Why robust identification of rare yeasts is the need of the hour
Anuradha Marathe, Brittany O'Brien, YanChun Zhu, Mayuri Vaidya, Sudha Chaturvedi

TL;DR
This paper shows how adding rare yeast profiles to a MALDI-TOF MS library improves rapid and accurate identification of these yeasts, which is critical for treating immunocompromised patients.
Contribution
The study creates and validates an in-house MALDI-TOF MS library for 141 rare yeast isolates, improving identification accuracy and speed.
Findings
All 141 rare yeast isolates were identified to the species level with a score of 2.0 or higher using the enriched library.
The ethanol/formic acid extraction method with bead-beating improved protein profiling for some yeast species.
The enriched library's profiles are publicly available via CDC's MicrobeNet database for broader use.
Abstract
Infections caused by rare yeasts are surging, mainly due to the increase in immunocompromised or seriously ill patients. With the increasing clinical significance of rare yeasts, there is an urgent need for a rapid diagnostic method. The current process relies on the PCR of the ribosomal gene’s internal transcribed spacer (ITS) region, followed by sequencing and BLAST search, which are labor- and time-intensive. Matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) has revolutionized the rapid identification of yeasts at the species level. One limitation of MALDI-TOF MS is the inadequacy or lack of main spectrum profiles (MSPs) in the reference library, which results in low scores and no organism identification. The focus of this study was to create an in-house Bruker library for 141 rare yeast isolates, representing 25 genera and 68 species of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
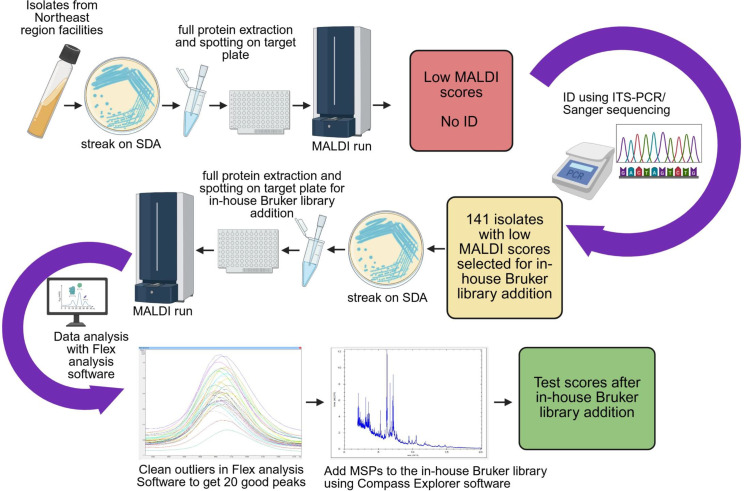 Fig 1
Fig 1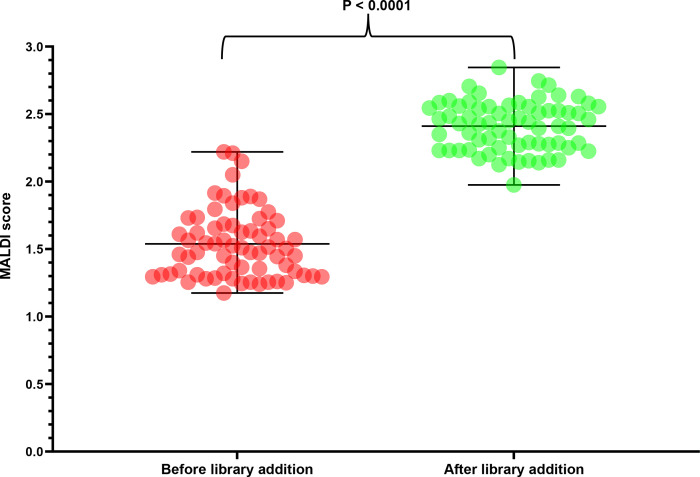 Fig 2
Fig 2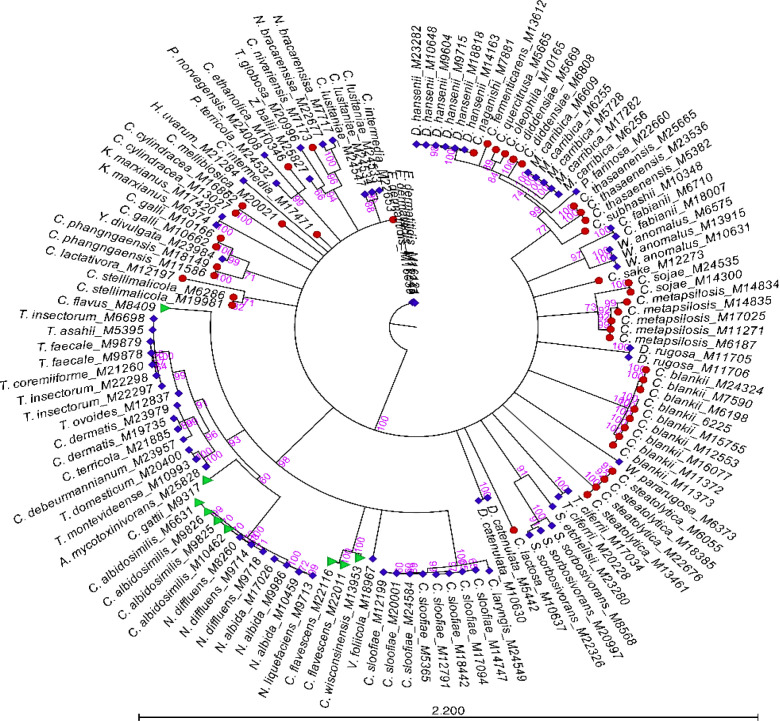 Fig 3
Fig 3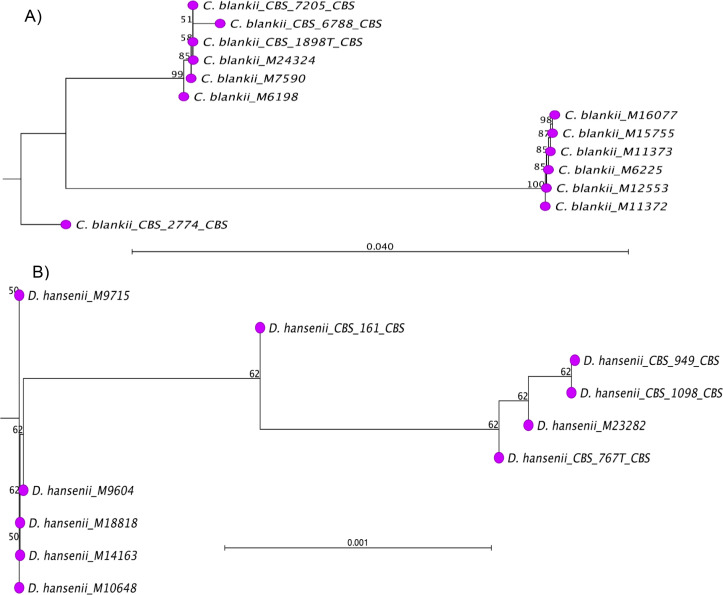 Fig 4
