First report of an Escherichia coli ST131 clinical isolate co-harboring blaKPC-2 and blaNDM-13 on an IncB/O/K/Z plasmid in China
Yi-Yu Lyu, Jie-Hao Tai, Cui-Ying Guo, Yin-Yin Zhang, Yao Chen, Qiang Zhou, Wen-Wen Chu, Yi-Le Wu

TL;DR
A dangerous E. coli strain in China carries two antibiotic resistance genes on a stable plasmid, raising concerns about global spread.
Contribution
First report of blaKPC-2 and blaNDM-13 co-occurring on an IncB/O/K/Z plasmid in a clinical E. coli isolate.
Findings
The plasmid pB5-KPC-NDM showed high stability and low fitness cost in recipient bacteria.
blaKPC-2 and blaNDM-13 were found within specific transposon structures that promote horizontal gene transfer.
The isolate B5 is resistant to most antibiotics except tigecycline and colistin.
Abstract
In carbapenem-resistant Enterobacteriaceae, the co-occurrence of carbapenem resistance genes poses a significant threat to global public health. This study investigated the phenotypic and genotypic characteristics of a clinical carbapenem-resistant Escherichia coli strain B5, which harbors both blaKPC-2 and blaNDM-13. Antimicrobial susceptibility testing and plasmid conjugation assays were performed on isolate B5, using E. coli J53 (a standard recipient strain resistant to sodium azide) as the recipient, whereas passaging experiments and growth rate determination were conducted on J53 (pB5-KPC-NDM). Genetic characteristics of B5 were analyzed via whole-genome sequencing (WGS). B5 exhibits an extensive multidrug resistance phenotype, with susceptibility only to tigecycline and colistin. WGS revealed that B5 belongs to ST131, carries 11 plasmids, and co-harbors blaKPC-2 and blaNDM-13 on…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
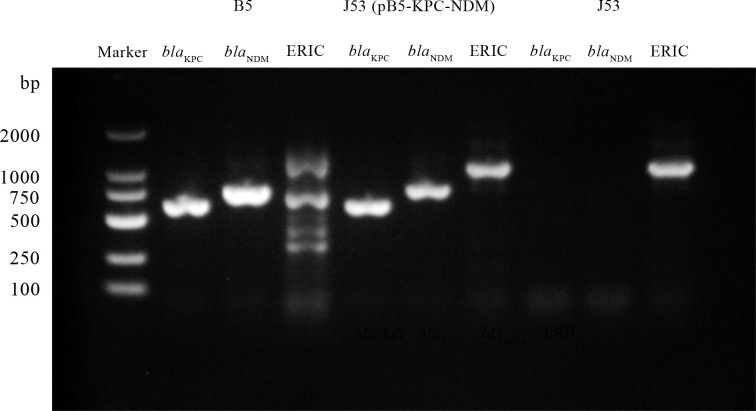 Fig 1
Fig 1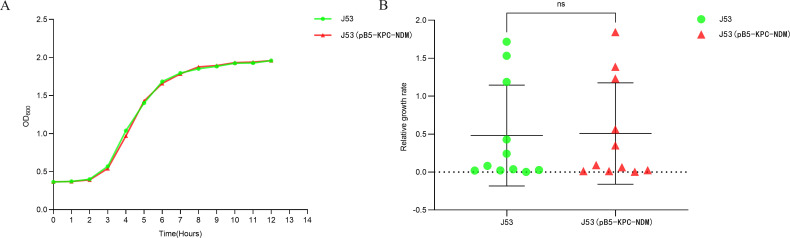 Fig 2
Fig 2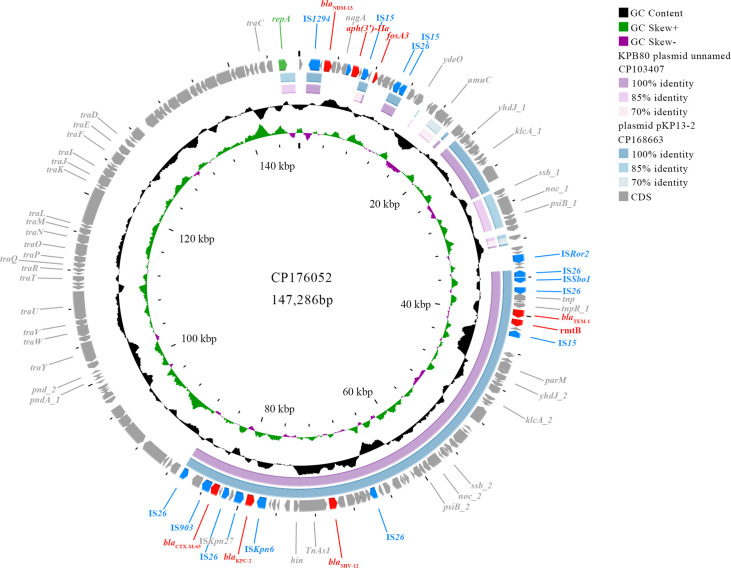 Fig 3
Fig 3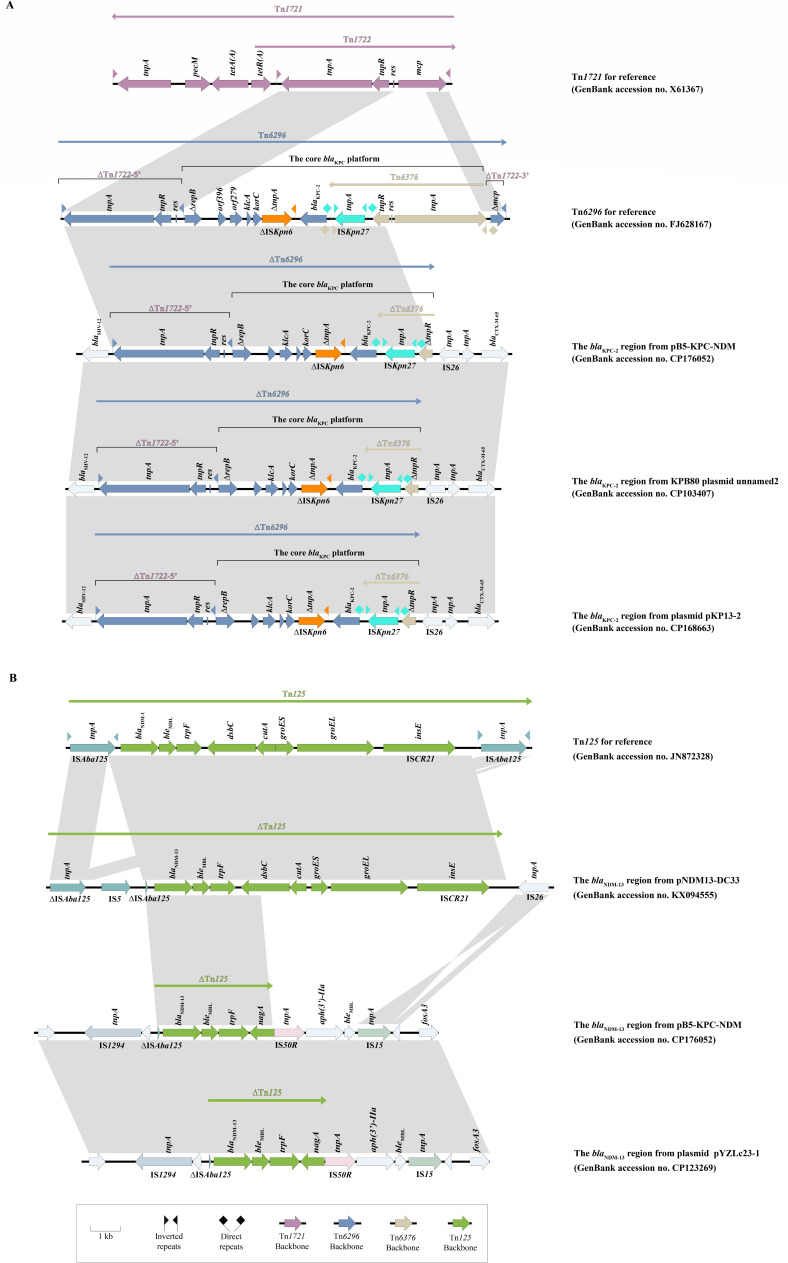 Fig 4
Fig 4| Isolates | AMC | TZP | CXM | FOX | CAZ | CRO | FEP | ETP | IPM | MEM | AMK | LVX | TGC | CST |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B5 | >32/16 | >64/4 | >16 | >16 | >32 | >32 | >16 | >2 | >8 | >8 | >32 | >8 | ≤1 | ≤1 |
| J53 (pB5-KPC-NDM) | >32/16 | >64/4 | >16 | >16 | >32 | >32 | >16 | >2 | >8 | >8 | >32 | ≤1 | ≤1 | ≤1 |
| J53 | ≤8 | ≤4/4 | ≤4 | ≤4 | ≤1 | ≤1 | ≤1 | ≤0.25 | 0.5 | ≤0.125 | ≤8 | ≤1 | ≤1 | ≤1 |
| Genetic material | Accession number | Plasmid type | Size (bp) | GC content (%) | Antimicrobial resistance genes |
|---|---|---|---|---|---|
| Chromosome |
| – | 5,117,658 | 50.64 | |
| pB5-KPC-NDM |
| IncB/O/K/Z | 147,286 | 53.93 | |
