fosZ, a novel plasmid-borne fosfomycin resistance gene in Pseudomonas species, especially carbapenem-resistant Pseudomonas aeruginosa isolates
Youxing Shao, Xin Lan, Xuefei Zhang, Leilei Wang, Fan Yang, Minggui Wang, Qinglan Guo

TL;DR
A new gene called fosZ, found in Pseudomonas bacteria, helps them resist the antibiotic fosfomycin and spreads through plasmids, especially in drug-resistant strains.
Contribution
Discovery of fosZ, a novel plasmid-borne fosfomycin resistance gene in Pseudomonas species.
Findings
fosZ reduces susceptibility to fosfomycin and its inhibitor PPF in Pseudomonas strains.
ISPa75-fosZ is found on plasmids and chromosomes, often linked to carbapenem resistance genes.
Structure analysis shows unique features in FosZ that affect drug binding.
Abstract
A novel fosfomycin-resistant glutathione S-transferase (FR-GST) gene, fosZ, was investigated, and its structural characteristics were characterized in Pseudomonas species. The fosZ gene was cloned and expressed in P. aeruginosa PAO1 and HS355, where it displayed reduced susceptibility to fosfomycin (8- to 64-fold) and its inhibitor sodium phosphonoformate (PPF). FosZ shares less than 70% amino acid identity with known FosA proteins. Bioinformatics analyzes revealed that fosZ was a transposable passenger gene within ISPa75, likely captured from Pseudomonas species. A total of 159 fosZ-bearing Pseudomonas strains were identified in GenBank over the past 22 years, sharing ten target site duplications (TSDs) associated with ISPa75. Among them, 34 strains were fully sequenced. ISPa75-fosZ was found on at least two chromosomes and 33 plasmids from four incompatibility groups, including the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
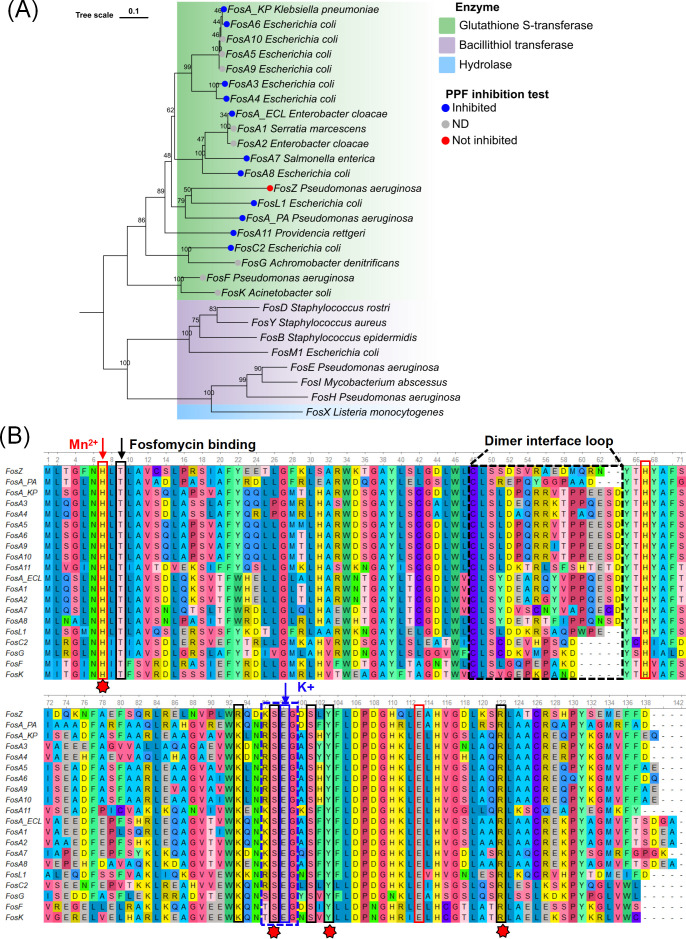 Fig 1
