A recombinant lysogenic bacteriophage inhibits Salmonella virulence
Erick Maosa Bosire, Mudasir Ali Rather, Katherine E. Bell, Paulina D. Pavinski Bitar, Ranju Ravindran Santhakumari Manoj, Craig Altier

TL;DR
Scientists engineered a bacteriophage to produce a chemical that inhibits Salmonella's ability to cause disease in the gut.
Contribution
A recombinant phage was created to deliver c2HDA, reducing Salmonella virulence and gut inflammation.
Findings
Salmonella with the engineered phage produced c2HDA and repressed virulence gene expression.
The phage reduced Salmonella invasion of epithelial cells by over 100-fold.
Phage carrying rpfF reduced gut inflammation more effectively than phage without the gene.
Abstract
The gut environment includes an abundance of chemicals emanating from the host and the microbiota. To colonize animals, Salmonella uses gut chemicals as locational cues to ensure expression of energy-intensive virulence factors only when their production is necessary. cis-2-hexadecenoic acid (c2HDA), a member of the diffusible signal factor family of quorum-sensing signals, potently represses virulence-gene expression by Salmonella. Here, we report the construction and use of a recombinant bacteriophage that can establish lysogeny within Salmonella and induce it to produce c2HDA, thus repressing functions essential to its own virulence. We engineered the temperate phage P22 to favor lysogeny through transposon mutagenesis of the sieB-esc region and caused it to produce c2HDA by introduction of rpfF of Cronobacter turicensis, encoding the dehydratase/thioesterase required for c2HDA…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
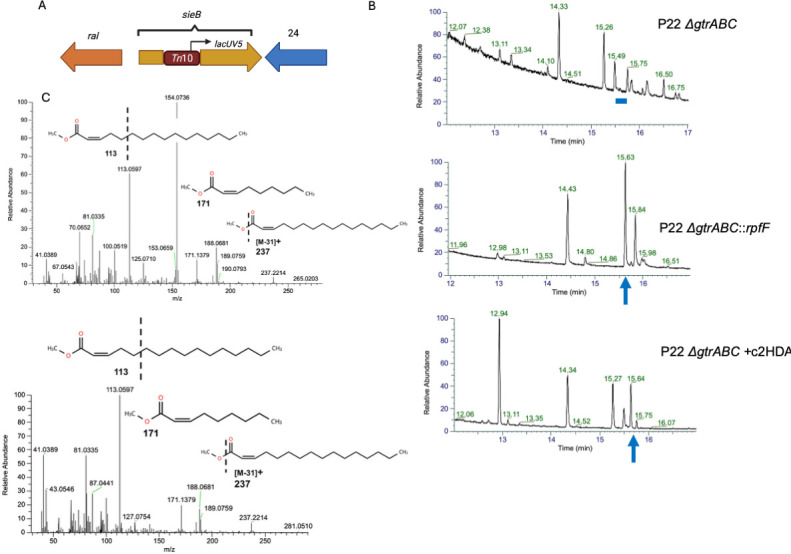 Fig 1
Fig 1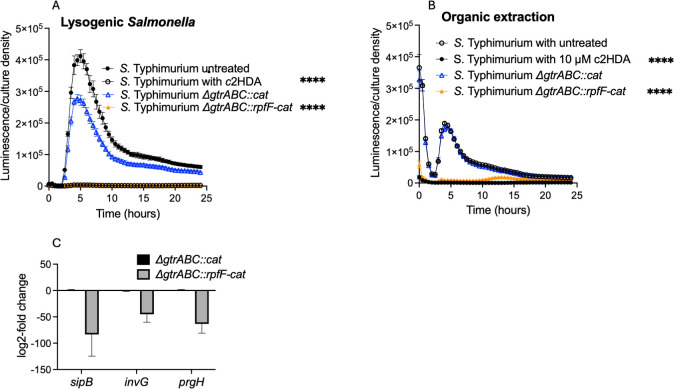 Fig 2
Fig 2 Fig 3
Fig 3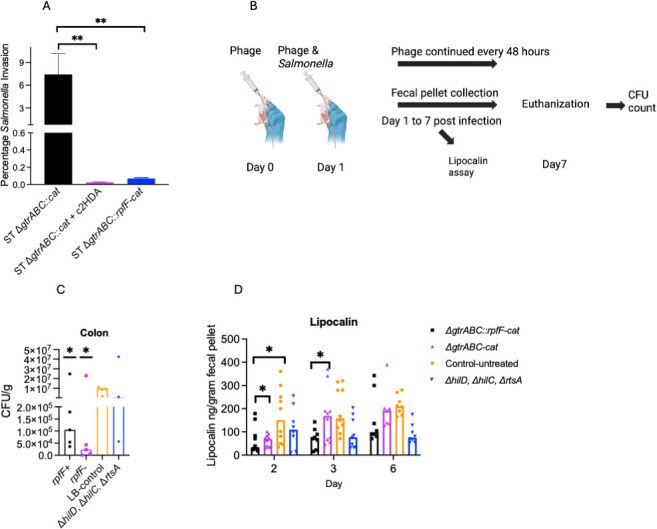 Fig 4
Fig 4| Primer name | Sequence |
|---|---|
| RT-PCR- |
|
| RT-PCR- |
|
| RT-PCR- |
|
| RT-PCR- |
|
| RT-PCR- |
|
| RT-PCR- |
|
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
| |
|
|
| Strain/plasmid | Genotype/sequence | Reference |
|---|---|---|
| CA32 | ATCC | |
| CA5395 | P22 | This study |
| CA2286 | ( | |
| CA5594 | Δ | This study |
| CA5560 | Δ | This study |
| CA5606 | Δ | This study |
| CA5602 | Δ | This study |
| CA5598 | Δ | This study |
| CA5567 | Δ | This study |
| CA5484 | This study | |
| CA5745 | Δ | This study |
| CA5744 | Δ | This study |
| CA4434 | ( | |
| CA3996 | ( |
- —Agriculture and Food Research Initiative Competitive Grant
- —NIH/NIAID
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Salmonella and Campylobacter epidemiology · Bacterial biofilms and quorum sensing
INTRODUCTION