| Plasmid2 |
| IncFIA/B/IncFII | 92,949 | 50.88 | |
| Plasmid3 |
| ColRNAI | 10,060 | 55.06 | |
| Plasmid4 |
| Col156 | 5,167 | 47.51 | |
| Plasmid5 |
| Col3M | 4,270 | 47.14 |
|
| Plasmid6 |
| – | 4,077 | 49.91 | |
| Plasmid7 |
| ColpEC648 | 4,063 | 51.42 | |
| Plasmid8 |
| – | 2,953 | 64.75 | |
| Plasmid9 |
| Col (BS512) | 2,101 | 47.12 | |
| Plasmid10 |
| Col (MG828) | 1,597 | 49.91 | |
| Plasmid11 |
| Col (MG828) | 1,549 | 51.00 |
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Natural science research project funding of higher education institutions of Anhui province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial Genetics and Biotechnology · Escherichia coli research studies
INTRODUCTION
Antimicrobial resistance has emerged as a critical global public health threat, leading to significant limitations in therapeutic options (1, 2). Carbapenems are the last resort antibiotics for the treatment of multidrug-resistant Enterobacteriaceae infections (3). With the extensive use of carbapenems, carbapenem-resistant Enterobacteriaceae (CRE) have emerged as a challenging issue and garnered widespread attention (4). Escherichia coli is one of the most prevalent Enterobacteriaceae in both hospital and community environments, capable of causing various serious infections (5). According to recent data from the 2023 China Antimicrobial Resistance Surveillance System, E. coli has become the first and most commonly isolated bacterium in clinics in China, and the resistance rate to carbapenems was 1.7% (6). It causes numerous adverse health effects on patients and imposes a considerable societal burden (3, 7). The primary mechanism of drug resistance gene transmission in CRE is due to the horizontal transfer of mobile genetic elements on plasmids that encode enzymes responsible for hydrolyzing carbapenems and other β-lactam agents. Notably, Klebsiella pneumoniae carbapenemase (KPC) and New Delhi metallo-β-lactamase (NDM) are the most commonly represented carbapenemases found in both inpatient and environmental CRE samples (3).
KPC is classified as class A according to the Ambler classification system and predominantly occurs in carbapenem-resistant K. pneumoniae (CRKP) (8) and is widely distributed worldwide (3). Currently, there are more than 150 known blaKPC subtypes, with the main novel blaKPC variety identified as a mutation of blaKPC-2 (8). It has been reported that the blaKPC gene can be carried by various plasmids, including IncFII, IncI2, IncX, IncA/C, IncN, and IncR (8–10). NDM, which belongs to class B β-lactamase, has at least 48 variants listed in the NCBI