Population differences in reproductive resource allocation and heterosis in the invasive vector Aedes albopictus
Ayda Khorramnejad, Claudia Alfaro, Stefano Quaranta, Alejandro Nabor Lozada-Chávez, Laila Gasmi, Hugo D. Perdomo, Laurent Roberto Chiarelli, Mariangela Bonizzoni

TL;DR
This study finds that invasive populations of Aedes albopictus mosquitoes have higher reproductive capacity and hybrid vigor, which may help explain their success in spreading.
Contribution
The study reveals population-level physiological and genetic differences in reproductive traits linked to invasion success in Aedes albopictus.
Findings
Invasive Aedes albopictus populations are larger and have higher reproductive output than native and old populations.
Hybrid vigor is observed in invasive populations, though it varies across different groups.
Reproductive advantages in invasive mosquitoes are supported by both physiological and genetic factors.
Abstract
An understanding of the traits that favour biological invasions has been considered to be an essential step in predicting which species would become successful invaders. Classical approaches test for differences between invasive versus non-invasive species and emphasize reproduction as a critical phenotype for successful establishment of an invasive species. However, cross-species comparisons underestimate intra-species differences, which may be relevant in species with complex invasion histories. We capitalize on the well-characterized invasion history of the arboviral vector Aedes albopictus, which has resulted in genetically distinct native, old and invasive populations, and compared the reproductive capacity (fertility and fecundity), development (timing of egg hatching, oviposition patterns and egg hatching) and physiology (blood digestion and nutrient movement during oogenesis)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
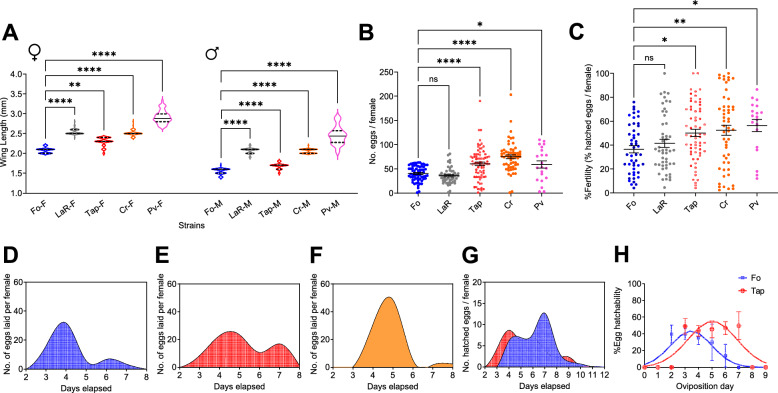 Figure 1
Figure 1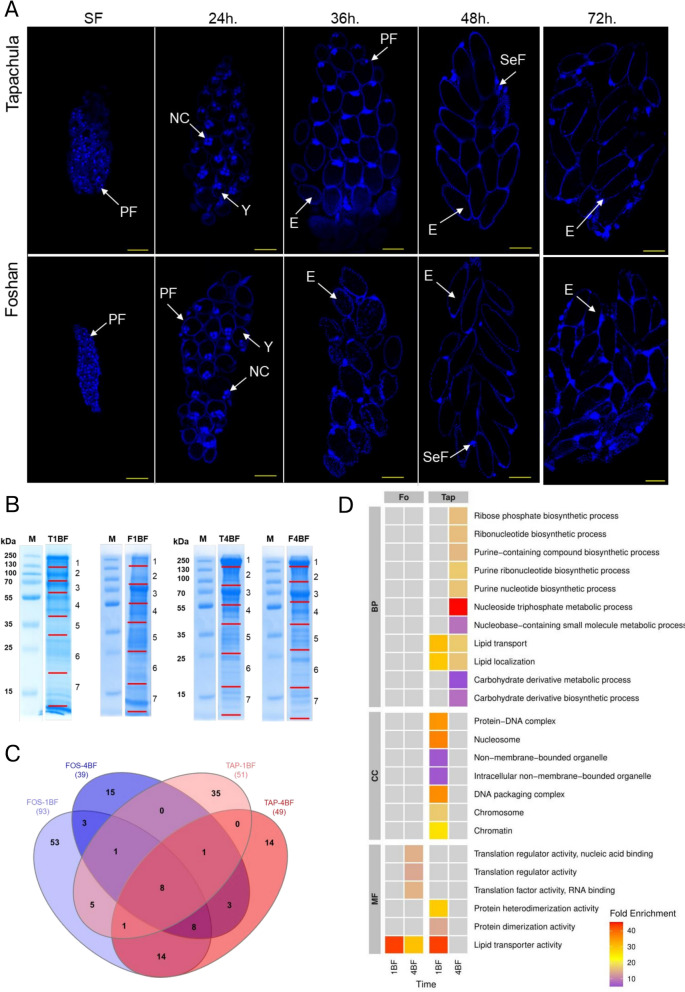 Figure 2
Figure 2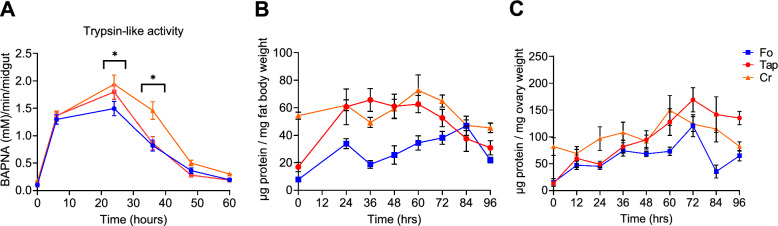 Figure 3
Figure 3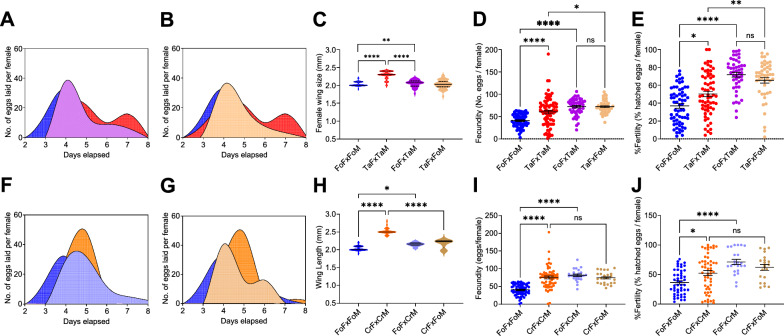 Figure 4
Figure 4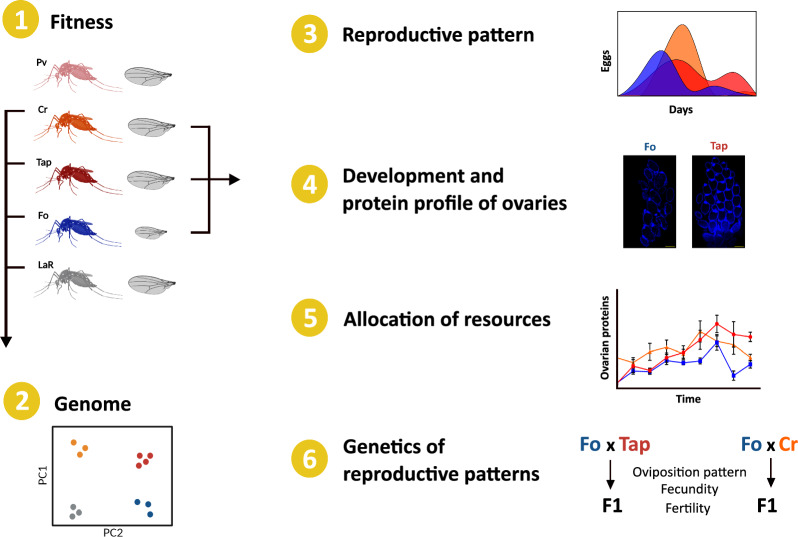 Figure 5
Figure 5- —EU funding within the NextGeneration EU-MUR PNRR Extended Partnership initiative on Emerging Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Malaria Research and Control · Neurobiology and Insect Physiology Research
Background
Biological invasions are complex and multifaceted processes that have been recognized since the late 1950 s as a rapidly increasing phenomenon with daunting ecological, sociological and economic consequences [1]. Therefore, understanding the key traits that support a species invasion has been considered to be an important step for the prediction and the management of invasive species [2]. A typical approach is the comparison of species with different invasion success [3–5]. Experimental data obtained from many taxa using a cross-species comparison approach highlight the crucial role of reproduction in the establishment of a species in a new area, following its introduction and preceding further spread [3]. For example, invasive pine species tend to have a small seed mass, which is indicative, in these plants, of a larger seed number, high germinability and higher relative growth rate of seedlings than non-invasive pine species [6]. Data from invasive insects, birds and vertebrate species correspond with life history traits of invasive plants, with successful invasions associated with a high reproductive capacity and short generation time of the invasive species [4, 7–9]. However, traditional approaches that rely on comparisons between invasive and not-invasive organisms tend to neglect intra-species variation, which could be significant in species with highly differentiated populations and/or complex invasion histories.