As Salmonella traverses the animal intestinal tract, it encounters changing chemical environments created by both the host and the resident microbiota. This pathogen has evolved, however, not only to survive within these environments but also to recognize the changes and to use them as cues to promote its own survival and replication. In the proximal small intestine, Salmonella first encounters high concentrations of bile, which represses its type III secretion apparatus required for the invasion of intestinal epithelial cells (1, 2). Within the distal small intestine, bile dissipates and the fatty acid formate predominates (3), inducing this same apparatus and creating a nidus of cell invasion and ensuing inflammation (4, 5). In more distal parts of the intestine, chemical signals repressive to invasion again predominate, including short- and long-chain fatty acids (6, 7), allowing Salmonella to dedicate its energy instead to growth and replication (8).
Among the most potent chemical repressors of Salmonella invasion is cis-2-hexadecenoic acid (c2HDA) (9). This molecule is a member of the diffusible signal factor (DSF) class of quorum-sensing signals that is produced and recognized by several bacterial species (10). Among these species, the activity of DSFs is typical of quorum-sensing systems, regulating aggregation, motility, and the formation and dispersion of biofilms (11–14). Their effects on Salmonella, however, are quite different. At micromolar concentrations, c2HDA inhibits Salmonella invasion by binding and preventing the activity of three AraC-type transcriptional activators, HilD, HilC, and RtsA (9, 15, 16), that function collaboratively to induce invasion (17). The inactivation of these regulators thus profoundly reduces the expression of the genes needed for invasion and the penetration of epithelial cells (9, 15, 16).
The hallmark structure of DSF quorum-sensing signals essential to their function is an unsaturation at the second position in the cis orientation (10). Their production is catalyzed from existing pools of long-chain fatty acids by a dehydratase/thioesterase typically termed RpfF, which both creates the double bond and cleaves the thioester linkage of the fatty acid to acyl carrier protein (18). We have previously shown that rpfF, when expressed in a probiotic strain of Escherichia coli, produces c2HDA in sufficient quantities to inhibit the invasion of Salmonella when grown in co-culture. Furthermore, this c2HDA-producing strain can colonize the gut of chickens and reduce colonization of Salmonella in experimental infection trials (19).
These findings thus suggest that c2HDA might be used as a practical means to reduce Salmonella carriage in animals. The efficacy of this approach, however, likely depends upon the achievable concentration of this molecule at its site of action, the animal intestine. The question then becomes one of efficient in situ delivery. Here, we test a novel approach, engineering a recombinant lysogenic bacteriophage that harbors rpfF and produces c2HDA. We find that this phage induces Salmonella to endogenously produce its own c2HDA, thus efficiently repressing its own invasion. The repression of this essential virulence function results in attenuated invasion of epithelial cells and consequently reduced gut inflammation.