Fig 4| Yeast species (old name) | Yeast species (new name) | # of isolates added to in-house Bruker library | MALDI score before library enrichment | MALDI score after library enrichment | Known to cause human infections |
|---|---|---|---|---|---|
|
| – | 2 | 1.26/1.25 | 2.53/2.58 | Invasive ( |
|
|
| 1 | 1.20/1.28 | 2.29/2.03 | Unknown |
|
| – | 1 | 1.23/1.29 | 2.54/2.58 | Unknown |
|
|
| 7 | 1.59/1.63 | 2.24/2.34 | Invasive ( |
|
| – | 1 | 1.29/1.30 | 2.41/2.52 | Unknown |
|
| – | 1 | 1.24/1.35 | 2.46/2.36 | Unknown |
|
| – | 1 | 1.23/1.39 | 2.47/2.40 | Unknown |
|
| – | 1 | 1.30/1.33 | 2.67/2.64 | Unknown |
|
|
| 1 | 1.22/1.29 | 2.51/2.50 | Unknown |
|
| – | 2 | 1.29/1.31 | 2.51/.260 | Unknown |
|
| – | 1 | 1.54/1.60 | 2.17/2.14 | Invasive ( |
|
| – | 1 | 1.23/1.27 | 2.23/2.23 | Unknown |
|
| – | 1 | 1.39/1.32 | 2.50/2.61 | Invasive ( |
|
|
| 3 | 1.41/1.67 | 2.53/2.55 | Unknown |
|
| – | 4 | 1.66/1.65 | 2.52/2.50 | Unknown |
|
| – | 3 | 1.28/1.28 | 2.50/2.55 | Invasive ( |
|
| – | 1 | 1.14/1.21 | 2.23/2.27 | Invasive ( |
|
| – | 1 | 1.22/1.35 | 2.72/2.77 | Unknown |
|
| – | 3 | 1.50/1.30 | 2.20/2.26 | Unknown |
|
|
| 1 | 1.28/1.23 | 2.20/2.20 | Unknown |
|
|
| 1 | 1.49/1.45 | 2.31/2.33 | Unknown |
|
| – | 1 | 1.54/1.62 | 2.20/2.35 | Invasive ( |
|
|
| 1 | 1.37/1.36 | 2.27/2.05 | Unknown |
|
|
| 9 | 1.58/1.55 | 2.18/2.16 | Unknown |
|
| – | 1 | 1.26/1.23 | 2.40/1.85 | Unknown |
|
|
| 2 | 1.37/1.76 | 2.40/2.39 | Non-invasive ( |
|
| – | 1 | 1.77/1.65 | 2.57/2.33 | Unknown |
|
| – | 3 | 1.51/1.51 | 2.51/2.50 | Invasive ( |
|
| – | 1 | 1.62/1.63 | 2.83/2.86 | Invasive ( |
|
| – | 1 | 1.46/1.22 | 2.62/2.64 | Unknown |
|
| – | 1 | 1.47/1.42 | 2.51/2.34 | Unknown |
| Yeast species (old name) | Yeast species (new name) | # of isolates | # of isolates added to in-house Bruker library | MALDI score before library enrichment | MALDI score after library enrichment | Known to cause human infections |
|---|---|---|---|---|---|---|
|
|