Fig 1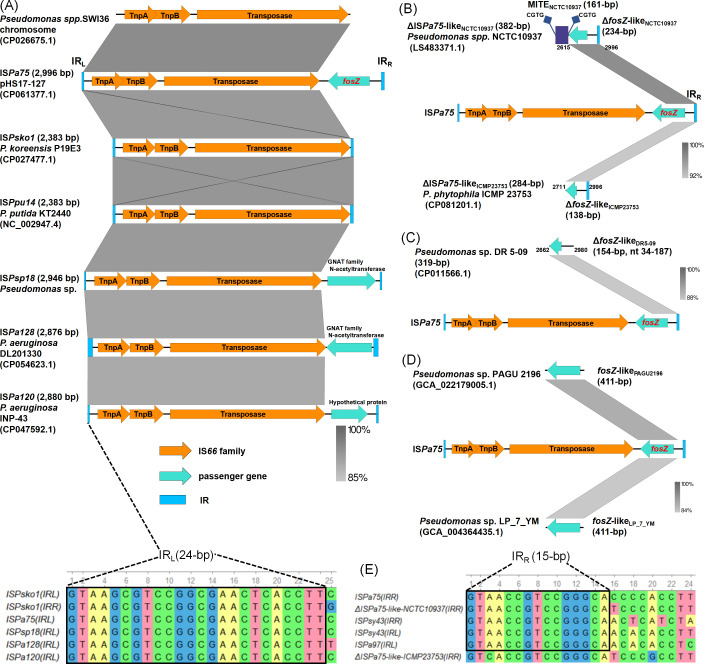 Fig 2
Fig 2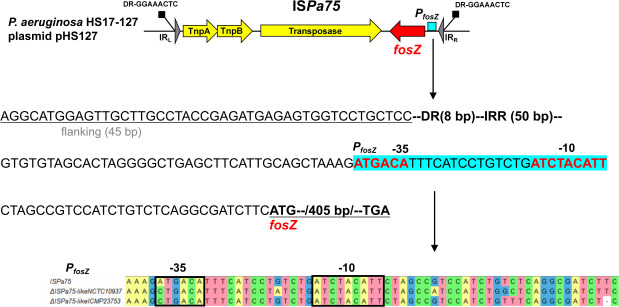 Fig 3
Fig 3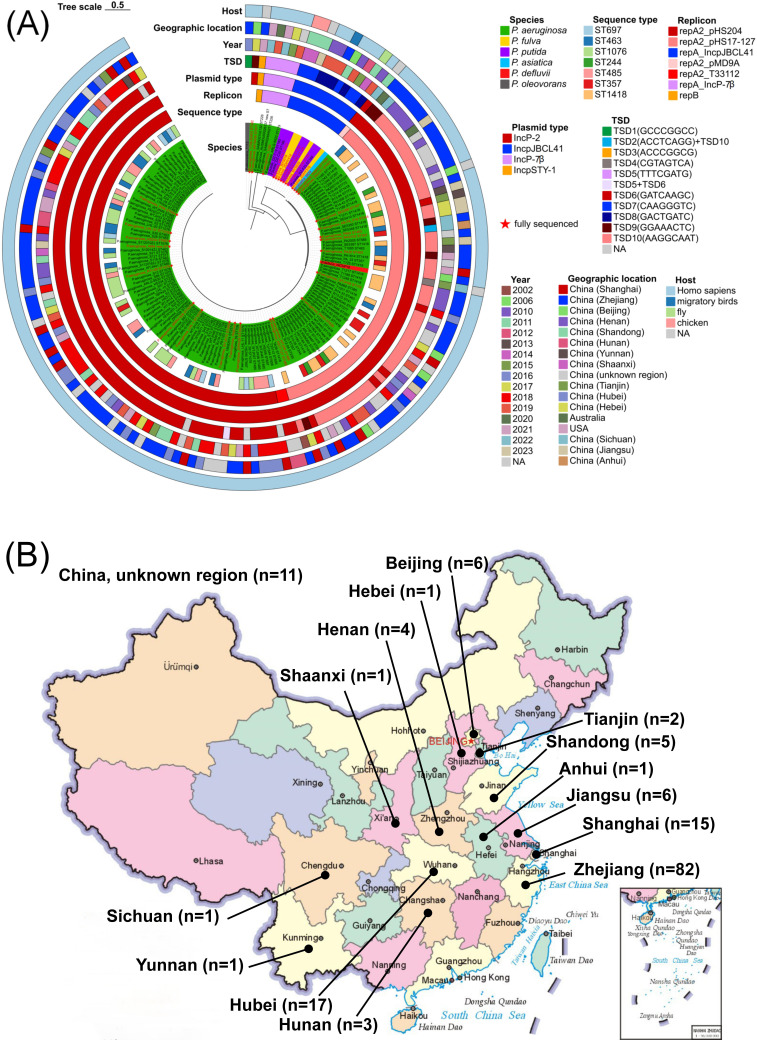 Fig 4
Fig 4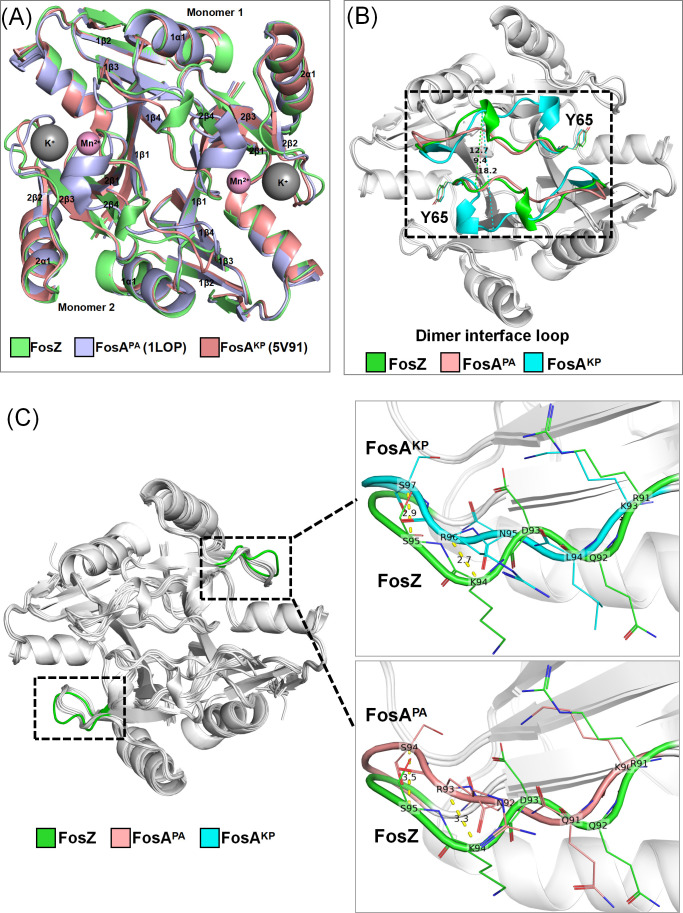 Fig 5
Fig 5| Isolate | Fosfomycin MIC (mg/L) | |||
|---|---|---|---|---|
| Without PPF | PPF | PPF | PPF | |
| 128 | 128 | 128 | 128 | |
| TcHS17-127 | 128 | 128 | 128 | 64 |
| 64 | 16 | 16 | 16 | |
| 64 | 16 | 16 | 16 | |
| 256 | 256 | 256 | 256 | |
| 512 | 512 | 512 | 512 | |
| 8 | 8 | 8 | 8 | |
| 512 | 512 | 512 | 512 | |
| 512 | 512 | 512 | 512 | |
| 1024 | 256 | 256 | 256 | |
| 0.5 | 1 | 1 | ND | |
| 0.5 | 1 | 1 | ND | |
| 2 | 2 | 4 | ND | |
- —National Key Research and Development Program of Chinahttp://dx.doi.org/10.13039/501100012166
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Glutathione Transferases and Polymorphisms · Cancer therapeutics and mechanisms
INTRODUCTION
Multidrug-resistant Pseudomonas aeruginosa (MDR-PA) poses a serious public health concern. The limited therapeutic options for combating MDR-PA and extensively drug-resistant P. aeruginosa (XDR-PA) have reignited interest in older, underutilized drugs like fosfomycin (1, 2). Fosfomycin has potent in vitro antibacterial activity against a broad spectrum, has led to its reemergence in combination with other antimicrobials for treating MDR-PA bacterial infections (3, 4). However, fosfomycin use encounters challenges from inherent and acquired resistance mechanisms. Fosfomycin resistance mechanisms in P. aeruginosa encompass (5) (i) the existence of recycling pathways bypassing de novo peptidoglycan biosynthesis, (ii) mutations in the fosfomycin transporter GlpT, and (iii) chromosomal fosfomycin-modifying enzymes, including FosA^PA^, FosE, FosF, and FosH.
