Phylogenetic diversification and fitness trade-offs of TetA variants in mediating eravacycline resistance in Klebsiella pneumoniae
Meimei Fan, Zhixun Zhang, Xiaowei Liu, Liqin Deng, Yijie Lei, Feng Chen, Yuanyuan Ying, Chengfeng Fan, Jiaxin Gao, Zheer Ren, Jiayi Gu, Yuying Xia, Yifan Yuan, Keyu Zhang, Junfei Zhu, Yilin Yang, Weining Sun, Kaiying Cheng, Alessandra Carattoli, Christoph M. Tang, Zhen Shen

TL;DR
This study explores how different TetA variants in Klebsiella pneumoniae affect antibiotic resistance and bacterial fitness, revealing evolutionary trade-offs that limit their spread.
Contribution
The study systematically characterizes TetA variants in K. pneumoniae, linking their resistance to eravacycline with fitness costs and ecological trade-offs.
Findings
Four TetA variants confer eravacycline resistance with variable tigecycline susceptibility.
Resistance variants impose a significant fitness cost, reducing tolerance to H2O2 and cadmium.
Limited dissemination of these variants suggests evolutionary trade-offs constrain their spread.
Abstract
The role of TetA variants in mediating tigecycline and eravacycline resistance in Klebsiella pneumoniae remains a critical area of investigation. However, there has been a lack of systematic characterization of the epidemiology, resistance phenotypes, and fitness costs of TetA variants. Here, we identified 28 TetA variants in K. pneumoniae from the National Center for Biotechnology Information database from 824 isolates, categorizing them into three phylogenetically distinct