| 2 | 1 | 2.07/2.23 | 2.34/2.36 | Invasive ( |
|
|
| 2 | 1 | 1.66/1.64 | 2.54/2.63 | Invasive ( |
|
|
| 4 | 13 | 1.51/1.54 | 2.29/2.28 | Invasive ( |
|
|
| 3 | 2 | 1.81/1.78 | 2.29/2.27 | Invasive ( |
|
|
| 7 | 2 | 1.66/1.71 | 2.29/2.25 | Invasive ( |
|
|
| 3 | 2 | 1.51/1.52 | 2.47/2.45 | Invasive ( |
|
|
| 1 | 2 | 1.48/1.40 | 2.55/2.47 | Unknown |
|
|
| 8 | 3 | 1.89/1.87 | 2.28/2.22 | Invasive ( |
|
|
| 11 | 8 | 1.54/1.60 | 2.49/2.46 | Invasive ( |
|
|
| 1 | 2 | 1.46/1.46 | 2.58/2.60 | Invasive ( |
|
|
| 9 | 2 | 1.70/1.76 | 2.47/2.15 | Invasive ( |
|
|
| 17 | 2 | 1.89/1.89 | 2.31/2.35 | Invasive ( |
|
|
| 11 | 7 | 1.75/1.80 | 2.49/2.48 | Invasive ( |
|
|
| 8 | 2 | 1.47/1.48 | 2.37/2.51 | Invasive ( |
|
|
| 8 | 1 | 1.48/1.47 | 2.05/1.90 | Invasive ( |
|
|
| 1 | 1 | 1.39/1.25 | 2.57/2.63 | Unknown |
|
|
| 2 | 3 | 2.23/2.21 | 2.57/2.48 | Invasive ( |
|
|
| 10 | 3 | 1.75/1.70 | 2.41/2.45 | Invasive ( |
|
|
| 5 | 3 | 1.52/1.49 | 2.17/2.12 | Invasive( |
|
|
| 3 | 2 | 1.73/1.74 | 2.37/2.38 | Invasive( |
|
|
| 15 | 3 | 1.65/1.62 | 2.59/2.50 | Invasive ( |
|
|
| 2 | 4 | 1.42/1.48 | 2.28/2.18 | Invasive ( |
|
|
| 1 | 3 | 1.34/1.42 | 2.18/2.10 | Invasive ( |
|
|
| 3 | 4 | 1.64/1.55 | 2.32/2.25 | Invasive ( |
|
|
| 2 | 1 | 1.40/1.69 | 2.25/2.20 | Invasive ( |
|
| 15 | 1 | 1.56/1.68 | 2.30/2.64 | Invasive ( | |
|
|
| 1 | 1 | 1.37/1.25 | 2.62/2.54 | Invasive ( |
|
|
| 3 | 8 | 1.87/1.81 | 2.38/2.34 | Invasive ( |
|
|
| 13 | 1 | 1.88/1.86 | 2.32/2.47 | Unknown |
|
|
| 3 | 1 | 1.98/1.81 | 2.42/2.05 | Invasive ( |
|
|
| 10 | 1 | 1.26/1.30 | 2.62/2.51 | Invasive ( |
|
|
| 2 | 1 | 2.17/2.25 | 2.53/2.72 | Invasive ( |
|
|
| 2 | 1 | 1.23/1.38 | 2.63/2.54 | Non-invasive ( |
|
|
| 1 | 2 | 1.46/1.44 | 2.61/2.67 | Invasive ( |
|
|
| 4 | 1 | 1.67/1.68 | 2.69/2.72 | Invasive ( |
|
|
| 2 | 1 | 1.32/1.35 | 2.74/2.69 | Unknown |
|
|
| 3 | 1 | 2.01/2.09 | 2.16/2.18 | Unknown |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Infections and Studies · Antifungal resistance and susceptibility · Yeasts and Rust Fungi Studies