Fosfomycin-modifying enzymes include three types of metalloenzymes (1, 2, 5): Mn^2+^- and K^+^-dependent glutathione S-transferase (e.g. FosA), Mg^2+^-dependent bacillithiol transferase (e.g., FosB), Mn^2+^-dependent hydrolase FosX, and fosfomycin kinases. Glutathione S-transferases, represented by FosA enzymes, are frequently found in chromosomes of Gram-negative species (6). Eleven variants of FosA (FosA1 to FosA11) have been characterized (7–12) and have been acquired by transposons and/or plasmids mostly in MDR Enterobacterales (Table S1). In P. aeruginosa, fosA homologs are present in nearly 99% of the 2,257 surveyed genomes (6). Several other acquired glutathione-S-transferase genes, like fosC2, fosK, and fosL1, have also been identified in Gram-negative species (13, 14).
Sodium phosphonoformate (PPF), approved for clinical use in treating cytomegalovirus infection and acyclovir-resistant herpes simplex virus infection, is a competitive inhibitor of glutathione S-transferase (15). PPF has been effective in detecting the production of glutathione S-transferase responsible for fosfomycin resistance (FR-GST), such as FosA3, FosA4, and FosC2 in Escherichia coli (16), and chromosomal fosA from multiple Gram-negative species, including P. aeruginosa (6).
In our prior studies, we reported a carbapenem-resistant P. aeruginosa (CRPA) isolate HS17-127 (17, 18), which co-carried the metallo-β-lactamase (MBL) genes blaAFM-1 and blaIMP-45 within an MDR transposon situated on a conjugative IncP-2 megaplasmid, pHS17-127. In this study, we identified a new FR-GST gene, named fosZ, located on pHS17-127, encoding FosZ enzyme, which is surprisingly not inhibited by PPF. This study represents the first exploration of plasmid-borne fosfomycin-modifying enzyme in P. aeruginosa.
MATERIALS AND METHODS
Bacterial isolates and antimicrobial susceptibility testing
The CRPA isolate HS17-127 and its transconjugant (18), along with the fosfomycin-susceptible P. aeruginosa isolate HS355 (19) and fosfomycin-resistant Enterobacter cloacae 19-4074 (*fosA3-*positive), were obtained in our previous study. Antimicrobial susceptibility testing for fosfomycin was performed using the agar dilution method, following 2024 Clinical and Laboratory Standards Institute (CLSI) recommendations (20). Glutathione S-transferase activity inhibition was examined using PPF, as previously described (6, 8).
Cloning experiments
The complete coding sequence of fosZ (411 bp) was cloned into Escherichia-Pseudomonas shuttle vector pUCP19 (Shanghai Bioresource Collection Center, China). To confirm the role of the promoter region in FosZ expression, which includes the full fosZ gene and a 300 bp upstream region with its native promoter, was also cloned into pUCP19. The recombinant plasmids were transformed into P. aeruginosa PAO1 and HS355, as well as E. coli DH5α. The specific primers are detailed in Table S2.
Bioinformatic analysis
The phylogenetic tree was built using the Neighbor-Joining method with MEGA-X (21) and visualized with ChiPlot (22). Antimicrobial resistance genes were screened using ResFinder (https://genepi.food.dtu.dk/resfinder), and mobile genetic elements (MGEs) were identified using ISfinder and BLASTN (https://blast.ncbi.nlm.nih.gov/Blast.cgi). BLASTP and BLASTN tools were used to search for homologues of FosZ and its encoding gene fosZ.
Protein structure prediction and study
The protein structures of FR-GSTs were predicted using ColabFold (https://github.com/sokrypton/ColabFold) (23), and these coordinates were visualized using PyMOL.
RESULTS
Identification of fosZ on pHS17-127
CRPA isolate HS17-127 exhibited a MDR profile (18). Both HS17-127 and its transconjugant (TcHS17-127) displayed a fosfomycin minimum inhibitory concentration (MIC) of 128 mg/L (Table 1). We found a 411 bp open reading frame (ORF) on pHS17-127, annotated as a putative glutathione S-transferase. This ORF encoded a protein of 136 amino acids, sharing 43% to 62% amino acid identity with other identified FR-GSTs (Fig. S1). We designated this new gene as fosZ, following the latest fosY nomenclature (24). Phylogenetic analysis of amino acid sequences of FosZ and other FR-GSTs revealed that FosZ was distantly related to the FosA family (less than 70%) and represented a distinct FR-GST cluster (Fig. 1A). FosZ showed the highest amino acid homology with chromosome-encoded FosA^PA^ (62%) in P. aeruginosa, and FosL1 (60%) in E. coli (Fig. S1, Fig. 1A). Amino acid alignment of FosZ with other FR-GSTs revealed conserved residues for Mn^2+^ and K^+^ cations, along with a K91R substitution in the fosfomycin-binding site (Fig. 1B).