clades. Among these, four variants were shown to mediate eravacycline resistance, with concurrent but variable effects on tigecycline susceptibility. Notably, these resistance-conferring variants exhibited limited dissemination across clinical and environmental strains. Analyses revealed that their expression imposes a significant fitness cost, markedly reducing bacterial tolerance to the clinical disinfectant H2O2…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
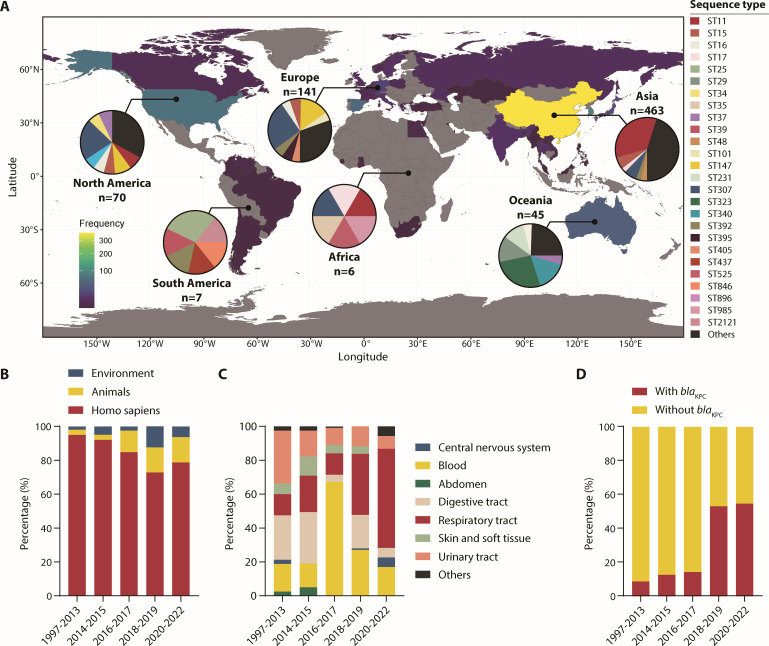 Fig 1
Fig 1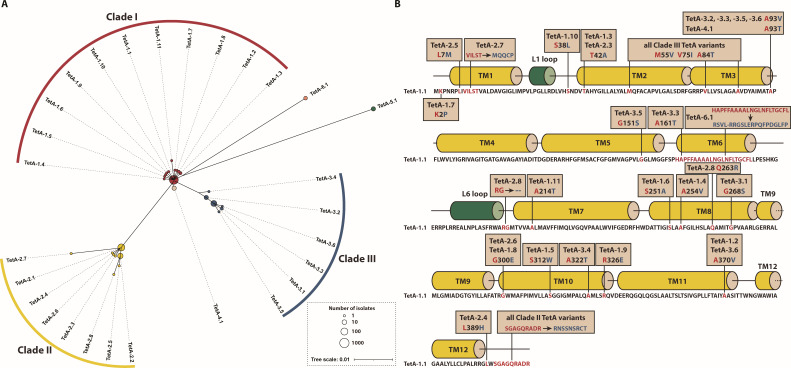 Fig 2
Fig 2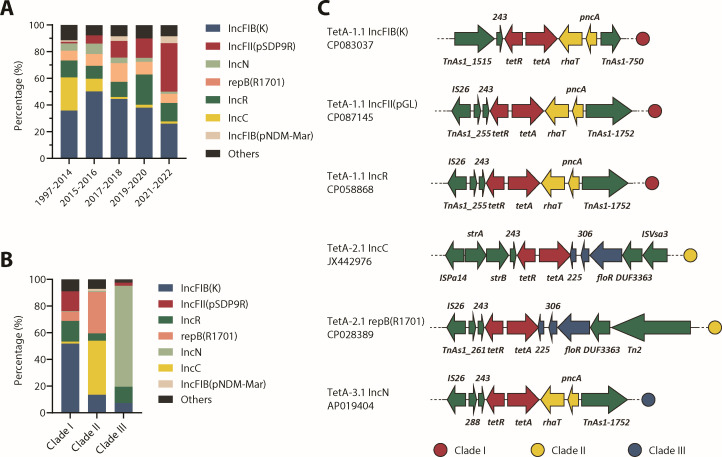 Fig 3
Fig 3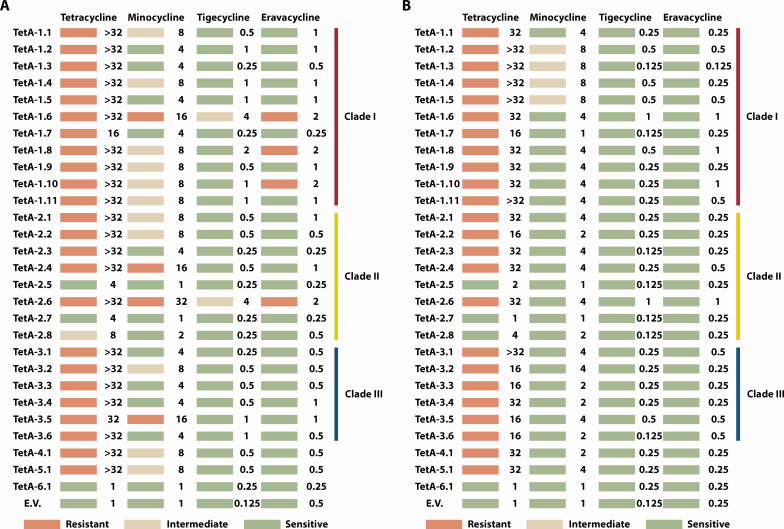 Fig 4
Fig 4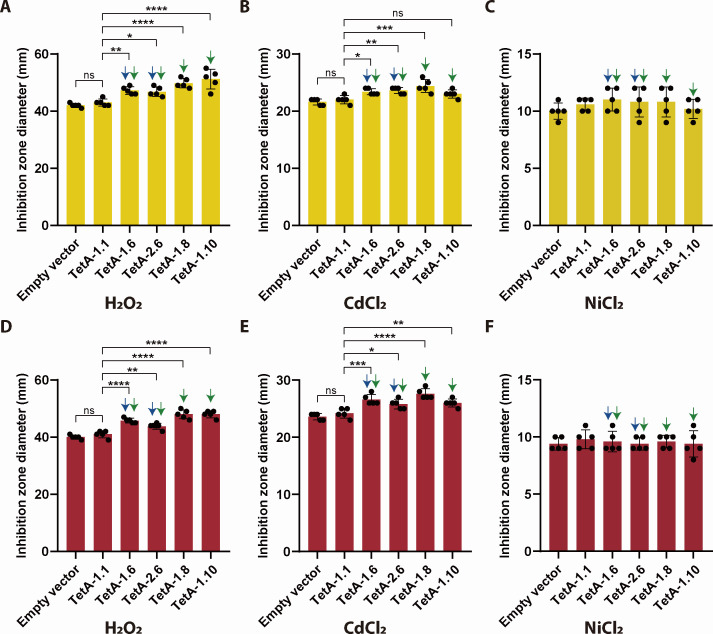 Fig 5
Fig 5- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Natural Science Foundation of Zhejiang Province
- —Hangzhou Youth Innovation Team Project
- —Scientific Research Project of the Zhejiang Provincial Education Department
- —Scientific Research Project of the Zhejiang Provincial Education Department
- —HZNU scientific research and innovation team project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Evolution and Genetic Dynamics
INTRODUCTION
Carbapenem-resistant Klebsiella pneumoniae (CRKP) poses a critical global health threat due to its propensity for multidrug resistance and nosocomial transmission (1, 2). While advanced tetracyclines such as tigecycline and eravacycline serve as last-line therapies against CRKP (3, 4), their escalating clinical use has precipitated a concerning rise in resistance (5–8).