database. The blaNDM-13 is a variant characterized by two amino acid substitutions (D95N and M154L) compared to blaNDM-1, which results in increased hydrolytic activity against cefotaxime (11). The blaNDM is primarily carried by plasmids, including IncHI1, IncHI2, IncHI3, IncX3, IncX1, and IncN (12). K. pneumoniae and E. coli are significant carriers of the blaNDM (12). Recently, the coexistence of blaKPC and blaNDM in bacteria, including K. pneumoniae and E. coli, has garnered more attention (13, 14).
This study provides the first evidence of blaKPC-2 and blaNDM-13 co-occurring on an IncB/O/K/Z plasmid, as well as the first detection of these genes in a clinical E. coli isolate (B5). A series of experiments and whole-genome sequencing (WGS) were employed to analyze the phenotypic and genotypic characteristics of the B5, highlighting the urgent need for surveillance of the potential widespread dissemination of such a concerning isolate and its mobile genetic elements on plasmids in the future.
MATERIALS AND METHODS
Bacterial isolate
During a 1-year multicenter CRE surveillance study involving 10 intensive care units (ICUs) from July 2023 to June 2024 in Anhui Province, China, an E. coli isolate (designated B5) co-harboring blaKPC-2 and blaNDM-13 was identified. The isolate was obtained from a rectal swab specimen of a 77-year-old female patient during active CRE screening upon admission to the ICU at Bengbu First People’s Hospital, Bengbu City, Anhui Province, China, on 25 July 2023. The patient presented with pulmonary infection symptoms following transfer from another medical facility, with a documented history of recent hospitalization and mechanical ventilation. Subsequent phenotypic and genomic analyses were conducted to characterize isolate B5.
Species identification and antimicrobial susceptibility testing
Species identification was performed using the Microflex LT automated microbial identification system (Bruker Daltonics, Germany). Antimicrobial susceptibility testing was performed using the Vitek2 Automated System (BioMérieux, France). Minimum inhibitory concentrations (MICs) for 14 antimicrobial agents, including β-lactam/β-lactamase inhibitor combinations (amoxicillin-clavulanate, piperacillin-tazobactam), cephalosporins (cefuroxime, cefoxitin, ceftazidime, ceftriaxone, and cefepime), carbapenems (ertapenem, imipenem, meropenem), amikacin, and levofloxacin, were assessed via broth microdilution following CLSI guidelines (15). EUCAST MIC breakpoints were applied for tigecycline and colistin interpretation (16). E. coli ATCC 25922 served as the quality control strain.