Phylogenetic reconstruction of known fosfomycin-modifying enzymes and structural alignment of FR-GSTs. (A) Phylogenetic tree of fosfomycin-modifying enzymes constructed using neighbor-joining algorithm in MEGAX. Branch lengths are scaled by 1,000 bootstrap replications. FosA with superscript notation indicates chromosomal-encoded FosA from different species: P. aeruginosa (PA), E. cloacae (ECL), and (K. pneumoniae (KP). (B) Amino acid sequence alignment of FR-GSTs using ClustalW. The residue positions in the sequence alignment were calibrated based on FosAKP. Residues associated with fosfomycin binding, Mn2+ coordination, and K+ coordination are highlighted with black, red, and blue colored boxes, respectively (25). Black and blue dashed boxes bracket amino acids of the dimer interface loop and K+-binding loop, respectively. The asterisk symbol indicates four residue positions where PPF competes with fosfomycin for binding inhibition (26).
FosZ enzyme confers fosfomycin resistance and escapes inhibition by PPF in P. aeruginosa
To investigate the functionality of fosZ, both the gene itself and the gene along with its native promoter region were cloned and subsequently transformed into P. aeruginosa PAO1, HS355 (19) and E. coli DH5α. HS355 was a clinical MDR isolate that was susceptible to fosfomycin. The MIC of fosfomycin for the PAO1 transformant (pUCP19-fosZ) was 128 mg/L, while the PAO1 transformant with the native promoter (pUCP19-pro-fosZ) exhibited a fosfomycin MIC of 512 mg/L, an eightfold increase compared to PAO1 carrying the empty vector (Table 1). HS355 transformants (pUCP19-fosZ and pUCP19-pro-fosZ) showed a fosfomycin MIC of 512 mg/L, representing a 64-fold increase relative to HS355 (MIC 8 mg/L) (Table 1). E. coli DH5α transformant (pUCP19-pro-fosZ) demonstrated a more moderate fourfold elevation in fosfomycin MIC (Table 1). These results imply that the FosZ enzyme can reduce fosfomycin susceptibility in P. aeruginosa.
The glutathione S-transferase inhibitor PPF was tested for its inhibitory effect on FosZ, FosA^PA^ in PAO1, and FosA3 in E. cloacae 19-4074. PPF reduced the fosfomycin MIC in PAO1 from 64 to 16 mg/L and in E. cloacae 19-4074 from 1,024 to 256 mg/L. In contrast, none of the tested fosZ-bearing strains (clinical isolate, transconjugant, and transformants) exhibited any change in fosfomycin MIC (Table 1).
Origin of FosZ and genetic context of fosZ
BLASTP results revealed that FosZ was identical to a vicinal oxygen chelate (VOC) family protein (WP_065760318.1) of Pseudomonas species (27) and related to 45 VOC proteins with identities between 70.6% and 89.7% in the Nonredundant Protein Database as of 9 Oct 2024 (Table S3). Most of them (44/45, 97.9%) identified in Pseudomonas species (Table S3). Two FosZ-like proteins, FosZ-like_PAGU2196_ (WP_238216096.1) and FosZ-like LP_7_YM (WP_133752033.1) recovered from Pseudomonas sp. PAGU 2196 and LP_7_YM were closest to FosZ with identities of 89.7% and 87.5%. The fosZ gene shared 89.8% and 84.4% nucleotide similarities with these two FosZ-like-encoding genes, respectively. No mobile genetic elements were found associated with the two fosZ-like genes by ISfinder. In contrast, we found an insertion sequence (IS) of IS66 family, ISPa75 (2,996 bp in length) had entrapped fosZ in pHS17-127. There are 110 and 138 FosZ-encoding sequences in the NCBI Reference Sequence (RefSeq) project and International Nucleotide Sequence Database Collaboration (INSDC) (https://www.ncbi.nlm.nih.gov/ipg/WP_065760318.1). After eliminating duplicate strains, 159 fosZ-bearing Pseudomonas strains of various species were obtained, including P. aeruginosa (n = 145), P. putida (n = 6), P. fulva (n = 5), and one each of P. asiatica, P. oleovorans, and P. defluvii (Table S4). All the fosZ genes retrieved were located in ISPa75 with identical sequences.
Like classic IS66 family members (cIS66s), ISPa75 encompassed three ORFs encoding proteins: IS66 TnpA, IS66 TnpB, and a transposase harboring a potential DDE catalytic triad motif (Fig. 2A). Beyond that, however, ISPa75 integrated a passenger resistance gene, fosZ, downstream of the DDE transposase. Searching in the ISfinder database, we found 254 IS66 family members, and 25 were discovered in Pseudomonas species (Table S5). Seven out of 25 carried passenger genes, referred to as tIS66 (denoting a frontier between ISs and transposons) (28). ISPa75 shared 93% nucleotide identity with cIS66 ISPsk01, 91% with cIS66 ISPpu14, 93% with tIS66 ISPsp18, 90% with tIS66 ISPa128, and 90% with tIS66 ISPa120 on the left side (2263 to 2312 bp), all of which were from Pseudomonas species (Fig. 2A). The ISPsko1 had 24-bp identical sequences at terminal inverted repeat left and right (IR_L_ and IR_R_, GTAAGCGTCCGGCGAACTCACCTT), which are same to those at IR_L_ of ISPa75, ISPsp18, ISPa120, and ISPa128 (Fig. 2A).