Mechanistically, resistance to advanced tetracyclines primarily arises from enzymatic inactivation by TetX-family oxidases or drug extrusion via efflux pumps such as TMexCD-TOprJ, OqxAB, and MacAB (9–12). Notably, these determinants are frequently environmental or zoonotic in origin and remain underrepresented in clinical isolates, suggesting niche-specific barriers to their human adaptation (13, 14). In stark contrast, variants of TetA—a major facilitator superfamily (MFS) efflux pump—have emerged as predominant clinical drivers of tigecycline resistance in K. pneumoniae (15–17). Despite their clinical relevance, the evolutionary trajectories and ecological constraints governing TetA variant dissemination across the One Health continuum remain enigmatic.
Here, we bridge this gap through a systematic phylogenetic and functional dissection of TetA variants in K. pneumoniae. By mining National Center for Biotechnology Information (NCBI) genomes, we identified 28 TetA variants, 4 of which conferred eravacycline resistance, a previously unreported phenotype for this efflux pump family. Surprisingly, molecular epidemiology revealed limited spread of these variants with potential risk (VWPRs), hinting at uncharacterized fitness costs offsetting their resistance advantage.
Guided by the One Health paradigm, we hypothesized that the environmental stressors commonly present in hospital and ecological settings might impose selective barriers limiting the spread of VWPRs (18, 19). Consistent with this, K. pneumoniae expressing the TetA VWPRs exhibited hypersusceptibility to H_2_O_2_ (a common disinfectant) and cadmium. These findings implicate environmental heavy metals as not merely passive contaminants but also as key players modulating the dissemination of resistance genes—a dynamic overlooked in current surveillance frameworks.
MATERIALS AND METHODS
Epidemiological and phylogenetic analysis of tetA variants
The tet(A)v1 allele (GenBank Accession number X61367) was used as a query in BLASTn searches against the NCBI K. pneumoniae genome database (cutoff: ≥90% identity). Metadata (isolation source, geographic origin, and collection year) were curated for 28 non-redundant tetA homologs (Table S1). Amino acid sequences of TetA variants (Table S2) were aligned with Clustal Omega (20), visualized via iTOL (https://itol.embl.de). Variants were divided into clades based on phylogeny, and within each clade, variants were designated based on allele frequency. Transmembrane domain prediction was based on TetA-1.1 structure predicted by AlphaFold3 (21).
Bacterial strains and culture conditions
All bacterial strains used in this study are listed in Table S3. Cloning host: Escherichia coli DH5α was cultivated in Lysogeny Broth (LB) medium at 37°C with agitation (150 rpm). K. pneumoniae strains—including clinical isolate Kp36 (22), TU37, HS11286 (23), and their derivatives—were grown under identical conditions. Antibiotics were supplemented as follows: carbenicillin (100 mg/L) for plasmid selection and kanamycin (50 mg/L) for mutant screening.
Plasmid construction and molecular cloning
The tetracycline resistance gene tetA-1.1 was originally from K. pneumoniae Kp36, cloned into a low-copy vector pUA139 (modified from pUA139 ada) (24). Gene fragments and vector backbones were amplified by PCR (Phanta Flash DNA Polymerase, Vazyme), assembled in vitro using Gibson Assembly Master Mix (ABclonal #RK21022), and transformed into E. coli DH5α via heat shock. Plasmid integrity was verified by Sanger sequencing (Tsingke Biotech), and validated constructs were purified (QIAprep Spin Miniprep Kit) for downstream applications. All plasmids and primers used in this study are listed in Tables S4 and S5, respectively.