Verification of the transferability of the plasmid carrying blaKPC-2 and blaNDM-13
A plasmid conjugation experiment was conducted to assess the transferability of the plasmid harboring blaKPC-2 and blaNDM-13, utilizing B5 as the donor isolate and sodium azide-resistant E. coli J53 as the recipient isolate. Donor and recipient isolates were, respectively, inoculated into fresh Luria–Bertani (LB) broth (Sangon Biotech, Shanghai, China) to prepare a 1.0 McFarland turbidity standard suspension (3 × 10^8^ CFU/mL). Then, they were mixed at a 1:2 donor-to-recipient ratio and incubated on LB agar plates at 37°C for 12 to 24 h. J53 (pB5-KPC-NDM) colonies were selected on Mueller-Hinton (MH) agar plates (Oxoid, Hampshire, UK) containing sodium azide (296 µg/mL) and meropenem (10 µg/mL). Target gene segments of blaKPC-2 and blaNDM-13 in J53 (pB5-KPC-NDM) isolates were amplified by polymerase chain reaction (PCR), and the PCR results were then subjected to agarose gel electrophoresis. The primers used in the PCR analysis were blaKPC-2 (5′-GTATCGCCGTCTAGTTCTGC-3′, 5′-GGTCGTGTTTCCCTTTAGCC-3′), blaNDM-13 (5′-ATGGAATTGCCCAATATTATGCAC-3′, 5′-TCAGCGCAGCTTGTCGGC-3′), and enterobacterial repetitive intergenic consensus (ERIC) (5′-AAGTAAGTGACTGGGGTGAGCG-3′, 5′-ATGTAAGCTCCTGGGGATTCAC-3′). Electrophoresis was performed with the BG-Power600i System (Boygene, Beijing, China) using 1.0% agarose in Tris Acetate-EDTA buffer at 100 V. The gel electrophoresis profiles were analyzed using Tanon-1600 (Tanon, Shanghai, China).
Stability of the plasmid carrying blaKPC-2 and blaNDM-13
Plasmid retention in J53 (pB5-KPC-NDM) was evaluated through serial passaging in antibiotic-free LB broth. Three single colonies were inoculated into 2 mL LB broth; cultures were diluted 1:1,000 daily in fresh medium and passaged for 10 days. Every fifth day, cultures were serially diluted in PBS and plated on MH agar without antibiotics. Fifty colonies per time point were screened by PCR for blaKPC-2 and blaNDM-13. Plasmid retention frequency was calculated as (positive clones/50) × 100%.
Determination of growth rate
Fitness costs associated with plasmid carriage were assessed by comparing the growth curves of J53 (pB5-KPC-NDM) and recipient J53. Overnight cultures were diluted 1:100 in LB broth, aliquoted into 96-well plates, and incubated with shaking at 37°C. Optical density (OD_600_) was measured hourly for 12 h. Growth curves were analyzed via one-way ANOVA (GraphPad Prism v10.0), with P <0.05 considered statistically significant.
Whole-genome sequencing and analysis
Next-generation sequencing was utilized to sequence and assemble the genome. Genomic DNA was extracted using a plant genomic DNA extraction kit (Tiangen, DP305). A library was prepared and sequenced on an Illumina NovaSeq platform (Illumina Inc., San Diego, CA, USA), generating 150 bp paired-end reads. Quality-controlled reads were assembled de novo using Unicycler v0.5.0 (https://github.com/rrwick/Unicycler). The resulting assemblies were integrated to produce a complete sequence. Gene prediction was conducted using Prokka v1.12 software (https://github.com/tseemann/prokka).
Third-generation sequencing was employed for genome sequencing and assembly to further investigate the genetic background of plasmids in isolate B5 and J53 (pB5-KPC-NDM), elucidating their formation pathways. Genomic DNA was extracted using a modified cetyltrimethyl ammonium bromide method. Sequencing was performed by Personal Biotechnology Company (Shanghai, China) using both the Nanopore PromethION 48 platform and the Illumina NovaSeq platform. The resulting data were assembled using Flye v2.9.1 (https://github.com/fenderglass/Flye) and Unicycler v0.5.0. The assemblies were then integrated to generate a complete sequence, which was further polished using Pilon v1.24 software (https://github.com/broadinstitute/pilon?tab=readme-ov-file) to achieve the final high-quality genome sequence.
Genome component prediction and analysis
Pair-wise average nucleotide identity (ANI) values were calculated using FastANI, with organisms belonging to the same species typically showing ≥95% ANI among themselves (17). Multilocus sequence typing (MLST), antimicrobial resistance genes, and plasmid replicons were identified using the Center for Genomic Epidemiology tools (MLST 2.1, ResFinder 4.1, and PlasmidFinder 1.3 at http://www.genomicepidemiology.org/). Comparative analysis of the IncB/O/K/Z plasmid against reference sequences (GenBank accession no. CP103407, CP168663, CP123269, X61367, FJ628167, JN872328, KX094555) was performed via BLASTn v2.4.0 (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and The Transposon Registry (https://transposon.lstmed.ac.uk/tn-registry). Transposon and insertion sequence (IS) elements were scanned using the ISFinder database (https://www-is.biotoul.fr/index.php). Genomic maps were generated using Proksee (https://proksee.ca/), and linear plasmid comparisons were visualized with Easyfig 2.2.5 (https://mjsull.github.io/Easyfig/files.html).