The Asian tiger mosquito Aedes albopictus (Skuse, 1894) is the archetype of a successful invasive species with a complex invasion process. This mosquito, which is native to Asia, colonized islands of the Indian Ocean and the Pacific, such as Hawaii and La Reunion Island, during the spice trade in the seventeenth–eighteenth century [10]. Through the global increases in commerce and human travel during the past century, Ae. albopictus moved globally and has currently established populations on every continent, with the exception of Antarctica [11]. Based on this dual wave of colonization, current Ae. albopictus populations are called native, old or invasive depending on their origin in Asia, the Indian Ocean and the Pacific or elsewhere in the world [11, 12]. Genetic data from studies performed in the last decades consistently showed that this classification also reflects population genetic differences and that invasive populations often derive from overlaying introductions of mosquitoes of various origins [12–14]. However, it has yet to be empirically tested whether populations with diverse invasion histories differ in their reproductive capacity. The fact that the invasion process of Ae. albopictus was mediated by the chaotic distribution of propagules through human activities further suggests genetic mixing among mosquitoes from overlaying introductions. Heterosis and genetic admixture have already been shown to foster the invasion success of multiple plant species, including the highly invasive seep monkeyflower Mimmulus gattatus [15, 16] and the common ragweed Ambrosia artemisiifolia L [17]. Whether these factors also apply to insect species remains uncertain, with contrasting experimental evidence [18–20].
In the study reported here, we first compared reproductive capacity across geographic populations of Ae. albopictus and then used physiological, microscopic and proteomics approaches to investigate mechanisms underpinning observed differences in their reproductive capacity, with a focus on invasive populations. In addition, reciprocal crosses were performed between invasive populations and mosquitoes from the Ae. albopictus Foshan (Fo) reference strain to test for heterosis.
Methods
Mosquitoes
For this study, we used mosquitoes from the long-established laboratory Fo population [21], together with mosquitoes adapted to the laboratory from eggs collected between 2017 and 2022 from La Reunion Island (LaR), Tapachula, Mexico (Tap), Crema, Italy (Cr) and Pavia, Italy (Pv) [21, 22]. Briefly, eggs were collected from multiple ovitraps across each site or from adults [21, 22]. For the Pv laboratory-adapted population, adult mosquitoes were sampled in the summer of 2022 in sufficient numbers to generate at least 100 adults for the G1 laboratory generation. All five populations were raised in Binder KBWF climatic chambers (Binder GmbH, Tuttlingen, Germany) under constant conditions (28 ± 1 °C; 70–80% relative humidity; photoperiod 12 h:12 h light:dark). The larvae were reared in plastic containers measuring 19 × 19 × 6 cm (BugDorm, MegaView Science Co., Taichung, Taiwan) at a controlled density (approx. 200 larvae in 1 L of water) to prevent competition. Food daily was provided in the form of fish food (Tetra Goldfish Gold Colour; Tetra Werke, Melle, Germany). Adults were kept in 30-cm^3^ cages and fed using cotton soaked with 20% sugar. Adult females were fed commercial defibrinated mutton blood (Biolife Italiana, Monza, Italy) using a Hemotek blood membrane feeding system.