Antimicrobial susceptibility profiling
Minimum inhibitory concentrations (MICs) for tigecycline and eravacycline were determined by broth microdilution (Clinical and Laboratory Standard Institute M07-A11) (25) in cation-adjusted Mueller-Hinton broth. K. pneumoniae TU37 harboring tetA variants was tested in triplicate. Susceptibility interpretations followed the US Food and Drug Administration (https://www.fda.gov/drugs/development-resources/tigecycline-injection-products) (tigecycline: susceptible ≤2 mg/L, intermediate 4 mg/L, and resistant ≥8 mg/L) and China ECAST (eravacycline: susceptible ≤1 mg/L) criteria (26).
Growth assays
Bacterial growth was measured in a real-time microbial growth curve system (Viewkr Biotechnology). Bacteria diluted to 6 × 10³ CFU/mL and the negative control (phosphate-buffered saline) were transferred to a 96-well plate at T0, which was then grown in LB, acidic LB (pH = 5.0), or M9 minimal media supplemented with anhydrotetracycline (aTC) (50 μg/L) as an inducer at 37°C. Bacterial growth data were acquired every 10 min for 35 h (n = 3). Before each measurement, the 96-well plate was agitated for 15 s at 30 Hz.
Heavy metals and disinfectants susceptibility assays
In the initial screen, tetracycline (10 mg/L, 30 min)-induced tetA-expressing strains (HS11286 and TU37) were adjusted to OD_600_ = 0.1, plated on LB agar with tetracycline (10 mg/L), and exposed to 6-mm filter discs impregnated with heavy metals (K_2_TeO_3_ [0.1 M], CuSO_4_ [0.1 M], AgNO_3_ [0.1 M], CdCl_2_ [0.1 M], and NiCl_2_ [0.01 M]) and disinfectants (benzalkonium chloride [neat], sodium hypochlorite [10% dilution], and H_2_O_2_ [3% vol/vol]). Inhibition zones were measured after 16-h incubation at 37°C (n = 3). Disinfectants and heavy metals exhibit differential inhibitory effects on TetA VWPRs compared to TetA-1.1 and were tested again following the same protocol except tetracycline was replaced by aTC (50 μg/L) as the inducer.
Time-kill assays
Overnight cultures of K. pneumoniae expressing TetA variants grown in LB supplemented with aTC (50 μg/L) and kanamycin (50 mg/L) were diluted and subcultured into 20 mL of LB supplemented with aTC (50 μg/L) and kanamycin (50 mg/L) with initial OD_600_ of 0.008, grown at 37°C with shaking (220 rpm) for 3 h before H_2_O_2_ or CdCl_2_ was added. Viable bacteria were measured by serial dilution and plating onto LB agar at 0, 2, 4, and 6 h post-addition of H_2_O_2_ or CdCl_2_. Final concentration of H_2_O_2_ used for killing is 0.012% (vol/vol) for both K. pneumoniae TU37 and HS11286. Final concentration of CdCl_2_ used for killing is 40 mM for K. pneumoniae TU37 and 2.5 mM for HS11286.
Statistical analysis
Data were analyzed in GraphPad Prism 9.0. One-way or two-way analysis of variance (Tukey’s post hoc) was used to assess differences between groups in the disc diffusion susceptibility assays or time-kill assays, respectively. Significance thresholds: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
RESULTS
Global dissemination and clinical shifts in TPKP isolates
A systematic analysis of 824 tetA-positive K. pneumoniae (TPKP) genomes isolated from 1997 to 2022 revealed their distribution across 45 countries, with Asia harboring the highest prevalence (63.5%) and Africa the lowest (1.0%) (Fig. 1A). TPKPs exhibited striking genetic diversity, spanning 135 sequence types (STs), though ST11 dominated among Asian isolates (30.1%), while ST307 prevailed in Europe (22.9%) and North America (20.5%). Notably, while clinical sources accounted for >70% of isolates, environmental and animal-derived TPKPs surged post-2015 (7.0% to 28.1%), implicating non-human reservoirs as sources for emerging tetA dissemination (Fig. 1B). Clinically, TPKP infection sites shifted markedly over time: respiratory tract infections rose from 14.6% (1997–2013) to 55.4% (2020–2022), whereas urinary tract cases declined from 30.5% to 7.1% (Fig. 1C). Strikingly, the rate of clinical TPKPs co-harboring blaKPC (KPC-TPKP) increased over fivefold from 9.7% (1997–2013) to 55.4% (2020–2022), with ST11 strains alone accounting for 58.5% (72/123) of all KPC-TPKP isolates. These findings suggest that tetA might have provided blaKPC-positive K. pneumoniae (KPC-KP) with selective advantages resisting tetracycline antibiotics, offering a potential explanation for the success of the highly transmissible ST11 lineage.