RESULTS
Antimicrobial susceptibility test
Antimicrobial susceptibility testing revealed that the isolate donor strain B5 exhibited extensive drug resistance to all tested antibiotics except tigecycline and colistin (Table 1). In contrast, the recipient strain J53 demonstrated full susceptibility to the same antimicrobial panel. The J53 (pB5-KPC-NDM) displayed significantly elevated MIC values compared to J53, confirming the successful horizontal transfer of a plasmid carrying resistance genes, including blaKPC-2 and blaNDM-13. While B5 and J53 (pB5-KPC-NDM) shared nearly identical resistance profiles, differential susceptibility to levofloxacin was observed between the two isolates. B5 was resistant to levofloxacin (MIC > 8 mg/L), whereas the J53 (pB5-KPC-NDM) was susceptible (MIC ≤ 1 mg/L), and so is J53.
Genetic features of B5
The total length of the B5 gene sequences was 1,090,782,928 bp, with an average GC content of 50.55%. WGS analysis indicated that B5 contained a 5,117,658 bp chromosome and 11 plasmids (Table 2). ANI analysis was performed between strain B5 and reference genomes of various species using FastANI software. The highest ANI value (96.79%) was observed with E. coli str. K-12 substr. MG1655 (NC_000913.3). MLST analysis showed that B5 belonged to ST131. Antimicrobial resistance genes were identified, including carbapenemases (blaKPC-2 and blaNDM-13), aminoglycosides (aph(3’)-IIa and rmtB), β-lactams (blaSHV-12, blaTEM-1B, and blaCTX-M-65), quinolones (qnrD1), and fosfomycin (fosA3) (Table 2). Notably, almost all antibiotic resistance genes, including blaKPC-2 and blaNDM-13, were located on pB5-KPC-NDM, while the qnrD1 gene is found on plasmid5.
Transferability and stability of pB5-KPC-NDM
Conjugation experiments confirmed the horizontal transfer of pB5-KPC-NDM from donor strain B5 to recipient strain J53, with blaKPC-2, blaNDM-13, and ERIC sequences of J53 detected in transconjugants (Fig. 1). Analysis of the third-generation sequencing result for the transconjugant demonstrated the presence of a complete plasmid, which was transferred to J53 with a sequence identical to pB5-KPC-NDM found in the donor strain B5. Furthermore, results from serial passaging experiments demonstrated that the stability retention rate of pB5-KPC-NDM in transconjugants was 88% (44/50) on day 5 and 74% (37/50) on day 10 in an antibiotic-free environment.
Amplifications of blaKPC, blaNDM, and ERIC sequences of E. coli isolate B5, J53 (pB5-KPC-NDM), and E. coli isolate J53.
Fitness cost of J53 (pB5-KPC-NDM)
J53 (pB5-KPC-NDM) exhibited no significant growth rate reduction relative to J53 (P = 0.972), indicating minimal fitness cost associated with acquisition of the pB5-KPC-NDM plasmid by J53 (pB5-KPC-NDM) (Fig. 2).
(A) Growth curves of J53, J53 (pB5-KPC-NDM). (B) Relative growth rates of J53, J53 (pB5-KPC-NDM).
Genetic characteristics of the IncB/O/K/Z plasmid pB5-KPC-NDM
pB5-KPC-NDM was 147,286 bp in size and had a GC content of 53.93% (Table 2). As shown in Fig. 3, pB5-KPC-NDM harbors various resistance genes alongside critical functional elements, including the replication initiator gene (RepB), conjugative transfer gene cluster (Tra), and IS elements. Comparative genomics using pB5-KPC-NDM as a reference showed 47% maximum sequence homology with related plasmids in NCBI databases, with all top 48 matches originating from K. pneumoniae isolates. Among these, two plasmids—plasmid pKP13-2 (CP168663) isolated from a K. pneumoniae in Jiangxi, and KPB80 plasmid unnamed2 (CP103407) isolated from a K. pneumoniae in Shanghai—exhibited 47% query coverage and 100% nucleotide identity when compared to pB5-KPC-NDM (Fig. 3). Both plasmid pKP13-2 and KPB80 plasmid unnamed2, belonging to the IncFII replicon type, carried blaKPC-2 from K. pneumoniae isolates that do not possess blaNDM-13. In this study, the blaKPC-2 region of pB5-KPC-NDM is approximately 10.5 kb in size and consists of a derivative of Tn6296, which was the same as the blaKPC-2 genetic environment section of plasmid pKP13-2 and KPB80 plasmid unnamed2 (Fig. 4A). Tn6296 arose through insertion of the core blaKPC-2 genetic platform (Tn6376-blaKPC-2-ISKpn6-korC-klcA-repB) into Tn1722, resulting in truncation of the mcp gene. This insertion truncated mcp and split Tn1722 into two segments: Tn1722-5′ (containing the left inverted repeat, tnpA, tnpR, and res) and Tn1722-3′ (containing the truncated mcp and the right inverted repeat). In this study, the blaKPC-2 gene in pB5-KPC-NDM was flanked by ∆Tn6376 (containing tnpR-ISKpn27) and ∆ISKpn6 (a member of the IS1182 family, which was originally 1,540 bp long but reduced to 981 bp), and was followed by the korC-klcA-∆repB genes. Comparison of the blaKPC region in pB5-KPC-NDM with the full Tn6296 sequence revealed that the 558 bp tnpR gene of Tn6376 in pB5-KPC-NDM was truncated to 402 bp. Additionally, the ∆Tn6376-5′ (tnpA-res) and ∆Tn1722-3′ regions were deleted and replaced by IS26. Other β-lactamase resistance genes, including blaSHV-12 and blaCTX-M-65, coexisted around the blaKPC-2 of pB5-KPC-NDM and the compared plasmids (plasmid pKP13-2 and KPB80 plasmid unnamed2).