Fitness assessment
Fitness evaluation was carried out within the first 10 generations after colony establishment for the Tap, Cr, Pv and LaR mosquito populations. Given the differences in the collection time among these populations, fitness assessments were conducted using the same conditions and protocols—but at different times—for each field-derived population, but with each fitness assessment carried out in parallel with fitness assessment of Fo mosquitoes. For each population, three replicates of 100 eggs each were hatched in plastic containers (17 × 6.5 × 12 cm) containing 200 ml of water and reared under the standard conditions described in section Mosquitoes. The larval to pupal developmental time was recorded until pupation. Emerging adults were sexed. In parallel, six more replicates of 100 eggs were hatched for each population in 200 ml of water for further measurements, and samples of 50 females and 50 males was collected for wing length measurement as a proxy for body size, as previously described [22]. For each population, a total of 200 females and 200 males were separated at emergence, and the survival of each individual was monitored until death. Oviposition pattern, fecundity and fertility were also monitored. Briefly, at least fifty 3- to 5-day-old females were placed in cages with 50 males to mate. After 7 days, females were offered a blood meal, following which only fully engorged females were collected, with each individual placed in a separate circular cup. We provided a damp filter paper for egg deposition 2 days after the blood meal and monitored the number of eggs laid by each female for the following 6 days to determine the patterns of oviposition. We also measured the total number of eggs laid per female (fecundity). Each filter paper was then dried for 2 days and subsequently placed in water for egg hatching; the percentage of hatched eggs per female (fertility) was recorded. In parallel, we took a minimum of thirty 3- to 5-day-old mated females per population and weighed them on a microbalance (Mettler AC100; Mettler-Toledo GmbH, Giessen, Germany) before and after the blood meal; the weight difference corresponded to the amount of blood ingested (blood intake). Reproductive fitness assessment for Fo strain mosquitoes showed temporal stability (Additional file 1: Figure S1).
The Prism 9.1 software program (GraphPad Software, San Diego, CA, USA) was used for statistical analyses of life history traits. We verified the normality of each dataset with the Shapiro–Wilk test and proceeded accordingly. Differences in wing length, fecundity and fertility among the different laboratory-adapted populations were tested using one-way analysis of variance (ANOVA), followed by Tukey’s multiple comparison [23]. We compared larval to pupal developmental time, sex ratio, wing size, weight and blood intake using the Student t-test [23] and tested differences in longevity using the Log-rank (Mantel Cox) analysis [23]. Oviposition patterns were compared using a cubic spline analysis [23].
Reciprocal crosses
We performed reciprocal crosses between mosquitoes from the Tap and Fo populations and from the Cr and Fo populations, respectively, by exposing 20 newly emerged females from one population to seven newly emerged males from the other population for 7 days; we then provided a blood meal and provided a damp filter paper for egg deposition. The F_1_ eggs were hatched and the progenies were interbred. Wing length, fecundity and fertility of the F_1_ mosquitoes were determined as described in section Fitness assessment. Wing length, fecundity and fertility were compared among the progeny of each cross and also with the parental Tap, Cr, and Fo populations, using a Mann–Whitney test in Prism 9.1 software (GraphPad Software) because the data were not parametric.
Protein and lipid content
A total of thirty-six 3- to 5-day-old female mosquitoes were assessed for protein and lipid content before the blood meal and at 24 and 96 h post blood meal (hpBM). The fat body of each individual mosquito was dissected in 75% ethanol and then weighed using a microbalance (Mettler AC100; Mettler-Toledo GmbH), following which the samples were homogenized individually in 180 μl of lysis buffer (100 mM KH_2_PO_4_ [Sigma-Aldrich, St. Louis, MO, USA], 1 mM DTT, 1 mM EDTA [Thermo Fisher Scientific, Waltham, MA, USA], pH 7.4) with pestles. The samples were stored at − 20 °C until processing to quantify the protein and lipid content using a colorimetric assay as previously described [22]. Following this same procedure, wer determined the protein content of the fat body and ovaries of 216 females that were sampled at 12, 24, 48, 60 and 72 hpBM. We compared protein time and lipid course using two-way ANOVA tests followed by Sidak’s multiple comparison tests using Prism 9.1 (GraphPad Software). Measurements of energy reserves were conducted in parallel for all tested populations.
Midgut trypsin-like activity
For the measurement of midgut trypsin-like activity, we provided a blood meal to 3- to 5-day-old females and subsequently sampled fully engorged females at 6, 24, 36, 48 and 60 hpBM; a sample of 3- to 5-day-old females (n = 144) were also sampled before the blood meal. The midguts of each mosquito were dissected on ice and trypsin-like activity subsequently measured as described previously [24], with slight modifications. Briefly, we homogenized each sample in 100 μl of an extraction buffer (20 mM Tris/20 mM CaCl_2_ [Sigma-Aldrich], pH 8) using a pestle, followed by centrifugation for 2 min at 14000* g*, 4 °C. The supernatants were collected and stored at − 80 °C until measurements were performed. For sugar-fed samples and samples collected 6 hpBM we used aliquots of 10 μl; for samples from all other time points we used aliquots of 5 μl and added these to 100 μl of 4 mM Nα Benzoyl-L- Arginine-p-Nitroanilide (BApNA; Sigma-Aldrich) in wells of a 96-well plate. The plates were incubated at 37 °C for 10 min, followed by an absorbance reading at 405 nm in a CLARIOstar plate reader (BMG Labtech, Ortenberg, Germany). Trypsin-like activity was quantified using a standard absorbance curve generated using 20 μg of bovine serum trypsin (Sigma-Aldrich) with BApNA standards (8.96, 4.48, 2.24, 1.12, 0.56, 0.28 and 0.14 mM).
Microscopic analysis of mosquito ovaries
A blood meal was provided to 3- to 5-day-old female mosquitoes, and fully engorged females were subsequently samples in groups of five at 24, 30, 36, 42, 48, 72 and 96 hpBM. The ovaries of each female were dissected in 75% ethanol, following which the tissues were fixed in a Canois solution (60% ethanol [Sigma-Aldrich], 30% chloroform [Sigma-Aldrich] and 10% acetic acid [Sigma-Aldrich]) for up to 1 week. The samples were then rinsed 3 times with 1 × phosphate buffered saline (PBS) and permeabilized with permeabilization solution (0.2% Triton [Sigma-Aldrich], 1 × PBS and 1% bovine serum albumin [BSA; Promega, Madison, WI, USA]) for 1 h. As a final step, the ovaries were stained with 4′-, 6-diamidino-2-phenylindole (DAPI [Thermo Fisher Scientific], diluted 1:50 in 1 × PBS) for 5 min before mounting the samples on the slide using polyvinyl alcohol medium (Sigma-Aldrich). Images were taken with a TCS Sp8 confocal microscope (Leica Microsystems, Wetzlar, Germany) at Centro Grandi Strumenti at the University of Pavia (https://cgs.unipv.it/?page_id=84). The complete set of images can be viewed at 10.6084/m9.figshare.30784160.