RESULTS
A recombinant lysogenizing phage produces a signal repressive to Salmonella virulence
c2HDA, a member of the DSF class of quorum-sensing molecules, can potently repress Salmonella genes required for virulence (9). This molecule can be produced recombinantly in E. coli, effectively inhibiting the virulence of Salmonella in co-culture (19). We reasoned that this repressive system could be made more efficient by inducing Salmonella to produce its own c2HDA through infection by a lysogenizing recombinant phage engineered to generate the signal. We first created a mutant of the temperate phage P22, which infects S. Typhimurium and is widely used as a genetic tool (20), to favor lysogeny over lytic growth. Using random transposon mutagenesis, we selected for lysogens under conditions that typically promote lysis. One such mutant was found to harbor its transposon insertion in the sieB-esc region, 254 bp downstream from the sieB initiating codon. sieB encodes a superinfection exclusion system that prevents the lytic process by superinfecting phages (21). esc encodes a peptide within the sieB ORF that prevents the production of full-length SieB and thus protects the phage from its own destruction (22). An insertion in this region would thus disrupt both ORFs, allowing superinfection by infecting bacteriophage. To test whether this mutation induced lysogeny over lytic growth, we compared it to a derivative of P22, harboring a precise deletion of sieB, previously shown to produce lysogens at a frequency unchanged from that of wild-type P22 (23). Comparing transductants by selection on media containing kanamycin (as both phage are resistant to this antibiotic) to plaque-forming units (pfu), we found that our mutant phage yielded lysogens at a rate of 2.5 transductants/pfu, compared to 1.25 transductants/pfu for the control phage, yielding a twofold advantage. The mechanism by which this mutation favors lysogeny over lytic growth is unclear. It should be noted, however, that the insertion lies adjacent to gene 24 (24), a λ gene N homolog encoding an antiterminator required for the expression of lytic phage proteins (25) (Fig. 1A). The transposon itself carries an outward-facing lacUV5 promoter (26), oriented in an antisense direction to gene 24. Effects on gene 24 might thus reduce the efficiency of phage lysis. We next introduced into this phage rpfF of Cronobacter turicensis, encoding a dehydratase/thioesterase that places the distinctive cis-2 unsaturation characteristic of DSF quorum-sensing signals into saturated long-chain fatty acids (12, 18). The gene was expressed under the control of a constitutive promoter and replaced the P22 gtrABC operon, which causes Salmonella seroconversion (24), but is not essential for phage infection or replication. We tested this strain for DSF production, comparing it to an isogenic lysogenic strain harboring a P22 ΔgtrABC mutant without rpfF. GC-MS analysis showed that a long-chain fatty acid was produced by lysogens harboring the rpfF-encoding phage, but not by those with the control phage (Fig. 1B). Mass fragmentation pattern analysis further revealed this product to be c2HDA, based upon its comparison to the fragmentation pattern of the commercially available c2HDA (Fig. 1C). These results thus show that Salmonella can be infected with this recombinant phage to produce stable lysogens and induce endogenous production of a foreign quorum-sensing signal.
P22 phage lysogens carrying the rpfF gene produce c2HDA signal. (A) Construction of lysogens. Mutants with the transposon inserted at the sieB-esc region were more lysogenic. (B) GC-MS retention time analysis. The blue arrow shows the peak with the expected retention time for c2HDA (15.63 min, which represents the time this compound eluted from the chromatography column after injection) produced by the strain carrying P22 ΔgtrABC::rpfF (second from top), and a dash indicates the missing c2HDA peak in P22 ΔgtrABC strain (top). This retention time was confirmed by spiking c2HDA into extracts of P22 ΔgtrABC lacking the 15.63 min peak as indicated (bottom chromatogram). (C) MS fragmentation of the 15.63 peak for the commercial c2HDA (top) and that produced by the P22 ΔgtrABC::rpfF lysogens (bottom).
Production of c2HDA by the recombinant lysogenic phage reduces the expression of Salmonella invasion genes
When supplied exogenously, members of the DSF class of quorum-sensing molecules, including c2HDA itself, are known to inhibit Salmonella invasion of epithelial cells, a function required for virulence (9). We sought to determine whether this signal would act similarly when produced endogenously by Salmonella itself. To test this, we first measured the expression of hilA, which encodes an activator of the invasion cascade (27), using a hilA-luxCDABE transcriptional reporter fusion. Wild-type Salmonella, without lysogenized P22, demonstrated the characteristic expression of hilA in laboratory medium, with a rapid induction and peak expression during exponential growth (9) (Fig. 2A). A strain carrying the control phage, with a gtrABC deletion and associated chloramphenicol-resistance gene but lacking rpfF (ΔgtrABC::cat), showed a modest but significant reduction in peak hilA expression of 30%. In the c2HDA-producing lysogen (ΔgtrABC::rpfF-cat), however, hilA expression was reduced profoundly, by 132-fold at peak expression, in comparison to the control lysogen strain. This repression was similar in its proportion to that caused by the addition of 20 μM c2HDA to the medium (115-fold). The presence of neither lysogenic phage, however, affected the growth of the strains, demonstrating that the effects on hilA expression were independent of growth-phase induction. To ensure that the effects observed were, in fact, due to the production of a diffusible signal of the DSF class, we performed an organic extraction on the culture supernatant. When added to growing cultures of the hilA-luxCDABE reporter strain, extract obtained from the ΔgtrABC::rpfF-cat lysogen strain reduced peak hilA expression by 19-fold, compared to extract from the ΔgtrABC::cat lysogen control strain, which did not differ from the untreated control (Fig. 2B). By comparison, the addition of 10 μM c2HDA reduced expression by 156-fold in this assay.