Characterization of the pB5-KPC-NDM of B5 with closely related plasmids. Plasmid structure of pB5-KPC-NDM (CP176052). pB5-KPC-NDM was used as the reference plasmid to perform genome alignment with the KPB80 plasmid unnamed2 (CP103407) and plasmid pKP13-2 (CP168663).
(A) The blaKPC-2 accessory regions in pB5-KPC-NDM, and comparison with Tn1721, Tn6296, and Tn6376. (B) The accessory blaNDM-13 regions in pB5-KPC-NDM, and comparison with Tn125. Genes are denoted by arrows. Genes, mobile elements, and other features are colored based on function classification. Shading denotes regions of homology (>95% nucleotide identity). The GenBank accession numbers of Tn1721, Tn6296, KPB80 plasmid unnamed2, plasmid pKP13-2, Tn125, pNDM13-DC33, and pYZLc23-1 are X61367, FJ628167, CP103407, CP168663, JN872328, KX094555, and CP123269, respectively.
In addition, further analysis showed that the surrounding genetic environment of blaNDM-13 carried by pB5-KPC-NDM partially matched a previously reported blaNDM-13-harboring plasmid from pYZLc23-1 (75% query coverage and 96.4% nucleotide identity, CP123269). The genetic environment of blaNDM-13 was revealed with a 4,041 bp conserved gene arrangement of IS1294-∆ISAba125-blaNDM-13-bleMBL-trpF-nagA, which was also detected in pYZLc23-1 (Fig. 4B). The IS1294 was located upstream of blaNDM-13, and bleMBL was located downstream of blaNDM-13. The genes trpF (encoding a phosphoribosyl anthranilate isomerase) and nagA (coding for the enzymes of N-acetylglucosamine uptake and metabolism) were present. The complete Tn125 structure was flanked by two ISAba125 elements, with a central region consisting of blaNDM-1-bleMBL-trpF-dsbC-cutA-groES-groEL-ISCR21. Analysis of the first reported complete sequence of the blaNDM-13-harboring plasmid (pNDM13-DC33) revealed that all gene structures of Tn125 were present in the vicinity of blaNDM-13, with the distinction that the truncated ISAba125 was distributed on one side of blaNDM-13. In comparison to the complete Tn125 structure, pB5-KPC-NDM retained a partial Tn125 structure (blaNDM-13-bleMBL-trpF-nagA), with a truncated ISAba125 upstream of blaNDM-13. This ∆ISAba125 shared 95% homology with the truncated ISAba125 in pNDM13-DC33. Notably, in the present study, IS1294 replaced ISAba125 and was located upstream of blaNDM-13, differing from the genetic context of blaNDM-13 in pNDM13-DC33.
DISCUSSION
It is extremely concerning when antimicrobial resistance genes spread via mobile genetic elements, and one of the most crucial ways is through the horizontal transfer of plasmids (18). WGS analysis of the B5 isolate revealed distinct genetic features compared to previously reported blaKPC-2 or blaNDM-13 carrying isolates. These variations are primarily attributed to distinct genomic architectures among isolates, particularly the differential arrangements of mobile genetic elements flanking resistance determinants and associated ISs within their chromosomal contexts (19–21). Notably, pB5-KPC-NDM co-harboring blaKPC-2 and blaNDM-13 maintained stability in B5 despite these genetic background variations. To our knowledge, this study provides the first evidence of blaKPC-2 and blaNDM-13 co-occurring on an IncB/O/K/Z plasmid, as well as the first detection of these genes in a clinical E. coli isolate (B5).