Comparison of genetic context of fosZ with representative IS66 family members. (A) Comparison of left side of ISPa75 with ISPa75-like on the chromosome of Pseudomonas sp. SWI36, ISPsko1, ISPpu14, ISPsp18, ISPa128, and ISPa120. Below is the nucleotide sequence alignment of the IRL and IRR of ISPsko1, and the IRL of ISPa75, ISPsp18, ISPa128, and ISPa120. (B) Comparison of ISPa75 with two remnants of ΔISPa75-likeNCTC10937 and ΔISPa75-likeICMP23753. The flanking direct repeats (CGTG) of MITENCTC10937 are indicated by the paired squares. (C) Comparative analysis of ISPa75 with ΔISPa75-likeDR5-09, a 319-bp fragment containing ΔfosZ-likeDR5-09 in the chromosome of Pseudomonas sp. DR 5-09. (D) Comparative analysis of fosZ with fosZ-likePAGU2196 and fosZ-likeLP_7_YM, with short flanking sequences. (E) Nucleotide sequence alignment of the IRR of ISPa75, ΔISPa75-likeNCTC10937, ISPsy43, and ΔISPa75-likeICMP23753 and the IRL of ISPsy43 and ISPa97.
Besides, BLASTN analysis in the core nucleotide database (core_nt) revealed a putative IS66 family member located in the chromosome of Pseudomonas sp. SWI36 (CP026675.1) with 94% nucleotide similarity with ISPa75 (nt 14 to 2333, Fig. 2A). We also found two remnants of ISPa75-like elements at the right side, named ΔISPa75-like_NCTC10937_ and ΔISPa75-like_ICMP23753_ in the chromosomes of Paucimonas lemoignei strain NCTC10937 (LS483371.1, which was reidentified as a potential new species of Pseudomonas sp. by JSpeciesWS, https://jspecies.ribohost.com/jspeciesws/#home), and P. phytophila strain ICMP 23753 (CP081201.1). The 382-bp-length ΔISPa75-like_NCTC10937_ had a nucleotide similarity of 96.6% at nt 2,615 to 2,996, containing 234-bp ΔfosZ-like gene (98.3% similarity) and 148-bp right-side sequence of ΔISPa75-like (93.9% similarity) (Fig. 2B). The 284-bp ΔISPa75-like_ICMP23753_ had a nucleotide similarity of 92.0% at nt 2,711 to 2,996, containing 138-bp ΔfosZ-like gene (92.8% similarity) and 146-bp right-end ΔISPa75-like (91.8% similarity) (Fig. 2B). We discovered a new miniature inverted-repeat transposable element (MITE), MITE_NCTC10937_ of 161-bp length which truncated the fosZ-like_NCTC10937_ gene and generated 4-bp DR (CGTG) (Fig. 2B). There also existed a fragment of 319-bp length in the chromosome of Pseudomonas sp. DR 5-09 (CP011566.1) sharing a similarity of 88.1% with ISPa75 at nt 2662 to 2980, including 154-bp-length of ΔfosZ-like gene (89.6% similarity, nt 34-187) (Fig. 2C). The fosZ-like_PAGU2196_ and fosZ-like_LP_7_YM_ genes with short flanking sequences (14 to 51 bp) had similarities of 88.5% and 84.4% to fosZ and its surrounding sequences (Fig. 2D).
The IR_R_ of ISPa75 was similar to the IR_R_ of ΔISPa75-like_NCTC10937_ and the IR_L_ of ISPsy43 (tIS66) and ISPa97, with similarities of 95.8% (23/24), 91.7% (22/24), and 87.5% (21/24) respectively (Fig. 2E). The 15-bp terminal IR (GTAACCGTCCGGGCA) was highly conserved (100%, 15/15) in the IR_R_s of ISPa75, ΔISPa75-like_NCTC10937_ and ISPsy43, and IR_L_s of ISPsy43 and ISPa97 (Fig. 2E). It is supposed that fosZ was derived from an unknown Pseudomonas species and entrapped by IS66s with terminals closely related to those of ISPsko1 and ISPsy43.
The genetic arrangement of ISPa75-fosZ is depicted in Fig. 3. A putative promoter named PfosZ was identified within ISPa75 between the IR_R_ and fosZ using BPROM (http://www.softberry.com). Sequence alignment with ΔISPa75-like_NCTC10937_ and ΔISPa75-like_ICMP23753_ revealed that PfosZ is highly conserved (Fig. 3), while the PfosZ sequence is distinct from the corresponding regions in ISPsy43 and ISPa97. These findings demonstrate that ISPa75 provides the native promoter for fosZ and was captured together with it as a composite unit from the original source.
Genetic arrangement of ISPa75-fosZ. This schematic illustrates the location of the BPROM-predicted PfosZ promoter within ISPa75. The −35 and −10 boxes of the promoter are highlighted in bold and blue. IRL, left inverted repeat; IRR, right inverted repeat; DR, direct repeat.
Characteristics of fosZ-bearing plasmids
We identified ten distinct ISPa75 target site duplications (TSDs) among 159 ISPa75_fosZ-bearing Pseudomonas strains (Table S4; Fig. 4A). Of these, 34 strains were fully sequenced (Table S6), including 33 plasmids and 2 chromosomes from P. aeruginosa strains 59 and MAS152, spanning 4 Pseudomonas species and 11 provinces in China between 2011 and 2023 (Fig. 4B; Table S6).
Characteristics of 159 fosZ-bearing Pseudomonas strains from GenBank worldwide. (A) The phylogenetic tree illustrates the replication initiator gene (replicon) of fosZ-bearing Pseudomonas strains. For P. oleovorans T113 and pNY7736-1 (ΔIncP-2), the replicon could not be identified, and these strains are therefore marked solely in the outer ring of the tree. (B) Distribution of fosZ-bearing Pseudomonas strains in China.