824 TPKPs from 1997 to 2022 were analyzed in this study, including their geographical distribution and STs (A), isolation sources (B), and infection sites (C). (D) Proportion of clinical TPKPs with or without blaKPC. STs of TPKPs were obtained by MLST and the map was generated with R and the ggplot2 package.
Phylogenetic diversification and structural hotspots of TetA variants
Classification of 28 TetA variants by prevalence and phylogeny resolved three Clades (I–III) with distinct evolutionary origins (Fig. 2A; Table S2), each of which was represented by an ancestral variant TetA-1.1, TetA-2.1, and TetA-3.1, respectively. The defining mutation that separates Clade II from Clade I is a C-terminal nine-amino acid substitution (SGAGQRADR→RNSSNSRCT), while the key mutation for distinguishing Clade III from Clade I is M55V/V75I/A84T/A93V/G268A. Strikingly, TetA-5.1—the most divergent variant—was found to be most frequently associated with Enterobacter hormaechei by BLASTn analysis (Fig. S1), implicating inter-genus horizontal transfer. Structural prediction of TetA-1.1 by AlphaFold3 localized mutation hotspots to transmembrane domains TM3, TM8, and TM10 (Fig. 2B), suggesting that these regions are critical for substrate recognition and efflux efficiency.
Phylogenetic tree (A) and the mutations of each TetA variant mapped onto their predicted secondary structure (B).
Plasmid dynamics drive clade-specific TetA spread
To gain insights into the horizontal transfer of TetA variants, we analyzed the genetic environments of tetA variants. TetA variants predominantly resided on IncFIB(K), IncFII(pSDP9R), and IncN plasmids: IncFII(pSDP9R) surged by 30-fold from 1.2% (1997–2014) to 36.2% (2021–2022), displacing IncC plasmids, which dropped from 24.8% to 1.7% in the same period (Fig. 3A). Clade-specific plasmid associations emerged: Clade I favored IncFIB(K) plus IncR plasmids (together 67.5%), Clade II IncC plus repB(R1701) (together 71.4%), and Clade III IncN (75.6%) (Fig. 3B). Genetic context analysis revealed conserved downstream elements: pncA (putative nicotinamidase-related amidase) and rhaT (putative drug/metabolite transporter superfamily permease) in Clades I/III vs floR (chloramphenicol efflux pump) and a 225-bp hypothetical gene-hp225 (Fig. 3C). Notably, hp225 has a 27-bp overlap with tetA in Clade II plasmids, responsible for Clade II’s defining C-terminal substitution, highlighting plasmid architecture as a driver of TetA diversification.
Shifting trends of replicon distribution of tetA plasmids over 1997–2022 (A). Replicon distribution (B) and the neighboring genes (C) of tetA representative of Clades I–III.
TetA VWPRs: resistance gains and ecological costs
To investigate the ability of the 28 TetA variants to mediate resistance to tetracycline family antibiotics, we measured the MICs of tetracycline family antibiotics after expressing tetA variants with tetR from their native genetic background on a low-copy plasmid in K. pneumoniae TU37 (ST23) and E. coli MG1655 (Fig. 4). Resistance phenotypes conferred by TetA variants decreased with the generational advancement of tetracycline antibiotics (Fig. 4). 85.7% of the variants confer resistance to tetracycline in both species, with TetA-2.5, TetA-2.7, TetA-2.8, and TetA-6.1 having MICs indistinguishable from the empty plasmid control, indicating that these variants might have lost their efflux ability due to their mutations. In contrast, four variants (TetA-1.6, TetA-2.4, TetA-2.6, and TetA-3.5) are capable of mediating resistance to the second-generation minocycline in K. pneumoniae. Notably, even to the third-generation tigecycline, TetA-1.1 led to fourfold higher MIC in K. pneumoniae. However, none of the 28 TetA variants mediated resistance above the clinical breakpoint (≥4 mg/L), with only TetA-1.6 and TetA-2.6 showing intermediate resistance in K. pneumoniae. Surprisingly, more variants (TetA-1.6, TetA-2.6, TetA-1.8, and TetA-1.10) conferred resistance against the fourth-generation eravacycline in K. pneumoniae, a novel phenotype for TetA. Based on the resistance phenotype to tigecycline and eravacycline, TetA-1.6, TetA-1.8, TetA-1.10, and TetA-2.6 were classified as VWPRs.