INTRODUCTION
Fungal pathogens are responsible for at least 13 million infections and 1.5 million deaths globally per year, primarily in the immunosuppressed or those hospitalized with serious, underlying diseases (1). Over the last three decades, the incidence of hospital-acquired fungal infections has increased. A single fungal pathogen can infect multiple tissues in an immunocompromised patient and can undergo morphogenic shifts during infection (2). As a result, symptoms of an invasive fungal infection are frequently nonspecific, and early diagnosis is often challenging to establish. Generic therapy carries risks of both treatment-associated toxicity and development of resistance in the pathogens (3). The most common fungal disease in hospitalized patients is invasive candidiasis, which is associated with mortality exceeding 40%, even with treatment (3). The spectrum of pathogens causing invasive candidiasis has also shifted from Candida albicans to non-albicans Candida species. There is also a rise in rare yeasts belonging to Candida or non-Candida genera as emerging threats in the healthcare setting due to their high virulence, proliferation within the body, and their drug-resistance profile (4–6). Therefore, the role of diagnostic mycology laboratories in adapting to the emergence of the new and rare yeasts is more crucial than ever (7).
Existing commercial identification systems, which traditionally identify pathogenic yeasts based on morphological and biochemical characteristics, are time-consuming, have limited databases, and are designed to identify only the more common medically important yeasts (8). On the other hand, the polymerase chain reaction (PCR) and sequencing of the ribosomal genes, the internal transcribed spacer (ITS) and D1/D2, are the gold standard methods for yeast identification (8, 9). These methods are excellent but time-consuming, which can lead to delays in patient care.