Among the 33 fosZ-bearing plasmids, 23 featured typical IncP-2 plasmid modules (17) found in P. aeruginosa (n = 22) and P. asiatica (n = 1) and eight were Inc_pJBCL41_ megaplasmids (29) originating from P. fulva (n = 4) and P. putida (n = 4) (Table S6). The other two plasmids (p1160-VIM and pAR19438) were classified as IncP-7β (30) and Inc_pSTY-1_ (31) plasmids, respectively. Based on replicon type, backbone, and ISPa75 TSDs, these plasmids were categorized into seven groups (A to G) (Table S6). Group A (n=6, TSD9: GGAAACTC): IncP-2 plasmids resembling pHS17-127; pNY7736-1 was a disabled IncP-2 (∆IncP-2) plasmid caused by ISPpu29-mediated recombination (Fig. S2). Group B (n = 16, TSD6: GATCAAGC): The most prevalent fosZ-bearing IncP-2 plasmids across China, carrying various MDR genes (mostly blaKPC-2) (Fig. S3; Table S6). Moreover, a fused one was observed in the chromosome of P. aeruginosa 59 (Fig. S2). Group C (n=2, TSD10: AAGGCAAT): IncP-2 plasmids pSE5388-PER and pMAS152 with similar backbones but distinct TSDs with Group A and B (Fig. S4). Group D (n=5, TSD8: GACTGATC) and Group E (n=3, TSD7: CAAGGGTC) were Inc_pJBCL41_ megaplasmids (Fig. S5) (29, 32), with seven of the eight plasmids co-harboring blaVIM/blaIMP and tmexCD3-toprJ3 (Table S6). Four Group D plasmids from Henan province showed >99.9% identity, suggesting P. fulva clonal spread (Fig. S5; Table S4).
fosZ was mostly linked to intact ISPa75 elements. Notably, ISPa75s were observed both on the IncP-2 plasmid pMAS152 (at TSD10) and on the chromosome of the same strain, P. aeruginosa MAS152, at a new target site (TSD2), indicating mobilization of ISPa75 between plasmid and chromosome (Table S6). Disrupted ISPa75s were discovered in pHS204 (Group B) (33), p1160-VIM (Group F), and the chromosome of P. aeruginosa strain 59 (Fig. S6). In pHS204, tandem ISPa75s (TSD6) exhibited partial inversion (Fig. S6B). In p1160-VIM, ISPa75 (TSD5) was interrupted by an ISPa79 variant (Fig. S6C). Interestingly, P. aeruginosa strain 59 acquired partial p1160-VIM segments (including ∆ISPa75-TSD5), into which ISPpu29 inserted, followed by integration of a Group B plasmid carrying ISPa75-TSD6 (Fig. S6D).
Seven TSDs were identified across the 33 complete sequenced plasmids, associated with four incompatibility types. IncP-2 plasmids carried four TSDs, mainly in P. aeruginosa with > 15 STs. TSD9 was carried mainly by IncP-2_repA2pHS17-127 plasmids (5/6) in P. aeruginosa especially ST463 strains (3/5), while TSD6 was always carried by IncP-2repA2pHS204 plasmids (11/16) in P. aeruginosa especially ST1076 ones (3/11). There were five nucleotide differences between the repA2 of pHS17-127 and that of pHS204. TSD7 and TSD8 were associated with Inc_pJBCL41 plasmids, with TSD8 linked to clonal P. fulva strains (Table S4; Fig. 4A).
Similarly, we found ISPa75 with the same TSDs among the incompletely sequenced Pseudomonas strains carrying plasmids with the same replicons mentioned above. For instance, ISPa75_TSD5s occurred with IncP-7β plasmids (4/4), ISPa75_TSD6s with IncP-2 repA2 of pHS204 (63/66), and ISPa75_TSD10 with IncP-2 repA2 of pHS17-127 (46/46). Two novel TSDs were also identified in P. defluvii and P. oleovorans (Table S4; Fig. 4A).
Among the 159 ISPa75_fosZ-bearing Pseudomonas strains, ISPa75_TSD6 was the most prevalent type (82/159), found in at least 21 P. aeruginosa STs including ST1076 (25/81), ST485 (14/81), and ST463 (12/81) (Fig. 4A; Table S4). ISPa75_TSD10s followed (48/159), recovered in 15 P. aeruginosa STs including ST1418 (23/48) and ST357 (7/46). P. aeruginosa ST463 high-risk clones possessed three TSD types, ISPa75TSD6s (12/17) associated with IncP-2repA2_pHS204, whereas ISPa75_TSD9s (3/17) and ISPa75TSD10s (12/17) associated with IncP-2repA2_pHS17-127 (Fig. 4A; Table S4).
Geographically, fosZ-bearing Pseudomonas strains were almost exclusively from China (98.1%, 156/159), especially Zhejiang province (82/156), Hubei province (17/156), and Shanghai (15/156) (Fig. 4B). In addition to clinical sources, isolates were recovered from environmental settings, sewage, chicken, migratory birds, and flies (Fig. 4A).
Protein structure analysis of FR-GSTs
We employed ColabFold to predict the protein structures of FosZ (Fig. 5A), FosA-family members (Fig. S7), and other FR-GSTs (Fig. S8). The predicted structures of all dimeric FR-GST proteins exhibited the typical VOC superfamily topology (27), consisting of two βαβββ units that form an incompletely closed β-sheet barrel around the metal ion. The superimposition of FosZ with all predicted FR-GSTs revealed substantial structure conservation, with Cα root mean square deviations (RMSD) values ranging from 0.39 to 0.71 Å.