Resistance phenotypes and MICs of K. pneumoniae TU37 (A) and E. coli MG1655 (B) toward four generations of tetracycline antibiotics upon expression of tetA variants on a low-copy plasmid. Breakpoints for tigecycline: susceptible ≤2 mg/L, intermediate 4 mg/L, resistant ≥8 mg/L and for eravacycline: susceptible ≤1 mg/L, resistant >1 mg/L. E.V.: empty vector control.
Paradoxically, despite conferring advanced tetracycline resistance, these VWPRs showed minimal epidemiological spread, with each variant only reported in patients once. This epidemiological paradox prompted us to investigate potential fitness costs through growth kinetic assays under nutrient-deprived (M9 minimal media) and weakly acidic (pH = 5.0 LB) conditions in two K. pneumoniae strains: TU37 (ST23) and HS11286 (ST11 CRKP). To our surprise, in HS11286, TetA-1.8 and TetA-1.10 conferred a significant growth advantage during exponential phase (5–12 hours) compared to TetA-1.1 in both LB and M9 media (Fig. S2A and C). Conversely, in TU37, TetA-1.6 expression caused a minor (≈5%) reduction in stationary-phase optical density in both normal and acidic LB, while showing no significant fitness defects in M9 (Fig. S2D and E).
The observation that VWPRs exhibited minor growth defects despite clinical scarcity motivated us to systematically evaluate their collateral sensitivity to environmental stressors. We selected a panel of hospital-relevant disinfectants including H_2_O_2_, disinfectant “84” (main ingredient: HClO), and quaternary ammonium compounds alongside environmental heavy metals (CdCl_2_, NiCl_2_, K_2_TeO_3_, and CuSO_4_) using disc diffusion assays. To bypass TetR-mediated repression, tetracycline was included as an inducer in the initial screen. Notably, expression of TetA-1.6 conferred hypersusceptibility to NiCl_2_ and CdCl_2_ than TetA-1.1 in both K. pneumoniae strains, and expression of TetA-1.8/1.10 resulted in higher sensitivity to H_2_O_2_ (Fig. S3A and D), while VWPRs showed equivalent susceptibility to other disinfectants and heavy metals tested compared to TetA-1.1 (Fig. S4). To rule out the potential confounding effects of tetracycline’s antimicrobial activity, we repeated assays with H_2_O_2_, CdCl_2_, and NiCl_2_ using aTC—a tetracycline analog with minimal antibacterial activity—as the inducer. Indeed, both K. pneumoniae strains displayed significantly lower sensitivity to NiCl_2_ (>30%) and CdCl_2_ (>20%), while differences in susceptibility of NiCl_2_ between variants were no longer observed (Fig. 5B, C, E and F), suggesting that tetracycline may synergistically enhance metal toxicity. Surprisingly, this aTC-based assay exacerbated H_2_O_2_ and CdCl_2_ toxicity in VWPR-expressing strains (Fig. 5A, B, D and E).
*Inhibition zone diameters of K. pneumoniae TU37 (A–C) and HS11286 (D–F) expressing TetA variants to H2O2, CdCl2, and NiCl2. Significance thresholds: *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
As disinfectants could also be applied to already established bacterial reservoirs in various clinical or environmental scenarios, we sought to test if the expression of VWPRs modulates the tolerance of K. pneumoniae to H_2_O_2_ and CdCl_2_ by conducting time-kill assays. Notably, expression of TetA variants led to a significant reduction of tolerance to both H_2_O_2_ and CdCl_2_ compared to the empty vector control (Fig. S5A through C). While the increased tolerance of TetA-1.1-expressing strains compared to VWPR-expressing strains did not reach statistical significance (P = 0.08–0.14 at 2 h post H_2_O_2_ treatment) due to time-kill assay variability, a consistent numerical trend was observed for both stressors (Fig. S5A through D). Overall, our findings underscore ecological trade-offs that constrain VWPRs to transient clinical niches rather than environmental persistence.