Proteomic analyses of ovaries
A blood meal was provided to 3- to 5-day-old female mosquitoes, and either 1 or 5 days later, we sampled 100 fully engorged females to dissect ovaries in 75% ethanol under a dissection stereoscope (Leica ZOOM 2000; Leica Microsystems). Ovaries from females of the same population were pooled and collected at the same time point post blood meal for protein extraction following Geiser et al. [25], with some modifications. Briefly, tissues were homogenized in 500 μl of lysis buffer (7 M urea, 2 M thiourea, 50 mM HEPES pH 8, 75 mM NaCl, 1 mM EDTA and a cocktail of 1 × Halt proteases inhibitors) with a pestle, and then step-wise incubated for 30 min on ice, sonicated using a probe in ice and centrifuged at 12,000 rpm for 10 min at 4 °C. For the quantification of protein content, we collected the supernatant and performed the Bradford assay [26]. Samples were then subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by an in-gel digestion. Briefly, 30 μg of proteins of each sample were loaded onto an SDS-PAGE gel (stacking 5%, resolving 12%) and proteins were separated in an electrophoretic run at 200 V for 40 min. The resulting profile was divided into seven fractions, and the gel was cut accordingly. Each gel piece was destained by covering it with a sufficient volume of a 100 mM ammonium bicarbonate + 50% acetonitrile solution for 15 min. This washing step was repeated until the gel pieces were completely destained. The gel pieces were then dehydrated in pure acetonitrile and incubated at room temperature; the acetonitrile-treated gel pieces were later dried at 60 °C for 5 min. The gel pieces were reduced in a freshly prepared 10 mM dithiothreitol (DTT) solution, with incubation at 37 °C for 30 min, followed by alkylation with freshly prepared 55 mM iodoacetamide (Sigma-Aldrich) and incubation for 45 min at 60 °C. These samples were washed twice with 200 μl of 100 mM ammonium bicarbonate before a second dehydration with 200 μl of pure acetonitrile. We then performed protein digestion by incubating samples on ice for 30 min with 50 μl of 20 ng/μl trypsin in 100 mM ammonium bicarbonate, followed by overnight incubation at 37 °C with sufficient 100 mM ammonium bicarbonate to avoid gel drying. The following day, each gel piece was treated with 100 μ of a 50% acetonitrile/5% formic acid solution for 15 min at 37 °C to extract proteins. The proteins of fragments coming from the same gel fragment were pooled and dried in a Speed Vac® without exceeding 30 °C for 4 h. Peptides were resuspended in 20 μl of distilled water and 1 μl of formic acid and stored at − 20 °C until mass spectrometry analysis.
The proteomic profiles of each sample were obtained via liquid chromatography–mass spectrometry (LC/MS) performed at the Centro Grandi Strumenti of University of Pavia (https://cgs.unipv.it/?page_id=96) using the LC/MS–MS LCQ Fleet instrument (Thermo Fisher Scientific). Mass spectra were generated using SCIEX software (Sciex) and proteins were identified using a threshold of at least one distinct peptide per protein with 95% confidence and a personalized database, which comprises all Ae. albopictus proteins (AalbF2 assembly) and Aedes aegypti proteins (AaegL5.3 assembly) as obtained from VectorBase (https://vectorbase.org/vectorbase/app).
Functional gene annotation and enrichment
Gene Ontology (GO) functional assignment and gene enrichment of protein-coding genes were done following Lozada-Chávez et al. [27]. This process allowed the annotation of 69% of the Ae. albopictus proteome. Our custom annotation database was used for GO enrichment analysis with clusterProfiler v4.2.2 [28] to identify functional groups that were enriched in proteins identified through the LC/MS analyses of the ovaries. P-values (P ≤ 0.05) obtained with clusterProfiler were corrected for multiple tests with the Benjamini–Hochberg procedure, and redundant enriched GO terms in each major GO classification were removed with clusterProfiler (‘simplify’ function).
Genomic analyses
Whole-genome sequencing (WGS) data from pools of 40 mosquitoes/population were used for the genomic analyses. Raw sequencing reads are available under the NIH BioProject number PRJNA1080321. We evaluated the quality of raw sequencing reads with FastQC (v. 0.12.1) [29], and trimmed adapters using fastp (v. 0.23.2) [30]. The resulting reads were aligned with the Ae. albopictus genome (AalbF2 assembly, VectorBase rel. 55) [31] using the bwa-mem2 aligner (v. 2.2.1) [32]. The pairwise fixation index (Fst) was calculated across the genome and Tajima’s D genetic diversity (Additional file 2: Tables S1, S2) was calculated using grenedalf (v. 0.3.0) [33] with the PoPoolation methods [34]. Both metrics were computed by excluding nucleotide positions with base counts < 2 (–filter-sample-min-count 2), coverage < 5 (–filter-sample-min-coverage 5), minimum mapping quality < 20 (–sam-min-map-qual 20) and minimum base quality score < 20 (–sam-min-base-qual 20).
Results
Invasive mosquitoes are larger and have a higher reproductive output than old and long-laboratory-adapted populations
Laboratory mosquito populations were established from invasive Ae. albopictus mosquitoes sampled in Tapachula, Mexico (Tap) and Crema, Italy (Cr) and Pavia, Italy (Pv) [21, 22]. We also adapted eggs collected on the Indian Ocean Island of La Reunion (LaR) to laboratory conditions; this population is referred to here as “old” because it belongs to the first wave of Ae. albopictus colonization outside the native range [11, 12]. We measured the body size and the reproductive output (fecundity and fertility) of mosquitoes of these populations within the first 10 generations of laboratory colonization, and compared these parameters to those of mosquitoes of the Ae. albopictus reference Foshan (Fo) strain [21].