Structural analysis of FosZ. (A) An overall alignment of FosZ with previously well-characterized FosAPA (PDB code 1LOP) in P. aeruginosa and FosAKP (PDB code 5V91) in K. pneumoniae. The superimposition of all three protein structures reveals an RMSD value of less than 0.7 Å between any two of them. Colors listed below each panel correspond to FosA sequences shown in all panels. Mn2+ ion and K+ ion are shown as colored spheres. (B) Comparison of the dimer interface loop in FosZ, FosA,KP and FosAPA. The cα RMSD values of loop dimensions in FosZ (12.7 Å, A56 in monomer one to R61 in monomer 2), FosAPA (9.4 Å, G57 in monomer one to A60 in monomer 2), and FosAKP (18.2 Å, V57 in monomer one to P59 in monomer 2) are represented as dashed lines. Y65, which bridges the active site and the dimer interface loop, is labeled as sticks. (C) Superimposition of FosZ with predicted FR-GSTs. The black dashed boxes denote expanded K+-binding loops in FosZ. The black box represents the K+-binding loop residues in FosAKP (R96 to G99), FosAPA (R93 to G96), and FosZ (K94 to G97). K94 in FosZ exhibits a Cα RMSD deviation of 2.7 Å from FosAKP and 3.3 Å from FosAPA. S95 in FosZ exhibits a cα RMSD deviation of 2.9 Å from FosAKP and 3.5 Å from FosAPA.
The length of the FosA dimer interface loop (FosA^KP^ peptide 48-64) impacts enzyme activity, FosA3 and FosA^KP^ with longer and more extended loops conferred greater fosfomycin resistance compared to FosA^PA^ (25). We calculated the Cα RMSD values between residues located at the dimer interface loops to quantify the loop dimensions. All FosA-family members had 17-residue-length dimer interface loops (Fig. 1B), with loop dimensions ranging from 17.5 to 19.4 Å (Fig. S7). To be noted, this loop in FosZ was two amino acid residues shorter than that in FosA^KP^ (with a loop dimension of 12.8 Å) but was longer and more extended than that in FosA^PA^ (Fig. 5B). In the FosL1 structure, the loop was two residues shorter than FosA^KP^ (Fig. 1B), with a loop dimension of 13.7 Å (Fig. S8C), which was similar to FosZ. In the structures of FosC2, FosG, FosF, and FosK, the loops were five amino acid residues shorter than in FosA^KP^ (Fig. 1B), with loops dimensions ranging from 5.3 to 5.4 Å (Fig. S8D through G), and crossed the dimer interface directly like FosA^PA^ (25).
In the binding pocket near the fosfomycin and/or PPF binding site, FosZ had two amino acid substitutions (K93R and N95D, according to the amino acid numbering of FosA^KP^) compared to all FR-GSTs (Fig. 1B). The mobile K^+^-binding loop in FosZ showed deviations when superimposed with all predicted FR-GST structures, exemplified by FosA^KP^ and FosA^PA^ (Fig. 5C). Residue S97 in FosZ was among the four residues involved in binding both fosfomycin and PPF (Fig. 1B) (26).
DISCUSSION
In recent times, fosfomycin, an antibiotic with a decade-long history, has surged into the spotlight as a potential remedy for combating MDR/XDR bacterial infection. Previous studies (5, 6) have primarily investigated chromosomal mutations as the dominant mechanism of fosfomycin resistance in P. aeruginosa. Here, we first characterized the plasmid-borne fosfomycin resistance gene fosZ encoding glutathione S-transferase FosZ, which conferred fosfomycin resistance in P. aeruginosa. The fosZ gene was captured by translocatable genetic element ISPa75, which may facilitate the dissemination of fosfomycin resistance among different Pseudomonas species, predominantly through ISPa75-medicated transposition within IncP-2 plasmids among P. aeruginosa, particularly in MBL-positive isolates. An IncP-2 plasmid sublineage has been linked to the dissemination of blaIMP-45 among CRPA in the intensive care unit (ICU) of a hospital in Shanghai, China (17). IncP-2 plasmid has accumulated multiple resistance genes during its evolution, including those conferring resistance to aminoglycosides, antipseudomonal carbapenems, cefiderocol (blaPER-1), ceftolozane-tazobactam (blaKPC-33), and fosfomycin, facilitating cross-species transfer of resistance genes. The fosZ-bearing Inc_pJBCL41_ megaplasmid pZXPA-20-602k from P. putida of migratory bird origin has been reported as a convergence of blaVIM-2 and tmexCD1-toprJ1 (32). We observed the spread of Inc_pJBCL41_ megaplasmid from P. fulva, which co-carried blaVIM-24, tmexCD3-toprJ3, and fosZ in a hospital in Henan, China, in 2019. These MDR megaplasmids may serve as reservoirs for resistance genes against carbapenems, tigecycline, and fosfomycin, presenting significant obstacles for clinical treatment and infection control.
Fosfomycin combined with new antimicrobials is gaining attention for treating MDR-PA infections (34, 35). The activity of such regimens in P. aeruginosa infections remains debated and has been explored in several in vitro studies. The ceftazidime/avibactam and fosfomycin combination significantly reduced P. aeruginosa colony-forming units, surpassing individual drugs and offering a practical alternative for MBL-negative isolates (3). The ceftolozane/tazobactam and fosfomycin combination displayed synergy in time-kill analyzes against MBL-positive isolates (4). CLSI lacks fosfomycin breakpoints specific for P. aeruginosa. European Committee on Antimicrobial Susceptibility Testing (EUCAST) mentions that (36), for wild-type P. aeruginosa isolates (with an epidemiological cutoff value of MIC 128 mg/L), fosfomycin has been employed in combination therapy. Our findings provide further evidence of transferable sources of fosfomycin resistance among Pseudomonas species. This may limit its clinical applicability in combination therapy for MDR-PA infections.
PPF (Foscarnet) is clinically used mainly for treating viral infections. In vitro studies suggest that at therapeutic plasma concentrations, PPF can enhance fosfomycin’s activity against some bacteria that produce GST (Table S1), potentially reversing fosfomycin resistance in GST-producing Gram-negative bacteria. This synergy requires further clinical validation but highlights a novel combinatorial strategy against resistant pathogens (15, 16). Our findings suggest that FosZ cannot be inhibited by PPF, which may be attributed to the structural differences between FosZ and other FR-GSTs like FosA^KP^ and FosA^PA^. FosZ exhibits an expanded K^+^-binding loop pocket, leading to deviation of residue S97, which participates in binding both the inhibitor and substrate (26). This structural alteration may reduce the competitive inhibition efficacy of PPF against fosfomycin. However, this remains a structural speculation and necessitates further biochemical evidence to support it.