Around 2010, the introduction of matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) technology into the clinical microbiology field was a game-changer, as clinical laboratories could identify organisms from crude protein suspensions within minutes, at very low processing costs, and minimal processing time. Over the recent years, MALDI-TOF MS has been successfully used to identify numerous yeast species (9–13). One limitation of MALDI-TOF MS is the absence or inadequacy of main spectrum profiles (MSPs) in the reference library, which can lead to low scores and no organism identification (14, 15). Moreover, cases of infection by newly emerging and newly pathogenic yeasts, including Candida auris, are on the rise (16–20). Another concern is the increasing incidences of rare yeast infections, such as Candida blankii and Debaryomyces hansenii in immunocompetent individuals over the last decade (21, 22). Under these circumstances, improving the MALDI database and having a better representation of the rare and emerging yeasts is the need of the hour and a key factor for the accurate identification of rare yeasts. For instance, in 2016, with the rapid emergence of C. auris as a global threat, many laboratories developed their in-house supplemental databases, which helped them accurately identify C. auris (23–26). This demonstrates that in-house developed databases can significantly improve performance and serve as a valuable measure for rapid response to emerging pathogens, provided they are rigorously validated and interpreted with care (10). In this investigation, we describe the utility of our in-house developed Bruker MALDI-TOF MS library as a powerful tool for identifying rare yeasts, offering speed, accuracy, and clinical relevance.
MATERIALS AND METHODS
For this study, 141 rare yeast isolates representing 68 species and 25 genera, identified by sequencing of the ITS region of the ribosomal gene, were selected. The ITS sequences of these rare yeasts were submitted to GenBank with accession numbers PQ644596https://www.ncbi.nlm.nih.gov/nuccore/PQ644596.1/–PQ644604, PQ644607–PQ644612, PV029103–PV029223, and PV029329. The majority of these isolates could not be identified by MALDI-TOF MS and were added to the library, while few were added for the purpose of library enrichment. All isolates in this investigation were obtained from the Mycology Culture Collection Repository (MCCR) (https://www.wadsworth.org/programs/id/mycology/culture-collection-repository). For MALDI-TOF MS protein extraction, yeasts were grown on Sabouraud dextrose agar (SDA) plates overnight at 30°C. The protein from each yeast isolate was extracted using ethanol/formic acid extraction method (Bruker Daltonics, Germany) (27). In brief, yeast colonies were placed into a 1.5-mL centrifuge tube containing 300 µL of liquid chromatography mass spectrometry (LCMS) grade water and mixed thoroughly, followed by addition of 900 µL of absolute ethanol. The tubes were centrifuged at 13,200 RPM for 2 min at room temperature (RT), the supernatant was discarded, and the pellet was air-dried for 30 min. The air-dried pellet was then mixed with 25 µL of 70% formic acid, incubated at RT for 10 min, followed by addition of 25 µL of acetonitrile, mixing thoroughly, and centrifuging at 13,200 RPM for 2 min at RT. One microliter of the supernatant was spotted on 12 spots of a 96-spot target plate (Bruker Daltonics, Germany). One microliter of Bacterial Test Standard (BTS, Bruker Daltonics, Germany) was spotted on one spot of the plate as a positive control. All spots were air-dried for approximately 5 min, and then each spot was overlaid with 1 µL of α-cyano-4-hydroxycinnamic acid (HCCA) matrix and air-dried completely (~10–15 min) before the MALDI-TOF MS run. Six isolates belonging to five rare yeast species, which failed to yield protein spectra in MALDI, were mixed with 25 µL of 70% formic acid and subjected to bead-beating at 4,700 RPM for 45 s (Precellys24 Homogenizer) and incubated at RT for 10 min followed by protein extraction and supernatant spotting on target plate as described above. MSPs were obtained following the manufacturer’s guidelines by using the MALDI Biotyper software version 3.1 (Bruker Daltonics, Germany). All spectra were scrutinized using the Flex analysis software (Bruker Daltonics, Germany). Spectra with outlier, low-quality, and noisy peaks were deleted, and a minimum of 20 high-quality spectra were selected for each yeast isolate. These spectra were stored in the in-house Bruker library as a reference MSP using Compass Explorer (Bruker Daltonics, Germany). According to Bruker, the resemblance between an unknown specimen spectrum and reference spectra is indicated by a log (score), which will be henceforth referred to as “MALDI score” (28). A MALDI score of ≥1.9 was considered as the cutoff for species-level identification. The entire process of library addition of rare yeasts has been shown in a flowchart (Fig. 1).