We observed that mosquitoes of both sexes derived from invasive populations, namely the Tap, Cr and Pv populations, have a wider wing length than Fo mosquitoes and a higher reproductive output compared to mosquitoes from both the old population of LaR and Fo mosquitoes, with the reproductive output not differing significantly among invasive mosquitoes (Fig. 1a–c), Additional file 2: Table S3). Using WGS data, we also verified that the tested populations are genetically different, with levels of genetic diversity and pairwise Fst values comparable to those estimated across invasive, old and native Ae. albopictus populations, albeit Fo mosquitoes have lower genetic diversity than populations recently adapted to laboratory conditions, as previously noted [12, 21, 35] (Additional file 2: Tables S1, S2).Fig. 1. Phenotypic variations among the Aedes albopictus populations tested. A Wing length of at least 50 females (F) and 50 males (M) of all populations. B Number of eggs laid per female per population. C Percentage of larvae developed from the eggs deposited by each female.** D–F **Cubic spline curves show the oviposition patterns during the first gonadotrophic cycle (GC) of Fo (D), Tap (E) and Cr (F) mosquitoes. G Cumulative pattern of egg hatching during the first GC. H Viability of each batch of eggs deposited by Fo and Tap females in relation to their deposition day during the first GC. Error bars represent the 95% confidence intervals. Asterisks indicate significant differences at *P < 0.05, **P < 0.01 and ****P < 0.0001; Ns, not significant. In all panels, data on Fo are in blue, data on Tap are in red and data on Cr are in orange. Fo, Ae. albopictus Foshan reference strain; LaR, Tap, Cr, Pv, laboratory mosquito populations established from eggs collected at La Reunion Island, Tapachula (Mexico), Crema (Italy) and Pavia (Italy), respectively
The higher reproductive capacity of invasive mosquitoes correlates with a delay in oviposition
Having established that genetically the laboratory mosquito populations we established from invasive Ae. albopictus mosquitoes (LaR, Tap, Cr, Pv) and the Fo reference strain reflect the observed distinction among native, old and invasive populations and that the fitness traits of Fo mosquitoes have been maintained constant across generations (Additional file 1: Figure S1), we checked if the higher fecundity of invasive Tap and Cr mosquitoes correlates with a different oviposition pattern with respect to that of Fo mosquitoes. We therefore counted the number of eggs deposited daily from the second to the eighth day pBM. The results revealed delayed oviposition in both Tap and Cr females with respect to Fo females, with peak oviposition occurring in the latter population between 3 and 4 days pBM (Fig. 1d). More than 95% of Fo females deposited their eggs in one or two batches, each with an average (± standard deviation [SD]) clutch size of 20.38 ± 2.99 eggs (Fig. 1d); fewer than 2% of the tested females laid > 2 clutches of eggs throughout the 6 days of our observation period. In contrast, both Tap and Cr mosquitoes had an oviposition peak between 4 and 5 days pBM (Fig. 1e, f), with an average clutch size of 60.05 ± 3.80 eggs for Tap mosquitoes and 74.88 ± 4.05 eggs for Cr mosquitoes. The Tap mosquitoes showed a second peak of oviposition at 7 day pBM; 21.12% ± 9.47% of Tap mosquitoes laid > 2 clutches of eggs, with an average clutch size of 22.80 ± 0.54 eggs (Fig. 1e). Given the presence of two oviposition peaks in Tap and Fo mosquitoes, we tested if egg hatching parallels the oviposition pattern by individually assessing the fertility of females from day 2 to day 8 pBM. We observed that the first batch of eggs deposited by Tap females hatched faster than the first batch produced by Fo females (Fig. 1g). Furthermore, the hatchability of Tap eggs remained constant independently of their oviposition day (Fig. 1h), while that of Fo eggs peaked 2 days after oviposition (38.63% ± 6.43%), then gradually declined. We also observed that while the higher reproductive output of Tap mosquitoes extends from the first to the second GC (Additional file 3: Figure S2), Tap mosquitoes started laying eggs earlier in the second GC, with an oviposition peak at 2 days pBM (Additional file 3: Figure S2).
The delay in oviposition of invasive females has physiological bases
The observed delay in the oviposition pattern of invasive versus Fo females could be a behavioural trait by which mosquitoes retain mature eggs in their ovaries, or it could result from the physiology of egg production. To discriminate between these alternatives, we combined two approaches, focusing on Tap and Fo mosquitoes based on their two-peak oviposition pattern. First, we studied egg development in maturing ovaries by acquiring micrographs at intervals between 6 and 72 hpBM, starting at 24 hpBM (Fig. 2a). Second, we studied temporal changes in ovarian proteins (Fig. 2b–d).Fig. 2. Morphology and proteomics of ovaries. A Micrographs of ovaries of female mosquitoes from the Foshan reference strain (Fo) and Tapachula (Tap) population that had been sugar-fed (SF) and subsequently sampled at 24, 36 and 48 hpBM, then stained with DAPI. Arrows point to primary follicles (PF), nurse cells (NC), yolk (Y), eggs (E) and secondary follicles (SeF). Yellow scale line: 50 µm. B Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis of ovarian proteins extracted from of ovaries of female mosquitoes of the Tap (T) and Fo (F) populations, dissected one (1) and four (4) days post blood meal (BF). The M lanes correspond to the PAGERuler Plus Prestained Protein Ladder (Thermo Fisher Scientific). Red lines mark the gel slices that were excised. C Number of proteins identified in each condition/population. D Functional enrichment of proteins from ovaries of Fo and Tap mosquitoes sampled 1 and 4 days pBF. BP, Biological processes; CC, cellular constituents; MF, molecular functions
Images of the ovaries of Fo and Tap females at 24, 30, 36, 48 and 72 hpBM were captured and analyzed in comparison to the images of ovaries of sugar-fed (SF) females (Fig. 2a). Primary follicles with little or no yolk were observed in all SF females (Fig. 2a). Primary follicles with nurse cells and an increasing amount of yolk in the ovaries were observed in both Fo and Tap females sampled at 24 and 30 hpBM; at 36 hpBM, we observed a clear difference in the images of Fo and Tap mosquitoes (Fig. 2a): mature eggs and few primary follicles were observed in the ovaries of Fo females, whereas ovarioles containing mostly primary follicles with high amounts of yolk were seen in Tap females. Mature eggs were seen in the ovaries of both populations after 48 hpBM. These results support the conclusion that the observed delay in the oviposition of Tap with respect to Fo females is caused by different dynamics of egg formation and not by egg retention in Tap mosquitoes.