Our BLAST analysis identified FosZ-like proteins in the chromosomes of many Pseudomonas species, suggesting that fosZ likely originated from Pseudomonas species. IS66 family elements (e.g., unknown IS in Pseudomonas spp. SWI36, and ISPsko1) likely captured a fosZ-like sequence, leading to the formation of ISPa75.
Some FosA variants such as FosA1, FosA2, and FosA11 exhibit amino acid homology below 80% or 70% with other FosA-family members but share similar protein structures, notably in the 17-residue-length dimer interface loops. The dimer interface loop serves as a key structural feature of FR-GSTs, distinguishing FosA-family members from other FR-GSTs, such as FosZ, which has a 15-residue-length loop.
In our recent research, we identified eight IncP-2 plasmids from CRPA isolates carrying blaPER-1 and ISPa75-fosZ (33). The fosZ gene, trapped in ISPa75, is now prevalent across both southern and northern China, particularly in Zhejiang province, Hubei province, and Shanghai, over the past 14 years (2011 to 2024). ISPa75 has been transferred among different plasmid incompatible groups, including IncP-2, Inc_pJBCL41_, IncP-7β, and Inc_pSTY-1_, and between plasmid and chromosome within the same isolate. The presence of multiple ISPa75 target sites across different plasmids, coupled with its relocation to various positions within plasmids of the same incompatibility type, demonstrates ISPa75’s high transferability. This mobility facilitates the dissemination of resistance genes among diverse species. Furthermore, this may reflect the evolutionary pressures faced by Pseudomonas species, which enables them to adapt quickly to changing environments and enhances their survival in the presence of antimicrobial agents. Group B plasmids, the most common fosZ-bearing IncP-2 types, have accumulated various MDR genes and have been captured by high-risk ST463 clones.
In summary, the emergence of the plasmid-borne fosfomycin resistance determinant fosZ poses critical concerns in antimicrobial stewardship, particularly given fosfomycin’s indispensable role as a combinatorial agent against MDR-PA isolates, specifically MBL-producing ones with extremely restricted treatment options.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Horcajada JP, Montero M, Oliver A, Sorlí L, Luque S, Gómez-Zorrilla S, Benito N, Grau S. 2019. Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clin Microbiol Rev 32:e 00031-19. doi:10.1128/CMR.00031-1931462403 PMC 6730496 · doi ↗ · pubmed ↗
- 2Falagas ME, Vouloumanou EK, Samonis G, Vardakas KZ. 2016. Fosfomycin. Clin Microbiol Rev 29:321–347. doi:10.1128/CMR.00068-1526960938 PMC 4786888 · doi ↗ · pubmed ↗
- 3Papp-Wallace KM, Zeiser ET, Becka SA, Park S, Wilson BM, Winkler ML, D’Souza R, Singh I, Sutton G, Fouts DE, Chen L, Kreiswirth BN, Ellis-Grosse EJ, Drusano GL, Perlin DS, Bonomo RA. 2019. Ceftazidime-avibactam in combination with fosfomycin: a novel therapeutic strategy against multidrug-resistant Pseudomonas aeruginosa. J Infect Dis 220:666–676. doi:10.1093/infdis/jiz 14931099835 PMC 6639593 · doi ↗ · pubmed ↗
- 4Cuba GT, Rocha-Santos G, Cayô R, Streling AP, Nodari CS, Gales AC, Pignatari ACC, Nicolau DP, Kiffer CRV. 2020. In vitro synergy of ceftolozane/tazobactam in combination with fosfomycin or aztreonam against MDR Pseudomonas aeruginosa. J Antimicrob Chemother 75:1874–1878. doi:10.1093/jac/dkaa 09532240299 · doi ↗ · pubmed ↗
- 5Zheng D, Bergen PJ, Landersdorfer CB, Hirsch EB. 2022. Differences in fosfomycin resistance mechanisms between Pseudomonas aeruginosa and Enterobacterales Antimicrob Agents Chemother 66:e 0144621. doi:10.1128/AAC.01446-2134807759 PMC 8846481 · doi ↗ · pubmed ↗
- 6Ito R, Mustapha MM, Tomich AD, Callaghan JD, Mc Elheny CL, Mettus RT, Shanks RMQ, Sluis-Cremer N, Doi Y. 2017. Widespread fosfomycin resistance in Gram-negative bacteria attributable to the chromosomal fos A gene . m Bio 8:e 00749–17. doi:10.1128/m Bio.00749-1728851843 PMC 5574708 · doi ↗ · pubmed ↗
- 7Wachino J, Yamane K, Suzuki S, Kimura K, Arakawa Y. 2010. Prevalence of fosfomycin resistance among CTX-M-producing Escherichia coli clinical isolates in Japan and identification of novel plasmid-mediated fosfomycin-modifying enzymes. Antimicrob Agents Chemother 54:3061–3064. doi:10.1128/AAC.01834-0920404116 PMC 2897269 · doi ↗ · pubmed ↗
- 8Guo Q, Tomich AD, Mc Elheny CL, Cooper VS, Stoesser N, Wang M, Sluis-Cremer N, Doi Y. 2016. Glutathione-S-transferase Fos A 6 of Klebsiella pneumoniae origin conferring fosfomycin resistance in ESBL-producing Escherichia coli. J Antimicrob Chemother 71:2460–2465. doi:10.1093/jac/dkw 17727261267 PMC 4992852 · doi ↗ · pubmed ↗