TSB with one volume of distilled water. The water treatment consisted of distilled water only. The seedling trays were inoculated by soaking them in 5 L of the corresponding inoculum for 5 min. The plants were later transported to the field for transplantation (28 days after sowing).
Field site and experimental design
A field experiment was conducted from June 2024 to July 2024 at the Agriculture and Agri-Food Canada experimental farm located in Sainte-Clotilde, QC, Canada (45.163326, −73.673278). Lettuce plantlets were mechanically transplanted in the field 28 days after sowing and harvested 72 days after sowing. During the 44 days the plants were in the field, there was 145 mm of accumulated precipitation, and the average temperature was 20.2°C. The soil was an organic soil representative of nearby commercial fields used for lettuce production. Total soil nitrogen was 25 g/kg. The field was fertilized once the day before transplantation. This was performed according to local fertilization guidelines (61). The entire field received calcium ammonium nitrate 25-0-0 (80 kg N/ha), triple superphosphate 0-46-0 (30 kg P/ha), and potassium chloride 0-0-60 (145 kg K/ha).
A randomized complete block design was used, with eight blocks in total. Each block was subdivided into five plots, and the treatments were randomized between the plots (Fig. 2). Each plot included 80 lettuce plants, planted with a 40 × 30 cm plant spacing. The plants were irrigated only once at planting. Field work included manual weeding and sampling.
Field sampling
Lettuce plants were collected six times throughout the growing season, at 35 (T1), 41 (T2), 49 (T3), 56 (T4), 63 (T5), and 72 (T6) days after sowing. One randomly selected lettuce plant per plot was sampled during the five intermediate samplings (T1–T5). Five lettuce plants per plot were harvested at the final sampling (T6). Four growth metrics (shoot fresh weight, lettuce length, and number of leaves) were measured. Lettuce shoots were oven-dried at 65°C for 1 week (or until completely dried), after which the dry weight was measured. All measurements were compiled in a database using the ArcGIS Survey123 (ESRI, Redlands, CA) software installed on a digital tablet (iPad Mini, Apple, Cupertino, CA).
Biomass growth estimation using UAV imaging
In addition to manual sampling, UAV imaging was also used to monitor biomass accumulation throughout the growing season. The protocol for crop monitoring leveraged the methodological framework developed by Vigneault et al. (62) for spatiotemporal treatment assessment. Comprehensive aerial surveillance was conducted throughout the complete lettuce development cycle using a DJI Matrice 350 Pro UAV with embedded Zenmuse P1 RGB sensor. Biweekly (twice a week) aerial surveys were performed at precisely calculated altitudes to achieve sub-centimeter spatial resolution—essential for optimal plant segmentation. Flight parameters maintained a minimum 70% image overlap to ensure orthomosaic integrity. Data acquisition accuracy was ensured through a rigorous two-component calibration method: (i) geometric calibration using precisely geolocated ground control points established with RTK GNSS technology (offering millimetric horizontal and centimetric vertical precision); and (ii) photogrammetric processing via Pix4Dmapper software to generate high-precision orthomosaics and digital elevation models at each temporal interval.
For analytical processing, a pre-trained PointRend deep learning architecture was implemented to execute automated segmentation of individual lettuce specimens from the orthomosaic data sets. The extracted morphological parameters (surface area, height, and volumetric measurements) were systematically compiled into a comprehensive database and modeled according to logistic growth functions as per established protocols (62, 63). This approach facilitated quantification of critical growth parameters—initial acceleration, growth rates, deceleration phase, and maximum growth potential—enabling precise assessment of cumulative biomass production and maturity indices across experimental treatments. The AUGC metric, calculated using surface area measurements, was determined to be the optimal parameter for quantifying cumulative treatment effects. This integrated methodology delivered robust, nuanced insights into lettuce growth responses through the synthesis of advanced remote sensing technology and rigorous analytical procedures.
Detection and quantification of the two Pseudomonas strains in rhizosphere soil using qPCR
Soil sampling and DNA extraction
At each sampling time, eight rhizosphere soil samples were collected for each treatment (one per block). Rhizosphere soil was manually harvested by collecting 20 mL of soil adhering to the lettuce root system. The samples were kept at −80°C and lyophilized using a FreeZone 6 Liter Console Freeze Dryers (Labconco, Kansas City, MO). DNA extraction was performed using the DNeasy PowerSoil Pro Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DNA samples were diluted 10-fold with distilled water and kept at −20°C.
Primer-probe design
To quantify the abundance of the two bacterial strains in rhizosphere soil throughout the growing season, we designed strain-specific qPCR primers and probes. First, we used a collection of 40 Pseudomonas strains to identify, using comparative genomics, potential coding DNA sequence (CDS) targets. The aim was to find CDSs that are only present in one strain and absent from the other 39 strains so that these CDSs might be used as targets for qPCR amplification. Potential CDS targets were identified for P. protegens B21-024 and P. putida B21-029 using the EDGAR web server (64, 65). The absence of these CDSs in other microorganisms was checked using BLASTn searches against the non-redundant nucleotide (nr/nt) and the whole-genome sequencing (wgs) databases. Two putative CDS targets were identified (one for each strain). Primers and TaqMan minor groove binder (MGB) probes targeting these CDSs were designed with Primer Express 3.0 (Thermo Fisher Scientific, Waltham, MA) using default parameters. The MGB probe was labeled with a 6-FAM reporter dye at the 5′ end, and a nonfluorescent quencher (NFQ) at the 3′ end. The primers and probes used in this study are listed in Table 1. The PCR primers were custom synthesized by Integrated DNA Technologies (Coralville, IA). The TaqMan MGB probes were purchased from Thermo Fisher Scientific.