According to the antimicrobial susceptibility test results, B5 exhibited extensive drug resistance to nearly all commonly used antibiotics except tigecycline and colistin. Due to the simultaneous production of blaKPC-2 and blaNDM-13, neither aztreonam nor any β-lactamase inhibitors (including avibactam) were effective against this isolate (3). Furthermore, the donor strain B5 was resistant to levofloxacin, whereas J53 (pB5-KPC-NDM) exhibited a susceptible phenotype identical to the recipient strain J53. This discrepancy finds a clear explanation in the genomic analysis. The qnrD1 gene, which confers resistance to fluoroquinolones, was located on plasmid5 of the donor strain B5. Crucially, sequencing of the transconjugant confirmed the absence of plasmid5, indicating that it was not co-transferred with the pB5-KPC-NDM plasmid during conjugation. Consequently, the lack of qnrD1 in J53 (pB5-KPC-NDM) accounts for its susceptible phenotype to levofloxacin. The sensitivity of this isolate to colistin and tigecycline suggests their potential as a therapeutic option. Nevertheless, the increasing prevalence of CRE has led to heightened reliance on colistin and tigecycline, which poses a significant risk of developing colistin and tigecycline resistance (22).
Research indicates that blaNDM-carrying E. coli isolates exhibit diverse MLST types without forming dominant clones, with notable diversity observed among the plasmids carrying blaNDM (12). This suggests the polyphyletic origins of blaNDM-positive strains and recurrent horizontal acquisition of blaNDM across bacterial species (12). Since its initial identification on the chromosome of E. coli from Nepal in 2015 (11), the blaNDM-13 has disseminated via multiple plasmid types (e.g., IncX3, IncFIB/IncFII) in E. coli (19–23). In 2003, four isolates of K. pneumoniae obtained from patients at a Maryland medical center were found to produce a novel class A, plasmid-mediated blaKPC-2, which shares 99% identity with blaKPC-1. Compared to K. pneumoniae, blaKPC-2 transmission in E. coli remains comparatively restricted, primarily involving IncP6, IncR, and IncC replicons (10, 24). Notably, previous studies have found that IncB/O/K/Z plasmids are recognized vectors for multidrug resistance genes (e.g., blaCTX-M and aadA5) in Enterobacteriaceae (25–27). The E. coli isolate B5 described in this study co-carries an IncB/O/K/Z plasmid harboring blaKPC-2 and blaNDM-13, thereby increasing the diversity of plasmids associated with blaNDM-13.
In a study cohort comprising up to seven CRKPs co-carrying blaKPC-2 and blaNDM-1, it was demonstrated that these seven CRKPs emerged from a CRKP carrying blaKPC-2 progenitor that subsequently acquired a highly transferable blaNDM-1 plasmid (28). This phenomenon suggests that pB5-KPC-NDM from isolate B5 may represent a fusion of multiple plasmids or mutations in genes, which appears to be a common mechanism by which bacteria acquire multiple carbapenem resistance genes. Structurally, plasmids comprise two distinct functional domains: conserved and variable. The conserved region can be further subdivided into two fragments: one encoding genes related to conjugative transfer and the other encoding backbone genes associated with plasmid replication and stability. In this study, the variable region was rich in IS elements (e.g., ISKpn27, IS26, ISKpn6, IS1294, ISAba125) and various resistance genes, including blaCTX-M-65 and blaSHV-12. Resistance genes can be moved by IS as a component of a composite transposon, which is a region that is surrounded by two identical or related IS copies that can move together (28). Transposons and ISs allow bacteria to migrate within or across DNA molecules and acquire antibiotic resistance determinants, which play a central role in promoting the acquisition and spread of resistance genes (28). Research indicates that the blaKPC gene is associated with diverse mobile genetic elements on transferable plasmids (13, 29). Internationally, blaKPC typically resides within Tn4401, which mediates its horizontal transfer (13). The elements Tn3-tnpR, Tn3-tnpA, ISKpn7, blaKPC, and ISKpn6 in the blaKPC-2 region of pB5-KPC-NDM are all components of the Tn4401 isoform (29). In contrast, a distinct genetic environment for blaKPC predominates in China, where nearly all blaKPC genes localize to Tn3-Tn4401 chimeras. These chimeras may insert into the transposon Tn1721. A recent study discovered that an isolate of K. pneumoniae co-carried blaKPC-2 and blaNDM-13, exhibiting a blaKPC-2 environment on an IncFII/IncR plasmid that was consistent with pB5-KPC-NDM (13). The IncFII/IncR plasmid possessed essential structural characteristics similar to Tn4401 and the Tn1721-based structure, which are two of the most prevalent genetic environments harboring blaKPC-2 in China, the US, and Europe (13). Furthermore, Tn6296—a complex transposon formed by the core blaKPC platform inserted into Tn1722 and co-existing with Tn1721—serves as a key element in the dissemination of blaKPC-2 in pB5-KPC-NDM. The replacement of missing regions ∆Tn6376-5′ (tnpA-res) and ∆Tn1722-3′ (mcp) by IS26 in pB5-KPC-NDM suggests its potential involvement in blaKPC-2 horizontal dissemination.