*P22 ΔgtrABC::rpfF lysogenic phage-produced c2HDA reduces expression of invasion genes. (A) Lysogenized phage carrying rpfF represses hilA expression in Salmonella grown in laboratory media. Lysogens additionally carrying PhilA-luxCDABE were cultured under SPI1-inducing conditions. Data are presented as mean luminescence corrected for culture density for five biological replicates. (B) Organic extracts from lysogenized phage carrying rpfF repress hilA expression. Culture supernatants were organically extracted, dissolved in ethanol, and added to growing cultures of the WT strain carrying a PhilA-luxCDABE reporter fusion. (C) Lysogenized phage reduces message levels of invasion genes. Strains carrying phage with and without rpfF were cultured under SPI1 inducing conditions; mRNA was extracted and quantified by qRT-PCR. Results were normalized using the rpoD as the housekeeping gene. Specific primers used are listed in Table 1. Gene expression is presented as log-2-fold change. Error bars represent standard deviations of three biological replicates. Asterisks denote significant differences from the control (***P < 0.0001). Statistical significance was tested using Student’s t-test, and the Mann-Whitney test was applied to compare ranks.
To determine whether repression of the activator hilA elicited effects on the invasion genes it controls, we compared their expression in the ΔgtrABC::cat and ΔgtrABC::rpfF-cat lysogen strains using qRT-PCR. Both prgH and invG encode components of the Salmonella SPI1 needle complex (28), required for secretion of effector proteins into epithelial cells, while sipB encodes one of those secreted effectors (29). Expression of all these genes was greatly reduced in the ΔgtrABC::rpfF-cat lysogen strain, 69-, 42-, and 60-fold (for sipB, invG, and prgH, respectively), when compared to the isogenic lysogen harboring ΔgtrABC::cat that does not produce c2HDA, demonstrating that production of c2HDA through infection by a recombinant phage can efficiently repress functions required for Salmonella virulence (Fig. 2C).
Endogenously produced c2HDA can act as an exogenous signal
Although these results demonstrate that c2HDA is produced within the cytoplasm of lysogenized strains, it is possible that Salmonella also secretes this signal into its surrounding environment to affect the expression of neighboring bacteria, as we have previously found for recombinant rpfF-expressing E. coli (19). To test this, we used a co-culture approach. The reporter strain in this case carried the hilA-luxCDABE transcriptional reporter fusion in an otherwise wild-type strain background and was grown together and in equal proportion with strains harboring the c2HDA-producing or control lysogenic phage. We then compared hilA expression to that of the two lysogenized strains that harbor their own hilA-luxCDABE fusions, described above. The reporter strain grown in co-culture with the control lysogen demonstrated a modest reduction in hilA expression, by 33%, similar to that of the strain carrying both the control phage and the reporter together. Co-culture of the c2HDA-producing lysogen, however, severely reduced the expression of hilA in neighboring bacteria, by 55-fold compared to the control lysogen (Fig. 3A). This reduction was, in fact, indistinguishable from that found through endogenous production of c2HDA by the strain harboring both the c2HDA-producing phage and the hilA reporter fusion (56-fold reduction). These results suggest that c2HDA produced by the recombinant phage is secreted into the culture medium, affecting expression of virulence genes in nearby Salmonella. Alternatively, however, the phage itself might infect these neighboring bacteria. Although the phage has been engineered to promote lysogeny, lytic growth can and does occur, raising the possibility that transfer of rpfF to naïve hosts induces endogenous c2HDA production. To test this possibility, we utilized Salmonella serovar Enteritidis, as phage P22 infects this serovar very poorly due to the activity of a restriction system not found in S. Typhimurium (30). We employed a dual-fluorescent Salmonella Enteritidis reporter strain constitutively producing blue fluorescent protein (BFP) used to determine total numbers of Salmonella, and encoding the gene for GFP linked to the promoter of the invasion gene sicA within the chromosome, assessing the proportion of the population that expressed sicA by flow cytometry (31). When grown alone in culture medium, 84% of the population of the S. Enteritidis reporter strain demonstrated detectable expression of sicA (Fig. 3B). Co-culture with the S. Typhimurium control lysogen strain lacking rpfF produced a small reduction in sicA expression to 73% of the population. Co-culture of S. Enteritidis with the S. Typhimurium strain bearing the c2HDA-producing prophage, however, significantly reduced the sicA-expressing population to 45%. These results, taken together, thus demonstrate that infection of Salmonella with a phage encoding rpfF and producing c2HDA can repress genes required for virulence both directly, through endogenous action of the quorum-sensing signal, and indirectly, by effects on the Salmonella bacteria that share their environment but have not themselves been infected.