Flowchart of library addition of rare yeasts. A flowchart illustrating the entire process of creation of an in-house Bruker library of rare yeasts. The schematic was created with BioRender.com. SDA, Sabouraud dextrose agar.
Statistical analysis of the MALDI data were performed using the built-in analysis feature of Graphpad Prism software (version 9.5.1). The paired t-test was used for comparison of two groups (MALDI scores before addition and MALDI scores after addition). Results were considered significant at a P-value < 0.05.
To determine genotypic differences within the yeast species, ITS sequences were aligned using the Create Alignment function in the QIAGEN CLC Genomics Workbench (version 24.0.2). The neighbor-joining algorithm and Jukes-Cantor nucleotide substitution model were implemented for constructing a phylogenetic tree representing all rare yeasts, as well as several individual phylogenetic trees in the QIAGEN CLC Genomics Workbench (version 24.0.2). All the gaps were excluded from the analysis, and branch support was ascertained using 1,000 bootstrap replicates.
RESULTS AND DISCUSSION
We successfully added 141 yeast isolates representing 68 species and 25 genera to the in-house Bruker library. The library addition resulted in accurate identification of rare yeasts to the species level with a MALDI score of 2.0 or above. The ethanol/formic acid protein extraction was adequate for 135 isolates (95.7%), representing 63 rare yeasts for identification, whereas six isolates (4.3%) representing five species (Candida fermenticarens, Candida sake, Candida terebra, Candida lactosa, Trichomonascus ciferrii) required an additional bead-beating step ([Tables 1 and 2](#T1 T2)). The in-house Bruker library was further evaluated by testing 47 rare yeast isolates in a blinded fashion, and MALDI score of 2.0 or above confirmed the accuracy of the in-house Bruker library (Table S3). In summary, addition of rare yeasts to the in-house Bruker library led to a significant (P < 0.0001) improvement in MALDI score distribution (Fig. 2).
Distribution of MALDI scores of rare yeasts before and after library addition. A column graph was created in GraphPad Prism version 9.5.1 using the scatter plot function, which shows mean and ranges as dots. Red dots represent the mean MALDI score before, and green dots represent the mean MALDI scores after the addition of rare yeasts into the in-house Bruker library.
Of the 68 species of 25 genera of rare yeasts added, 31 species of 11 genera had no reference spectra in the commercial Bruker library. Therefore, the lack of identification for these yeasts by MALDI-TOF MS was not surprising (Table 1). However, despite some of the rare yeasts being adequately represented in the commercial Bruker library, they failed to yield a good score for species-level identification (Table 2). The multiple alignments of ITS sequences revealed single-nucleotide polymorphisms (SNPs) in isolates within a species. Some of these species were C. blankii, Candida cylindracea, Candida diddensiae, Candida stellimalicola, Cystobasidium sloofiae, Naganishia albida, Starmerella sorbosivorans, and Cutaneotrichosporon dermatis, resulting in clustering on different branches of the phylogenetic tree (Fig. 3). These results indicate that there are several genotypes within the species, and library enrichment with all possible genotypes is key for successful identification through MALDI-TOF MS. When ITS sequences of Bruker C. blankii and D. hansenii (formerly known as Candida famata) isolates were compared with ITS sequences of C. blankii and D. hansenii isolates from the northeastern United States, a distinct clustering of Bruker isolates to that of U.S. isolates was observed (Fig. 4A and B). These results indicate the possible cause for low MALDI score in the initial phase of this investigation, and adding local strains to the library yielded successful identification (Table 2). The Bruker isolates were derived from other parts of the globe (Table S1), whereas the local isolates were derived from the northeastern United States (Table S2). Genetic variation in isolates can be influenced by geographical distribution and other factors such as climate, ecological niches, changes in chromosomal copy number, and hyphal network. According to a haplotype and network analysis, high intraspecific variability of ITS sequences was observed in pathogenic yeast species, including Candida albicans, Candida tropicalis, Nakaseomyces glabratus (formerly known as Candida glabrata), and Clavispora lusitaniae (53). Research suggests that species with high genetic diversity are most frequently human commensals, and this finding could explain the existence of additional genetic adaptation within normal microbiota with older evolutionary origins (23, 24).