Tn125, a composite transposon flanked by two ISAba125 elements, facilitates the inter-species transfer of blaNDM among bacteria (30). As a member of the IS30 family, ISAba125 encodes a 322-amino-acid DDE-type transposase and enhances carbapenem resistance by promoting blaNDM-1 expression and transmission (31). Among Enterobacteriaceae, the ISs flanking blaNDM (both upstream and downstream) are diverse, including IS3000, IS26, ISEC33, and ISCR1. These ISs are often deleted or truncated by other ISs, leading to structural variations (30). Despite differences in the ISs that mediate blaNDM-1 transfer, the partial Tn125 structure (ISAba125-blaNDM-1-bleMBL-trpF) remains relatively conserved, as does pB5-KPC-NDM (18, 32). Compared to previous reports (19, 20), IS1294, which was located in the blaNDM-13 conserved genetic sequence in pB5-KPC-NDM of B5, replaces ISAba125. IS1294, which mobilizes via rolling circle replication, participated in the movement of blaCMY-2 (18). Interestingly, this study identifies IS1294, a member of the IS91 family, which may supplant ISAba125 in blaNDM-13 mobilization and propagation, though blaNDM-13 remains underrepresented in surveillance data due to its historical plasmid rarity (32). Recent reports of blaNDM-13 dissemination across species underscore its emerging epidemic potential (13, 19, 20). Consequently, plasmids harboring blaNDM-13 may eventually become widespread through evolutionary processes, and urgent measures must be taken to prevent the further spread of such plasmids.
In addition, the growth rate determination results indicate that pB5-KPC-NDM incurs low fitness costs to the recipient isolate. While a previous study reported a 72% stability rate for blaKPC-2 on the tenth day (21), the similarly noteworthy stability observed in this study (pB5-KPC-NDM) establishes favorable conditions for the dissemination of carbapenem resistance genes. However, only pB5-KPC-NDM was experimentally confirmed in this study for its presence, conjugative transfer, and genetic stability. The nature of the other 10 putative plasmids remains undetermined. The convergence of blaKPC-2 and blaNDM-13 on a transmissible IncB/O/K/Z plasmid constitutes a critical public health threat, mandating intensified surveillance and containment strategies.
Conclusion
This study characterizes a novel IncB/O/K/Z plasmid co-harboring blaKPC-2 and blaNDM-13, demonstrating exceptional conjugation stability in clinical E. coli isolates. Genomic analyses revealed that blaKPC-2 and blaNDM-13 are embedded within mobile genetic architectures containing multiple IS elements, suggesting potential mechanisms for horizontal gene transfer. Notably, this represents the first documentation of blaKPC-2 and blaNDM-13 co-localization on a single plasmid within the IncB/O/K/Z incompatibility group, expanding the known diversity of plasmids carrying blaKPC-2 and blaNDM-13. In addition, the genomic plasticity observed in this hybrid plasmid poses significant clinical challenges. Further research on isolates concurrently harboring multiple carbapenemases is urgently needed, given that these groups may exacerbate the spread of resistance genes and restrict antibiotic treatment options. Implementation of enhanced surveillance protocols by healthcare institutions and regulatory agencies, coupled with the development of targeted intervention strategies, is urgently required to mitigate this emerging resistance crisis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Collaborators AR. 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399:629–655. doi:10.1016/S 0140-6736(21)02724-035065702 PMC 8841637 · doi ↗ · pubmed ↗
- 2Niu G, Li W. 2019. Next-generation drug discovery to combat antimicrobial resistance. Trends Biochem Sci 44:961–972. doi:10.1016/j.tibs.2019.05.00531256981 · doi ↗ · pubmed ↗
- 3Potter RF, D’Souza AW, Dantas G. 2016. The rapid spread of carbapenem-resistant Enterobacteriaceae. Drug Resist Updat 29:30–46. doi:10.1016/j.drup.2016.09.00227912842 PMC 5140036 · doi ↗ · pubmed ↗
- 4Durante-Mangoni E, Andini R, Zampino R. 2019. Management of carbapenem-resistant Enterobacteriaceae infections. Clin Microbiol Infect 25:943–950. doi:10.1016/j.cmi.2019.04.01331004767 · doi ↗ · pubmed ↗
- 5Perez F, Bonomo RA. 2019. Carbapenem-resistant Enterobacteriaceae: global action required. Lancet Infect Dis 19:561–562. doi:10.1016/S 1473-3099(19)30210-531047851 · doi ↗ · pubmed ↗
- 6China Antimicrobial Resistance Surveillance System (CARSS). 2023. National antimicrobial resistance monitoring report 2023 (brief edition). Available from: https://www.carss.cn
- 7Bartsch SM, Mc Kinnell JA, Mueller LE, Miller LG, Gohil SK, Huang SS, Lee BY. 2017. Potential economic burden of carbapenem-resistant Enterobacteriaceae (CRE) in the United States. Clin Microbiol Infect 23:48. doi:10.1016/j.cmi.2016.09.003 · doi ↗
- 8Ding L, Shen S, Chen J, Tian Z, Shi Q, Han R, Guo Y, Hu F. 2023. Klebsiella pneumoniae carbapenemase variants: the new threat to global public health. Clin Microbiol Rev 36:e 0000823. doi:10.1128/cmr.00008-2337937997 PMC 10732083 · doi ↗ · pubmed ↗