*Strains carrying lysogenized phage with rpfF produce c2HDA that exogenously represses invasion genes in neighboring bacteria. (A) Co-culture assay of wild type carrying PhilA-luxCDABE reporter fusion and strains harboring lysogenized phage with or without rpfF. As controls, the wild-type and the lysogenized phage strains both carrying the PhilA-luxCDABE were cultured separately for comparison. Error bars represent standard deviations of five biological replicates. (B) Lysogenized phage produce c2HDA that can exogenously repress invasion-gene expression in S. Enteritidis. S. Enteritidis strain carrying a sicA-GFP reporter fusion and constitutively expressing BFP was co-cultured with an S. Typhimurium carrying lysogenic phage with or without rpfF. The number of bacteria expressing GFP was assessed by flow cytometry, and BFP was used to gate for S. Enteritidis. Abbreviations: ST, Salmonella Typhimurium and SE, Salmonella Enteritidis. Data are presented as percentage of bacteria expressing sicA for five biological replicates. Asterisks denote significant differences from the control (*P < 0.05, ***P < 0.0001). Statistical significance was tested using the Student’s t-test, and the Mann-Whitney test was applied to compare ranks.
Production of c2HDA by infection with a recombinant phage inhibits Salmonella invasion and consequently inflammation
Repression of Salmonella invasion genes can reduce the ability of this pathogen to penetrate the intestinal epithelium, an early required function for both Salmonella carriage and disease. To test whether c2HDA production by a lysogenized phage reduced invasion, we used a well-established gentamicin-protection cell culture assay. HEp-2 cells were inoculated with Salmonella strains, treated with gentamicin to eliminate extracellular bacteria, and then lysed to assay the number of bacteria that had invaded the cells. We found that for the control strain carrying the ΔgtrABC::cat lysogenized phage, 7.4% of the inoculated population invaded cells (Fig. 4A). For the c2HDA-producing ΔgtrABC::rpfF-cat lysogen, however, invasion was reduced by more than 100-fold, to 0.07%. As a control, we grew the ΔgtrABC::cat control strain in the presence of 20 μM c2HDA prior to the assay, which reduced invasion by >300-fold, to 0.02%. The production of c2HDA and its repression of virulence genes thus has the functional consequence of reduced cell invasion.
Our data demonstrated that a lysogenized phage bearing rpfF can reduce penetration of Salmonella into epithelial cells through its repressive effects on invasion genes. We therefore anticipated that lysogenized phage would reduce expression of Salmonella virulence genes in the gut and consequently its ability to cause inflammation and colonize the gut. To test the effects of the phage under native gut conditions without alterating gut hemeostasis, we used 129X1/SvJ mice that can be colonized by Salmonella without pre-treatment with antibiotics (32). We administered the ΔgtrABC::rpfF-cat phage or the ΔgtrABC-cat phage to mice and then infected them with Salmonella (Fig. 4B). Phage treatment was continued every 48 h for 7 days. We found that administration of either the phage harboring rpfF or the control phage without it greatly and similarly reduced Salmonella colonization compared to untreated controls (Fig. 4C), by ~93- and 420-fold. This may be attributed to the ability of the P22 phage to lyse Salmonella. While we engineered the recombinant phage for lysogeny, it is possible that conditions in the gut could induce lytic growth, resulting in reduction of Salmonella viability. Inflammation levels were then tested using fecal pellets for levels of lipocalin-2, a common marker of inflammation induced by Salmonella. We found that the phage carrying rpfF reduced inflammation levels by ~5-fold, significantly more than by the isogenic phage bearing no rpfF, which reduced lipocalin-2 by ~3-fold (Fig. 4D). Invasion-gene expression is controlled by the central AraC-type transcriptional regulators HilD, HilC, and RtsA (17). Mutants of these regulators have reduced expression of effectors that are required for Salmonella to elicit inflammation in the gut (33, 34). As HilD, HilC, and RtsA are targets of c2HDA (9, 15, 35), we compared the levels of inflammation to those of mice infected with a ΔhilD, ΔhilC, ΔrtsA mutant strain. We found that inflammation in mice treated with phage carrying rpfF was reduced to a level close to that of mice that received the ΔhilD, ΔhilC, ΔrtsA strain (Fig. 4D). This indicates that orally administered recombinant phage bearing rpfF can infect Salmonella in the gut and induce it to produce c2HDA, repressing virulence effectors and thus reducing inflammation.