Phylogenetic analysis of rare yeasts. The phylogenetic tree of 132 isolates of 65 rare yeast species was constructed using the QIAGEN CLC Genomics Workbench (version 24.0.2). Multiple alignment of ITS genes was performed using the Create Alignment function in the CLC Genomics Workbench. The neighbor-joining algorithm and Jukes-Cantor nucleotide substitution model were implemented. All gaps were excluded from the analysis, and branch support was ascertained using 1,000 bootstrap replicates. The numbers next to the branches indicate the bootstrap values.
Phylogenetic analysis of Candida blankii and Debaromyces hansenii based on geographical distribution. Multiple alignment of ITS genes was performed using the Create Alignment function of the QIAGEN CLC Genomics Workbench (version 24.0.2). The neighbor-joining algorithm and Jukes-Cantor nucleotide substitution model were implemented. All gaps were excluded from the analysis, and branch support was ascertained using 1,000 bootstrap replicates. The numbers next to the branches indicate the bootstrap values. Phylogenetic analyses of C. blankii (A) and D. hansenii (B) are shown.
The antifungal susceptibility testing (AFST) data available for certain rare yeasts were analyzed retrospectively (Table S4). The resistance pattern of rare yeasts varied from low to high MIC for azoles, echinocandins, amphotericin B, and 5-fluorocytosine (Table S4). Although no breakpoints are available for rare yeasts, several of them, including Candida allociferrii, C. blankii, Candida oleophila, Candida subhashii, Kluyveromyces marxianus, Nakaseomyces bracarensis, and Wickerhamiella pararugosa, showed high MIC against first-line drug fluconazole (MIC range 16 to >256 µg/mL), Apiotrichum myxotoxinivorans, D. hansenii, and Trichosporon asahii showed high MIC against amphotericin B (MIC range 16 to >32 µg/mL), and Apiotrichum myxotoxinivorans, C. blankii, Candida lactativora, Candida steatolytica, Cutaneotrichosporon spp., and Naganishia spp. showed high MIC against echinocandins (MIC range 4 to >16 µg/mL) (Table S4).
Additionally, we also calculated MIC_50_ and MIC_90_ for yeast species with 10 or more isolates, and the majority of these species showed elevated MIC against fluconazole and few against voriconazole or amphotericin B, the drugs often used as the first choice of antifungals in the empirical antifungal therapy (Table S5). These results are not surprising as several of these rare yeasts have shown high MIC against azoles, echinocandins, and amphotericin B (6, 7, 29, 30). Of particular interest, C. blankii, which was believed to be non-pathogenic to humans until 2015, was reported as the cause of infection in 12 cases worldwide, and nine of them came from hospital-associated outbreaks in India (21, 54). In another case of C. blankii bloodstream infection in New York, the second C. blankii case in the United States, an unidentified yeast and Aspergillus niger were isolated from respiratory secretions. Voriconazole was administered as an empiric drug for both A. niger and the unidentified yeast. Unfortunately, the patient died within a week of isolating the yeast. Eventually, C. blankii was identified as the causative agent of fungemia in our laboratory by ITS-PCR and sequencing, and AFST revealed azole resistance (39). Such cases reinforce the significance of rapid and accurate identification of rare yeasts.