We subsequently compared the protein profile of ovaries of fully-engorged blood-fed Tap and Fo mosquitoes at 1 and 4 days pBM; this time interval represents the trophic and posttrophic stages of vitellogenesis in mosquitoes, respectively [36, 37]. The quantity of total proteins extracted from the ovaries at 1 day pBM did not differ between Fo and Tap mosquitoes (mean ± SD: 47.96 ± 4.99 µg/mg in Fo and 42.08 ± 6.93 µg/mg in Tap mosquitoes), but the complexity and abundance of proteins did differ (Fig. 2b). At 4 days pBM, protein quantity was higher in the ovaries of Tap mosquitoes than in those of Fo mosquitoes (the total mean protein content was 107.72 ± 17.65 µg/mg in Fo and 163.92 ± 18.88 µg/mg in Tap; t = 2.919,* df* = 44, P-value = 0.0110; Additional file 4: Figure S3), while the SDS-PAGE profile of total ovarian proteins appeared visually similar. The Tap ovaries sampled 1 day pBM were enriched in proteins associated with carbohydrate metabolic process and proteins related to non-membrane-bounded organelles, which are associated with cell structure and motility [38] (Fig. 2c, d; Table 1; Additional file 2: Table S4). At the same time point pBM, proteins found uniquely in Fo ovaries were enriched in lipid transporter activity and proteins associated with the development of oocytes and the eggs. A total of 15 proteins (Fig. 2c; Table 1), among which the presence of vitellogenin precursor (AALF008766) is notable (Additional file 2: Table S4), were found in the ovaries of both Fo and Tap females 1 day pBM (Fig. 2c, d). At 4 days pBM, we detected proteins in Tap ovaries that were enriched in functions such as carbohydrate metabolic processes, proteolysis and lipid localization related to oocyte development [39]. At the same time point, proteins exclusive to ovaries from Fo female mosquitoes were enriched in proteins linked to translation regulatory activity, lipid transporter and proteins that mediate the late stages of oogenesis and egg activation [40]. A total of 20 proteins, some of which are involved in oogenesis (e.g. vitellogenin precursor [AALF008766]) were found in the ovaries of Fo and Tap mosquitoes at 4 days pBM (Table 1). Based on the observed delay in oviposition in Tap mosquitoes with respect to their Fo counterparts, we compared the proteins detected in the ovaries sampled at 4 days pBM in Tap and 1 day pBM in Fo females (Fig. 2c; Table 1). We found 31 proteins associated with oogenesis, carbohydrate metabolic process, protein biosynthesis and glycolytic process. Importantly, none of these 31 proteins was found in the ovaries of Tap mosquitoes sampled at 1 day pBM, except for vitellogenin-A1 precursors (AALF008766 and AAEL010434) (Additional file 2: Table S4). Collectively, proteomics data support a delay in the biochemical activity of Tap mosquitoes for oogenesis in alignment with fitness results and data from ovarian micrographs. Table 1. List of the proteins identified in the ovaries of female mosquitoes of the Foshan reference strain and Tapachula population at 1 and 4 days post blood meal, and their gene ontology based on either molecular function or biological processFoTapAccession no.NameGO (MF/BP)Accession no.NameGO (MF/BP) One day pBMAALF000045Unspecified productProtein catabolic processAALF000221Odorant receptorSensory perception of smellAALF002182Alpha-amylaseCarbohydrate metabolic processAALF000790Histone H2BNucleosome assemblyAALF002986AMP dependent ligaseCatalytic activityAALF000911Unspecified productNucleic acid bindingAALF003502Kynurenine aminotransferaseKynurenine metabolic processAALF001196C-Type Lectin (CTL)-mannose bindingCarbohydrate bindingAALF004064Unspecified productCatalytic activityAALF002028Ran-binding proteinIntracellular transportAALF007667^a,b^Elongation factor 1-alphaTranslational elongationAALF002835Histone H2ANucleosome assemblyAALF007724ATP synthase subunit betaProton motive force-driven ATP synthesisAALF005503Histone H4DNA-templated transcription initiationAALF008379^a,b^Unspecified productLipid transportAALF007667^a^Elongation factor 1-alphaTranslational elongationAALF008766^a,b^Vitellogenin-A1 precursorLipid transportAALF007713ProfilinLarval central nervous system remodelingAALF009209^a,b^Serine/threonine kinaseProtein phosphorylationAALF007792Unspecified productProtein bindingAALF010133^a,b^Elongation factor 1-alphaTranslational elongationAALF008379Unspecified productLipid transportAALF010343^b^Unspecified productCarbohydrate bindingAALF008382Serine-type enodpeptidase 2CProteolysisAALF011822Leucine-rich immune proteinProtein bindingAALF008646Histone H2BDNA bindingAALF011930Unspecified productNucleic acid bindingAALF008766^a^Vitellogenin-A1 precursorLipid transportAALF012772Tyrosine-protein kinase receptorProtein phosphorylationAALF008869Unspecified productmRNA processingAALF013670Unspecified productSignal peptide processingAALF008985Histone H2BNucleosome assemblyAALF014888Unspecified productIron-sulfur cluster assemblyAALF009114Histone H2BNucleosome assemblyAALF015638Shc transforming proteinIntracellular signal transductionAALF009200Unspecified productTransferase activityAALF016323Unspecified productMAPK cascadeAALF009209^a^Serine/threonine kinaseProtein phosphorylationAALF017232^b^Cathepsin bProteolysisAALF010133^a^Elongation factor 1-alphaTranslational elongationAALF017233^b^Unspecified productProteolysisAALF010136Histone H2ANucleosome assemblyAALF018275^b^Unspecified productProteolysisAALF010978Odorant receptorSensory perception of smellAALF018287Histone H2AChromatin organizationAALF012625Unspecified productNucleic acid bindingAALF018448Unspecified productLyase activityAALF013617Histone H2ASomatic stem cell population maintenanceAALF018477^a^Serine/threonine protein kinaseProtein phosphorylationAALF015445NephrinNAAALF019930^a,b^Unspecified productLipid transportAALF018477^a^Serine/threonine protein kinaseProtein phosphorylationAALF020589tRNA pseudouridine synthaseRNA modificationAALF019930^a^Unspecified productLipid transportAALF022027Gdp mannose-4 2C6-dehydrataseGDP-mannose metabolic processAALF022150Histone H2BProtein heterodimerization activityAALF022345Unspecified productATP bindingAALF022690Unspecified productNAAALF022346^b^Actin-1Nucleotide bindingAALF024624Histone H4DNA-templated transcription initiationAALF023299atbf1Regulation of DNA-templated transcriptionAALF024650SpectrinActin filament cappingAALF023780^b^Unspecified productTetrahydrobiopterin biosynthetic processAALF025146Glycerol kinaseGlycerol-3-phosphate metabolic processAALF026435Histone H2BNucleosome assemblyAALF025156Putative 40 s ribosomal protein s2TranslationAALF027368^a,b^Unspecified productLipid transportAALF027368Unspecified productLipid transportFoTapAccession no.NameGO (MF/BP)Accession