DISCUSSION
Since the initial linkage of tetA to tigecycline resistance in K. pneumoniae (17), a few tetA variants have been shown to mediate tigecycline resistance in K. pneumoniae (27, 28). Moreover, an epidemiological description of tetA in K. pneumoniae is still lacking. Our global analysis revealed that the association of tetA with multidrug-resistant high-risk clones—ST11 in Asia (29), ST307/ST147 in Europe/North America (30), and ST323 in Australia (31)—has become stronger, placing tetA as a novel genomic hallmark of these epidemic lineages, raising concerns that TetA-mediated resistance to advanced tetracyclines (e.g., eravacycline) could undermine last-line therapies. Unlike other clinically important resistance genes such as blaKPC and mcr, whose variants and resistance phenotypes have been well characterized (32, 33), TetA variants remain systematically underexplored. To fill this gap, we searched the NCBI database and obtained 824 TPKPs from 1997 to 2022, from which we identified 28 variants and classified them into 3 major clades. TetA-1.1 was initially reported as tet(A) type I variant (Tet(A)v1) (17). Thus, we believe it is more appropriate to designate tetA-1.1 as the “wild-type” tetA allele in K. pneumoniae due to its prevalence (73.7%). Moreover, tetA variants exhibit clade-specific plasmid associations. Given the crucial roles played by plasmids in resistance transmission (34, 35), our findings enable real-time tracking of variant evolution, mechanistic dissections of the tetA transmission dynamics, and the potential design of novel plasmid-centric interventions.
Eravacycline, a synthetic fluorocycline, is a fourth-generation tetracycline (36). Since its approval by the FDA in 2018, reports of eravacycline resistance in K. pneumoniae have remained scarce. The identification of TetA-1.6/1.8/1.10/2.6 as eravacycline resistance determinants expands the known mechanisms of fluorocycline resistance beyond TetX enzymes and efflux pumps such as OqxAB (8, 12), highlighting the underappreciated role of TetA variants in shaping resistance. Strikingly, both TetA-1.6 (S251A) and TetA-1.8 (G300E) emerged during tigecycline therapy (16, 37, 38), suggesting that sequential exposure to antibiotics selects for “pan-tetracycline” MFS transporters. In contrast, TetA-2.6 (G300E + C-terminal SGAGQRADR to RNSSNSRCT) and TetA-1.10 (S38L) were both recovered from patients (39) and emerged in the absence of tigecycline treatment, hinting at the presence of stealthy evolutionary pathway without direct selection. Notably, TetA-2.6 exhibited higher resistance to minocycline and tigecycline than TetA-1.8, suggesting that functional divergence served as the basis for clade formation. The identification of these TetA variants not only expands the mechanistic repertoire of eravacycline resistance but also underscores TetA as an evolutionarily adaptable and clinically underappreciated resistance determinant, demanding prioritized surveillance.
K. pneumoniae is known to be a key trafficker of AMR genes from environmental microbes to clinically important pathogens (19, 40). Here, this is exemplified by the spread of TetA to 135 distinct STs and its continued isolation from animals and environmental sources (41, 42). Despite their clinical emergence, VWPRs so far have demonstrated restricted dissemination, a paradox reflected by ecological trade-offs. In this study, strains expressing all four VWPRs exhibited heightened susceptibility to H_2_O_2_ (which is ubiquitous in healthcare settings) and the heavy metal cadmium. We employed two complementary methodologies to assess resistance and tolerance, each with distinct advantages and limitations. First, the disc diffusion assay, a well-established and reproducible technique for resistance profiling, excels in detecting subtle differences between strains as it generates a continuous gradient of stressors tested. However, this method remains largely qualitative. Second, the time-kill assay provides quantitative insights into tolerance but exhibits inherent variability in kinetic measurements, which may affect reproducibility.