While commercial databases, such as Bruker or Vitek MS undergo frequent updates, there is a need for in-house library enrichment to include isolates of adequate worldwide representation. In the present investigation, MALDI library enrichment reduced the turnaround time of rare yeasts identification without the need to perform ITS-PCR and sequencing, which is time-consuming and labor-intensive. Taverna et al. developed an in-house MALDI database by adding regional strains and demonstrating improved MALDI scores (13). Similarly, studies all over the world have shown successful addition of regional isolates of Malassezia spp., C. auris, Candida haemulonii, Candida duobushaemulonii, Candida krusei, and Kodamaea ohmeri to their in-house MALDI databases (23, 24, 55). As a public health laboratory of New York State and the Centers for Disease Control and Prevention’s Northeast Regional Laboratory for the Antibiotic Resistance Laboratory Network of the United States, we urgently need to have robust identification technologies, including MALDI-TOF MS. Implementing an in-house Bruker library will enable the correct identification of an increasing number of rare yeasts and help in administering proper treatment options while controlling spread to the vulnerable population. The enriched library spectrum profiles generated in this study, comprising local isolates of rare yeasts from the northeastern United States, are now available publicly through the Centers for Disease Control and Prevention’s MicrobeNet database (https://microbenet.cdc.gov/).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pfaller MA, Diekema DJ. 2010. Epidemiology of invasive mycoses in North America. Crit Rev Microbiol 36:1–53. doi:10.3109/1040841090324144420088682 · doi ↗ · pubmed ↗
- 2Rodrigues ML, Nosanchuk JD. 2020. Fungal diseases as neglected pathogens: a wake-up call to public health officials. P Lo S Negl Trop Dis 14:e 0007964. doi:10.1371/journal.pntd.000796432078635 PMC 7032689 · doi ↗ · pubmed ↗
- 3Formanek PE, Dilling DF. 2019. Advances in the diagnosis and management of invasive fungal disease. Chest 156:834–842. doi:10.1016/j.chest.2019.06.03231351046 · doi ↗ · pubmed ↗
- 4Nagy F, Bozó A, Tóth Z, Daróczi L, Majoros L, Kovács R. 2018. In vitro antifungal susceptibility patterns of planktonic and sessile Candida kefyr clinical isolates. Med Mycol 56:493–500. doi:10.1093/mmy/myx 06228992253 · doi ↗ · pubmed ↗
- 5Sugita T, Takeo K, Ohkusu M, Virtudazo E, Takashima M, Asako E, Ohshima F, Harada S, Yanaka C, Nishikawa A, Majoros L, Sipiczki M. 2004. Fluconazole-resistant pathogens Candida inconspicua and C. norvegensis: DNA sequence diversity of the r RNA intergenic spacer region, antifungal drug susceptibility, and extracellular enzyme production. Microbiol Immunol 48:761–766. doi:10.1111/j.1348-0421.2004.tb 03602.x 15502409 · doi ↗ · pubmed ↗
- 6Kumar S, Kumar A, Roudbary M, Mohammadi R, Černáková L, Rodrigues CF. 2022. Overview on the infections related to rare Candida species. Pathogens 11:963. doi:10.3390/pathogens 1109096336145394 PMC 9505029 · doi ↗ · pubmed ↗
- 7Sharma M, Chakrabarti A. 2023. Candidiasis and other emerging yeasts. Curr Fungal Infect Rep 17:15–24. doi:10.1007/s 12281-023-00455-336741271 PMC 9886541 · doi ↗ · pubmed ↗
- 8Aydin M, Kustimur S, Kalkanci A, Duran T. 2019. Identification of medically important yeasts by sequence analysis of the internal transcribed spacer and D 1/D 2 region of the large ribosomal subunit. Rev Iberoam Micol 36:129–138. doi:10.1016/j.riam.2019.05.00231690527 · doi ↗ · pubmed ↗