*c2HDA produced by lysogenized phage inhibits invasion of epithelial cells and reduces inflammation in the gut. (A) Invasion of HEp-2 cells by Salmonella carrying lysogenized phage with or without rpfF. Data are presented as the percentage of Salmonella invading HEp-2 cells. Error bars denote standard deviation of five biological replicates. Asterisks denote significant differences from the untreated control (**P < 0.01, *P < 0.05). Abbreviations: ST, Salmonella Typhimurium. (B) Lysogenized phage carrying rpfF reduces inflammation in the gut. Schematic showing the procedure for treating mice with the recombinant phage. (C) Salmonella load in the colon at day 7 post-infection. Asterisks denote significant differences from the mice inoculated with the wild type but not treated with phage. (D) Inflammation levels in the fecal pellets. Inflammation was assessed using the lipocalin-2 assay. Data are presented as lipocalin per gram of fecal pellet. Each group contained nine mice, and asterisks denote significant differences between indicated groups (P < 0.05). Statistical significance was tested using the Student’s t-test, and the Mann-Whitney test was used to compare ranks.
DISCUSSION
Here, we have described a bacteriophage engineered to deliver to Salmonella a single gene that induces this pathogen to repress its own virulence. Rather than attempting to kill the organism, this form of phage therapy instead seeks to prevent Salmonella survival and proliferation by eliminating a function required for colonization of an animal host. This is achieved through the endogenous production of a quorum-sensing molecule, c2HDA, normally used by Salmonella as an exogenous signal to modulate its virulence.
c2HDA can be found in the animal intestine and therefore likely represents a means of interspecies chemical signaling, through which Salmonella “eavesdrops” on the quorum-sensing of neighboring bacteria to recognize its environment and modulate its gene expression, maximizing its own survival. The potential to use this system to mitigate salmonellosis therefore requires that c2HDA be delivered to Salmonella within the animal intestine in concentrations greater than those that occur naturally, essentially undermining the carefully controlled virulence program of this pathogen. If provided directly through oral inoculation, members of the DSF class of quorum-sensing molecules, being long-chain fatty acids, would be rapidly absorbed by the enterocytes of the small intestine. To circumvent this problem, we have previously created a recombinant probiotic E. coli strain that can readily colonize the gut and deliver c2HDA to the surrounding environment (19). The production of this molecule within Salmonella itself holds promise to improve the efficiency of this mitigation strategy, due to the high local concentrations likely to ensue.
The development of resistance to bacteriophage by their bacterial targets remains a persistent obstacle to the use of phage therapy as a tool for disease treatment or prevention. Salmonella has indeed been shown to develop resistance to P22, the phage used here, by multiple means (36). The proliferation of such mutants might, however, be mitigated at least in part by the loss of fitness or virulence that such mutations can impart (37). Additionally, P22 is but one of many related phage, sensitivity to which has long been used to determine the serotypes of Salmonella isolates (38). The availability of a well-characterized set of such phage in a genetically tractable organism such as Salmonella might provide useful alternatives to P22 that could extend and augment this approach.
Treating or preventing infection by enteric pathogens is particularly difficult due to the complex bacterial milieu that surrounds them. The use of broad therapies such as antibiotics risks wholesale disruption of the intestinal microbiota, resulting in widespread dysbiosis with severe consequences to health. The use of phage instead offers the possibility of a highly targeted disease-mitigation strategy. Infection by P22 and other phage of Salmonella can be directed to specific serovars and, in some cases, to strains within serovars. They therefore offer the possibility of directed prevention, targeting the Salmonella types found in specific animal species, locations, and production systems.