Our findings are consistent with recent evidence that TetA-1.1 imposes fitness costs for growth (37), with the concept now extended to trade-offs for susceptibility to environmental factors. Heavy metals exert significant selective pressures on bacteria in the environment, which is evidenced by the ubiquitous carriage of heavy metal resistance genes (HMRGs) by multidrug-resistant Enterobacteriaceae, such as K. pneumoniae (43, 44). Thus, heavy metals enriched in agricultural/industrial habitats by anthropogenic activities (45) likely purge TetA VWPRs from environmental reservoirs, restricting their success. Thus, while clinical antibiotic pressure selects for TetA diversification, environmental metal stress counter-selects against the spread of VWPRs—a delicate equilibrium exploitable for intervention.
In summary, our findings suggest that dissemination of resistance could be viewed as an ecological tug-of-war, in which the stability of VWPRs is kept in a delicate equilibrium; the benefits of antibiotic resistance are offset by collateral sensitivity to other agents. This such informs approaches for a sustainable One Health strategy to mitigate dissemination of resistance to advanced tetracyclines, which integrates four different dimensions: (i) antibiotic stewardship to reduce eravacycline/tigecycline selection, (ii) application of H_2_O_2_-based disinfectants and monitor resistant mutants, (iii) sampling of heavy metal polluted hotspots to watch for acquisition events of HMRGs by TPKPs, and (iv) plasmid-centric surveillance prioritizing the major replicons.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ikuta KS, Swetschinski LR, Robles Aguilar G, Sharara F, Mestrovic T, Gray AP, Davis Weaver N, Wool EE, Han C, Gershberg Hayoon A, et al.. 2022. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. The Lancet 400:2221–2248. doi:10.1016/S 0140-6736(22)02185-7 · doi ↗
- 2Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, Pulcini C, Kahlmeter G, Kluytmans J, Carmeli Y, Ouellette M, Outterson K, Patel J, Cavaleri M, Cox EM, Houchens CR, Grayson ML, Hansen P, Singh N, Theuretzbacher U, Magrini N, WHO Pathogens Priority List Working Group. 2018. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 18:318–327. doi:10.1016/S 1473-3099(17)30753-329276051 · doi ↗ · pubmed ↗
- 3Agyeman AA, Bergen PJ, Rao GG, Nation RL, Landersdorfer CB. 2020. A systematic review and meta-analysis of treatment outcomes following antibiotic therapy among patients with carbapenem-resistant Klebsiella pneumoniae infections. Int J Antimicrob Agents 55:105833. doi:10.1016/j.ijantimicag.2019.10.01431730892 · doi ↗ · pubmed ↗
- 4Blanchard LS, Armstrong TP, Kresken M, Emery CL, Ying YX, Sauvonnet V, Zambardi G. 2023. Multicenter clinical evaluation of ETEST eravacycline for susceptibility testing of Enterobacteriaceae and enterococci. J Clin Microbiol 61:e 0165022. doi:10.1128/jcm.01650-2236877080 PMC 10035295 · doi ↗ · pubmed ↗
- 5Liao W, Liu Y, Zhang W. 2020. Virulence evolution, molecular mechanisms of resistance and prevalence of ST 11 carbapenem-resistant Klebsiella pneumoniae in China: a review over the last 10 years. J Glob Antimicrob Resist 23:174–180. doi:10.1016/j.jgar.2020.09.00432971292 · doi ↗ · pubmed ↗
- 6Zhang Y, Liu D, Liu Y, Li Q, Liu H, Zhou P, Liu Y, Chen L, Yin W, Lu Y. 2024. Detection and characterization of eravacycline heteroresistance in clinical bacterial isolates. Front Microbiol 15:1332458. doi:10.3389/fmicb.2024.133245838601926 PMC 11004243 · doi ↗ · pubmed ↗
- 7Seifert H, Blondeau J, Dowzicky MJ. 2018. In vitro activity of tigecycline and comparators (2014-2016) among key WHO “priority pathogens” and longitudinal assessment (2004-2016) of antimicrobial resistance: a report from the T.E.S.T. study. Int J Antimicrob Agents 52:474–484. doi:10.1016/j.ijantimicag.2018.07.00330012439 · doi ↗ · pubmed ↗
- 8Liu X, Liu Y, Ma X, Chen R, Li C, Fu H, et al.. 2024. Emergence of plasmid-borne tet(X 4) resistance gene in clinical isolate of eravacycline and omadacycline. Microbiol Spectr 12. doi:10.1128/spectrum.00496-24 · doi ↗