no.NameGO (MF/BP) Four days pBMAALF000831Unspecified productProteolysisAALF000045Unspecified productProtein catabolic processAALF001196^c^C-Type Lectin (CTL)-mannose bindingCarbohydrate bindingAALF001196^c^C-Type Lectin (CTL)-mannose bindingCarbohydrate bindingAALF002028^c^Ran-binding proteinIntracellular transportAALF001261Glutathione transferaseProtein bindingAALF002713Protein regulator of cytokinesis 1 prc1Microtubule cytoskeleton organizationAALF002028^c^Ran-binding proteinIntracellular transportAALF003152Heat shock cognate 70 isoform BProtein foldingAALF002114Cell division control proteinDNA replication initiationAALF003990MannosyltransferaseGlycosyltransferase activityAALF003247Mannosyltransferase 1 2CProtein O-linked glycosylationAALF005662^c^Lethal(2)essential for life protein 2 C l2eflMetal ion bindingAALF003502Kynurenine aminotransferaseKynurenine metabolic processAALF006045Unspecified productNAAALF005085ATP-binding cassette transporterTransmembrane transportAALF006185Unspecified productNAAALF005662^c^Lethal(2)essential for life protein 2 C l2eflMetal ion bindingAALF007667^c^Elongation factor 1-alphaTranslational elongationAALF005708Unspecified productCytoskeleton organizationAALF008063Unspecified productProtein bindingAALF007572Unspecified productProtein bindingAALF008379^c^Unspecified productLipid transportAALF007667^c^Elongation factor 1-alphaTranslational elongationAALF008766^c^Vitellogenin-A1 precursorLipid transportAALF007724ATP synthase subunit betaProton motive force-driven ATP synthesisAALF009209^c^Serine/threonine kinaseProtein phosphorylationAALF008130Glycogen synthaseGlycogen biosynthetic processAALF010133^c^Elongation factor 1-alphaTranslational elongationAALF008379^c^Unspecified productLipid transportAALF010343^c^Unspecified productCarbohydrate bindingAALF008766^c^Vitellogenin-A1 precursorLipid transportAALF010627Unspecified productProtein bindingAALF009209^c^Serine/threonine_kinaseProtein phosphorylationAALF011026^c^Unspecified productNucleic acid bindingAALF009854Unspecified productProtein bindingAALF011409^c^Unspecified productProteolysisAALF010133^c^Elongation factor 1-alphaTranslational elongationAALF012426Signal recognition particle receptor alpha subunitIntracellular protein transportAALF010304Calcium-binding protein E63-1Calcium ion bindingAALF012812CalminCellular component organizationAALF010343^c^Unspecified productCarbohydrate bindingAALF012831Calcium transporting ATPase 2 (ATPase 2)Calcium ion transmembrane transportAALF011026^c^Unspecified productNucleic acid bindingAALF016841^c^Cathepsin bProteolysisAALF011409^c^Unspecified productProteolysisAALF017232^c^Cathepsin bProteolysisAALF011822Leucine-rich immune protein (Coil-less)Protein bindingAALF017233^c^Unspecified productProteolysisAALF011930Unspecified productNucleic acid bindingAALF017331AmidophosphoribosyltransferasePurine nucleotide biosynthetic processAALF012374GTP:AMP phosphotransferaseITP metabolic processAALF017662OviductinProteolysisAALF013007Protein tyrosine phosphatase 69dProtein dephosphorylationAALF018200GTP-binding protein alpha subunit 2 C gnaSignal transductionAALF013502Translation initiation factorCellular metabolic processAALF018275^c^Unspecified productProteolysisAALF016841^c^Cathepsin bProteolysisAALF018689Unspecified productNitrogen compound metabolic processAALF017233^c^Unspecified productProteolysisAALF018796DNA polymerase vRegulation of DNA transcriptionAALF017772Unspecified productNAAALF019930^c^Unspecified productLipid transportAALF018275^c^Unspecified productProteolysisAALF020554^c^ActinNucleotide bindingAALF018477Non-specific serine/threonine protein kinaseProtein phosphorylationAALF020972LipaseLipid metabolic processAALF019930^c^Unspecified productLipid transportAALF021555Unspecified productATP bindingAALF020554^c^ActinNucleotide bindingAALF022346^c^Actin-1Nucleotide bindingAALF020589tRNA pseudouridine synthaseRNA modificationAALF022559Alcohol dehydrogenaseObsolete oxidation–reduction processAALF022027Gdp mannose-4 2C6-dehydrataseGDP-mannose metabolic processAALF022690^c^Unspecified productNAAALF022345Unspecified productATP bindingAALF023780^c^Unspecified productTetrahydrobiopterin biosynthetic processAALF022346^c^Actin-1Nucleotide bindingAALF024081^c^CarboxypeptidaseProteolysisAALF022690^c^Unspecified productNAAALF025260OviductinProteolysisAALF023780^c^Unspecified productTetrahydrobiopterin biosynthetic processAALF025991Histone-lysine n-methyltransferaseHistone lysine methylationAALF024081^c^CarboxypeptidaseProteolysisAALF026483Unspecified productEukaryotic translation initiationAALF025494Glyceraldehyde-3-phosphate dehydrogenaseGlycolytic processAALF026901Sdk-P1Anatomical structure morphogenesisAALF026319Tropomyosin invertebrateNAAALF027134Unspecified productProtein bindingAALF026435Histone H2BNucleosome assemblyAALF027368^c^Unspecified productLipid transportAALF027368^c^Unspecified productLipid transportProteins listed in table were identified by liquid chromatography–tandem mass spectrometry analysisBP Biological process,* Fo* Foshan reference strain,* GO* Gene Ontology,* MF* molecular function,* NA* not available,* pBM* post blood meal, * Tap* Tapachula (Mexico) population^a^Proteins found in common between Fo and Tap at 1 day pBM ^b^Proteins in common between Fo at 1 day pBM and Tap at 4 days pMB^c^Proteins found in common between Fo and Tap at 4 days pBM
Invasive mosquitoes efficiently allocate nutritional resources during oogenesis
To further probe the physiological basis of the observed delay in oviposition of invasive versus Fo females, we quantified trypsin-like enzyme activity after a blood meal, as a proxy for blood protein digestion [36]. We also measured and compared the accumulation and depletion of energy reserves in the fat body and their absorption in the ovaries (Fig. 3) because after acquisition and digestion of a blood meal, yolk precursor proteins are produced in the fat body, secreted into the hemolymph and transported to oocytes for egg production [37].Fig. 3. Oviposition-related metabolism. A Trypsin-like activity in female before blood meal (time 0) and at 10, 20, 30, 40, 50, and 60 h post blood meal (hpBM). Trypsin-like activity was detected using BAPNA, a chromogenic substrate. ** b**,** c** Protein content in the fat body (B) and ovaries (C) of females before the blood meal (time 0) and at 24, 36, 48, 60, 72, 84, and 96 hpBM. Asterisk indicates a significant difference at P < 0.05