Validation of the primer-probe sets’ specificity
The specificity of the two primer-probe sets was first validated in silico with BLASTn against the nr/nt and wgs databases using the amplicon sequences as baits. Primer-BLAST (66) was also used. In addition, the DNA of 20 closely related Pseudomonas strains was extracted using the DNeasy UltraClean microbial kit (Qiagen, Toronto, ON, Canada). The strains used are listed in Table S1. The DNA samples were diluted 10-fold and kept at −20°C. These DNA samples were used to validate the specificity of the primer-probe sets, and specific amplification was only detected when using DNA extracted from the two strains under study. The absence of both strains in rhizosphere soil samples from the TSB- and water-inoculated lettuce plants was also validated by qPCR.
Standard curves for absolute quantification
For each target CDS, a MiniGene containing the amplicon sequence was purchased from Integrated DNA Technologies. Serial dilutions were performed to generate a standard curve ranging from 10^1^ to 10^8^ amplicon copies per µL. This enabled absolute quantification of the target genes.
qPCR
qPCR was performed using an AriaMx real-time PCR system (Agilent, Santa Clara, CA) and the iTaq universal probe supermix kit (Bio-Rad Laboratories, Hercules, CA). Each 10 µL reaction contained 2 µL of template DNA, 5 µL of iTaq universal probe supermix (1×), 0.8 µL of the probe, forward and reverse primers (final concentration of 200 Nmol/L), and 0.6 µL of sterile distilled water. The cycling conditions were 95°C for 2 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Negative-control reactions were performed during each qPCR run by using sterile distilled water instead of template DNA. Each sample was analyzed in triplicate. Absolute target gene copy numbers were adjusted according to rhizosphere soil dry weight and dilution factor. As each target gene is only present in one copy in the genome of the bacteria under study, the values obtained in gene copy number per gram of dry soil can be converted to bacteria per gram of dry soil.
Statistical analysis
Statistical analyses were performed in RStudio 2024.09.0 using the package “agricolae” 1.3-7 (Statistical Procedures for Agricultural Research, Felipe de Mendiburu). The non-parametric Friedman test was used to compare treatments in the greenhouse and field experiments. For the field experiment, the five treatments were compared to each other for each sampling time separately. The Fisher’s least significant difference test was used as a post hoc test (α = 0.05). To compare the abundance of the two strains in the soil throughout the growing season, one-way analysis of variance (ANOVA) and two-way ANOVA were used. The normality was validated using the Shapiro-Wilk test, and the equality of variance was verified with the Bartlett test (or by plotting residuals). Tukey’s honestly significant difference test was used as a post hoc test (α = 0.05).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Palleroni NJ. 2005. Genus Pseudomonas, p 323–379. In Brenner D, Krieg NR, Staley JT (ed), Bergey’s manual of systematic bacteriology volume 2: the proteobacteria, part B: the gammaproteobacteria, 2nd ed. Springer, New York.
- 2Girard L, Lood C, Höfte M, Vandamme P, Rokni-Zadeh H, van Noort V, Lavigne R, De Mot R. 2021. The ever-expanding Pseudomonas genus: description of 43 new species and partition of the Pseudomonas putida Group. Microorganisms 9:1766. doi:10.3390/microorganisms 908176634442845 PMC 8401041 · doi ↗ · pubmed ↗
- 3Xin X-F, Kvitko B, He SY. 2018. Pseudomonas syringae: what it takes to be a pathogen. Nat Rev Microbiol 16:316–328. doi:10.1038/nrmicro.2018.1729479077 PMC 5972017 · doi ↗ · pubmed ↗
- 4Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, Ronald P, Dow M, Verdier V, Beer SV, Machado MA, Toth I, Salmond G, Foster GD. 2012. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol 13:614–629. doi:10.1111/j.1364-3703.2012.00804.x 22672649 PMC 6638704 · doi ↗ · pubmed ↗
- 5Haas D, Défago G. 2005. Biological control of soil-borne pathogens by fluorescent Pseudomonads. Nat Rev Microbiol 3:307–319. doi:10.1038/nrmicro 112915759041 · doi ↗ · pubmed ↗
- 6Mercado-Blanco J, Bakker PAHM. 2007. Interactions between plants and beneficial Pseudomonas spp.: exploiting bacterial traits for crop protection. Antonie Van Leeuwenhoek 92:367–389. doi:10.1007/s 10482-007-9167-117588129 · doi ↗ · pubmed ↗
- 7Höfte M. 2021. The use of Pseudomonas spp. as bacterial biocontrol agents to control plant disease. In Köhl J, Ravensberg WJ (ed), Microbial bioprotectants for plant disease management https://doi.org/10.19103/AS.2021.0093.11. Burleigh Dodds Science Publishing, Sawston.
- 8Lugtenberg B, Kamilova F. 2009. Plant-growth-promoting rhizobacteria. Annu Rev Microbiol 63:541–556. doi:10.1146/annurev.micro.62.081307.16291819575558 · doi ↗ · pubmed ↗