MATERIALS AND METHODS
Strains, plasmids, and growth conditions
Salmonella enterica subsp. enterica serovar Typhimurium 14028s and Salmonella enterica subsp. enterica serovar Enteritidis MD15, and mutants thereof, were used throughout, as listed in Table 2. All bacterial cultures were grown in LB broth at 37°C with aeration unless otherwise described. The derivative of phage P22 used in these studies was created by selecting transposon mutants that exhibited lysogeny with high frequency. A S. Typhimurium strain was constructed harboring two plasmids: pNK2887 has a defective Tn10 derivative carrying kanamycin resistance and an outward-facing IPTG-inducible promoter (26). Its transposase is encoded on the plasmid, but not within the transposon, and is also inducible with IPTG. Thus, transposition can be induced, but once it occurs, it results in a stable transposon. pMS421 encodes lacI^q^ for overproduction of lac repressor (39). This strain was grown with 1 mM IPTG to induce transposition, wild-type P22 was added, and the strain was grown with aeration overnight. A phage lysate was prepared by the addition of chloroform and was used to infect wild-type S. Typhimurium. Strains harboring lysogens were selected by growth on LB agar with 50 μg/mL kanamycin. Phage with efficient lysogeny was identified by introducing lysogenic phage into strains with null mutations of katE and rpoS and inducing lytic growth with hydrogen peroxide (2.5 mM) overnight, harvesting the resulting phage lysate with chloroform, and transducing wild-type S. Typhimurium, comparing kanamycin-resistant colonies to plaque-forming units. rpfF was integrated into the phage genome under the control of the P2 promoter, reported to provide moderate expression (Addgene) (40), along with an associated chloramphenicol-resistance gene. These genes were introduced into the gtrABC operon of phage P22 using the one-step inactivation method as previously described (41).
Fatty acid extraction
Strains were cultured in LB broth overnight, and fatty acids produced by the bacterial strains were extracted using chloroform. Bacterial supernatants were mixed with two volumes of chloroform and mixed by rotation for 20 min. The chloroform phase was carefully decanted into a glass container. Extraction was repeated twice and the extracts pooled. Chloroform was evaporated, and the fatty acids extracted were air-dried. Extracts were derivatized and analyzed by GC-MS or dissolved in ethanol for treating bacterial cultures.
Identification of signal
For signal identification, extracted fatty acids were derivatized overnight with a 1:1 mixture of hexane and 3-(trifluoromethyl) phenyltrimethylammonium hydroxide (TCI chemicals). A volume of 0.05 mL extract in ethanol was derivatized in a volume of 0.2 mL of the derivatizing agent. Derivatized samples were separated on an Agilent 19,091B-102 Ultra 2 35 m × 200 µm × 0.33 µm column mounted on a Thermo GC-Orbitrap system coupled to a mass spectrometer. Commercially acquired c2HDA was used as a standard for retention time analysis and mass fragmentation.
Luciferase assays
Strains carrying luxCDABE reporter fusions on a plasmid were grown overnight in LB with the necessary antibiotics. Overnight cultures were diluted 100-fold into M9 minimal medium with glucose, antibiotics, and 1 mM nonanoic acid (added to repress SPI invasion gene expression to eliminate background luminescence) and grown overnight. Bacteria were inoculated at a starting optical density at 600 nm (OD_600_) of 0.02 into 150 µL of LB containing 100 mM morpholinepropanesulfonic acid (pH 6.7). Luminescence was measured in a BioTek Synergy H1 microplate reader at 37°C for 24 h.
qRT-PCR
Lysogens were cultured in LB broth as above for 4 h. RNA was extracted using the TRIzol reagent according to the manufacturer’s instructions. Briefly, 0.25 mL of bacterial sample was mixed with 1 mL of TRIzol. The sample was then lysed by bead-beating in 2 mL glass bead-preloaded microtubes (Omni International). Samples were centrifuged to separate the beads. The supernatant was mixed with 0.2 mL of chloroform, incubated for 3 min, and then centrifuged. The RNA sample was precipitated using isopropanol and washed twice with 75% ethanol. The dried sample was reconstituted in RNase-free water. qPCR was performed using the iTaq Universal SYBR Green One-step kit (Bio-Rad) according to the manufacturer’s instructions. For each sample, 100 ng of RNA was used, and experiments were performed with three technical replicates. Reverse transcription was performed at 50°C for 10 min, and the cDNA was initially denatured for 5 minutes at 95°C. Quantitative PCR was performed using specific primers for the genes to be tested. Results were normalized by comparing them with the housekeeping gene rpoD. All tests were performed in triplicate.
Flow cytometry
Strains carrying sicA-GFP and additionally constitutively expressing BFP were cultured overnight in LB broth. Bacterial cultures were pelleted and resuspended in 4% paraformaldehyde in PBS and fixed for 30 min at 4°C. Flow cytometry was performed as previously described (31). Recovered bacteria were analyzed for blue fluorescent (BFP) and green fluorescence (GFP) using an Attune analyzer NxT flow cytometer (Thermo Fisher). Salmonella was identified by BFP expression, and GFP was used to monitor invasion-gene expression.
Invasion assay
Invasion was determined using the gentamicin-protection assay as previously described (9). Strains to be tested were cultured overnight in LB. Bacteria were washed with PBS, and ~2 × 10^6^ were added to HEp-2 cells to maintain a multiplicity of infection of 10. Plates were centrifuged for 10 min at 100 × g and incubated for 1 hour at 37°C. Plates were then washed, and gentamicin was added at a concentration of 20 μg/mL. After 1 h of incubation, cells were washed and lysed with 1% Triton X-100. Lysates were plated for enumeration of recovered intracellular bacteria. Percentage invasion of the strains was calculated by comparing the number of bacteria inoculated with that of bacteria recovered.
Animal experiments
The 129X1/SvJ mouse model was used for testing the activity of lysogenized phage in the gut. This is a well-established mouse model that is susceptible to Salmonella colonization without the use of antibiotics and has been used for long-term infections (32). Mice 6–8 weeks old were acquired from the Jackson Laboratories. A total of 1 × 10^8^ plaque-forming units of extracted phage were administered in 100 µL by oral gavage a day before Salmonella infection and every other day after infection. Mice were inoculated with a total of 1 × 10^8^ CFU of Salmonella strains in 100 µL PBS. Fecal pellets were collected for plating to assess colonization, and inflammation was tested by lipocalin-2 assay (R&D systems) according to manufacturer’s guidelines.
Statistical analysis
Results are presented as means with standard deviations. Comparisons between controls and tests were evaluated with GraphPad Prism 10 using the Student’s t-test.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Prouty AM, Gunn JS. 2000. Salmonella enterica serovar typhimurium invasion is repressed in the presence of bile. Infect Immun 68:6763–6769. doi:10.1128/IAI.68.12.6763-6769.200011083793 PMC 97778 · doi ↗ · pubmed ↗
- 2Eade CR, Hung CC, Bullard B, Gonzalez-Escobedo G, Gunn JS, Altier C. 2016. Bile acids function synergistically to repress invasion gene expression in Salmonella by destabilizing the invasion regulator Hil D. Infect Immun 84:2198–2208. doi:10.1128/IAI.00177-1627185788 PMC 4962646 · doi ↗ · pubmed ↗
- 3Garner CD, Antonopoulos DA, Wagner B, Duhamel GE, Keresztes I, Ross DA, Young VB, Altier C. 2009. Perturbation of the small intestine microbial ecology by streptomycin alters pathology in a Salmonella enterica serovar Typhimurium murine model of infection. Infect Immun 77:2691–2702. doi:10.1128/IAI.01570-0819433544 PMC 2708583 · doi ↗ · pubmed ↗
- 4Huang Y, Suyemoto M, Garner CD, Cicconi KM, Altier C. 2008. Formate acts as a diffusible signal to induce Salmonella invasion. J Bacteriol 190:4233–4241. doi:10.1128/JB.00205-0818424519 PMC 2446767 · doi ↗ · pubmed ↗
- 5Chowdhury R, Pavinski Bitar PD, Chapman HM, Altier C. 2023. Salmonella invasion is controlled by competition among intestinal chemical signals. m Bio 14:e 0001223. doi:10.1128/mbio.00012-2337017539 PMC 10127606 · doi ↗ · pubmed ↗
- 6Golubeva YA, Ellermeier JR, Cott Chubiz JE, Slauch JM. 2016. Intestinal long-chain fatty acids act as a direct signal to modulate expression of the Salmonella pathogenicity Island 1 type III secretion system. m Bio 7:e 02170-15. doi:10.1128/m Bio.02170-1526884427 PMC 4752608 · doi ↗ · pubmed ↗
- 7Hung C-C, Garner CD, Slauch JM, Dwyer ZW, Lawhon SD, Frye JG, Mc Clelland M, Ahmer BMM, Altier C. 2013. The intestinal fatty acid propionate inhibits Salmonella invasion through the post-translational control of Hil D. Mol Microbiol 87:1045–1060. doi:10.1111/mmi.1214923289537 PMC 3581741 · doi ↗ · pubmed ↗
- 8Diard M, Garcia V, Maier L, Remus-Emsermann MNP, Regoes RR, Ackermann M, Hardt W-D. 2013. Stabilization of cooperative virulence by the expression of an avirulent phenotype. Nature 494:353–356. doi:10.1038/nature 1191323426324 · doi ↗ · pubmed ↗