Mosquitoes aged between 3 and 5 days were weighed and then provided with a blood meal; after mosquitoes had flown to a resting spot, we weighed them again to avoid having the quantification of blood intake biased by diuresis (Additional file 3: Figure S2). The protease activity of trypsin-like enzymes showed similar trends in Fo, Tap and Cr females (Fig. 3a). Protease activity was detected from 6 to 60 hpBM, with a peak at 24 hpBM in all tested populations, but a significantly higher trypsin-like activity was found in invasive versus Fo mosquitoes at 24 hpBM (Fig. 3a). These results suggest that invasive mosquitoes process a blood meal more efficiently than those of the Fo reference population. To confirm this result, we tracked the mobilization of proteins from the fat body to ovaries post blood meal at 12-h intervals for up to 4 days pBM (Fig. 3 b, c). Similar patterns were observed in Tap and Cr females, with protein accumulation in the fat body gradually increasing up to 60–72 hpBM, then gradually decreasing, and reaching its minimum level at 96 hpBM. In contrast, protein content in the fat body of Fo females decreased at 36 hpBM and then gradually increased, reaching its maximum level at 84 hpBM, the once again declining at 96 hpBM (Fig. 3b). Protein mobilization from the fat body to the ovary showed a similar pattern across populations, with a gradual increase and a peak at 60 hpBM in Cr mosquitoes and at 72 hpBM in Fo and Tap mosquitoes, followed by a marked decrease only in Fo mosquitoes at 84 hpBM (Fig. 3c); these results are in agreement with an earlier start of oviposition (Fig. 1d). Monitoring of lipids revealed that there was a gradual accumulation of lipids in the ovaries of Tap and Fo mosquitoes, followed by a decrease at 60 or 72 hpBM in Fo or Tap mosquitoes, respectively (Additional file 4: Figure S3).
Variation in levels of heterosis
To test for heterosis, we established reciprocal crosses between Tap and Fo and between Cr and Fo mosquitoes. We measured the wing length, fecundity and fertility of F1 females of all reciprocal crosses and monitored their oviposition patterns, and compared these to these traits of the respective parents (Fig. 4). In the matings between Tap and Fo mosquitoes, the F1 progeny showed higher fecundity and fertility than both parental strains, supporting heterosis (Fig. 4d, e; Additional File 2: Table S5). We also observed that the percentage of sterile females was significantly lower in the F1 progeny of crosses between Fo females and Tap males in comparison with the crosses of parental Fo, but not parental Tap mosquitoes (F = 6.538, P = 0.0152) (Additional file 5: Figure S4). The same trend was observed for egg viability (Additional file 5: Figure S4), suggesting a contribution of Tap males to modulating the reproductive output of Fo females. In the matings between the Cr and Fo mosquitoes, the F1 progeny showed a higher reproductive output than the parental Fo, but not significantly different than that of the invasive Cr mosquitoes (Fig. 4h–l). In both crosses, the wing length and the oviposition patterns of the F1 progeny showed intermediate phenotypes compared to those of the parental populations, suggesting Mendelian inheritance of quantitative traits (Fig. 4).Fig. 4. Varying levels of heterosis in crosses with invasive mosquitoes. A Oviposition patterns of F1 progenies of crosses between females of the Foshan reference strain (Fo) and males of the Tapachula population (Tap), in comparison with that of the parental populations. B Oviposition patterns of F1 progenies of crosses between Fo males and Tap females in comparison to that of the parental populations. C–E** Wing size (C) fecundity (D) and fertility (E) of F1 progeny of crosses between Fo females and Tap males or Fo males and Tap females, in comparison to that of the parental populations. F Oviposition patterns of F1 progenies of crosses between Fo females with Crema (Cr) males with respect to that of parental populations. G Oviposition patterns of F1 progenies of crosses between Cr males with Fo females in comparison to that of parental populations. H–J Wing size (H), fecundity (I) and fertility (J) of F1 progeny of crosses between Fo females and Cr males or Fo males and Cr females in comparison with that of parental populations. Asterisks indicate a significant difference at *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001; ns, not significant
Discussion
Reproduction is an essential process to ensure the survival of a species and a key determinant of the success of a species invasion because it facilitates the ability of a species to establish itself in a new area. In the present study, we have shifted the focus from species to populations, and we show differences in the reproductive capacity across Ae. albopictus populations, which can be associated with their history. The limited number of populations we studied and particularly the use of a single long-established laboratory colony from the native range constrain our ability to generalize our results. Observed phenotypic differences likely reflect the contribution of both population differences and laboratory adaptation. However, the observed temporal stability in fitness traits across our populations (Additional file 1: Figure S1; Fig. 4) [22], support the conclusion that their genetic background contributes to observed differences. All our invasive populations showed a consistent pattern, namely they exhibit higher reproductive output compared to mosquitoes from both the long laboratory adapted Foshan reference population and the LaR population. These repeated associations prompted us to investigate their underlying biological mechanisms. We observed that the higher reproductive output of mosquitoes from invasive populations has physiological basis: these mosquitoes optimized their nutrient investment during oogenesis, resulting in increased egg production associated with a delay in oogenesis. In Aedes spp. mosquitoes, the acquisition of a blood meal and its digestion stimulate vitellogenesis and change the metabolic activity of the fat body, resulting in the production of yolk precursor proteins, the secretion of yolk proteins and lipids, and the deposition of yolk proteins and lipids in ovaries [36, 37, 41–43]. Oviposition-related metabolism also promotes morphological and physiological changes in ovaries during oogenesis [37]. Aedes spp. females emerge with two ovaries, each divided into 50–60 ovarioles containing an oocyte, nurse cells and surrounding follicle cells forming the primary egg chamber [37]. During vitellogenesis, developing oocytes grow synchronously through the acquisition of proteins, lipids and carbohydrates to form mature eggs [37]. The oocyte of the secondary egg chamber will develop into a mature egg after a secondary blood meal [37]. Egg production is cyclical. A round of ovarian development from blood intake to egg laying is called a gonotrophic cycle. We observed that the fat body and ovaries of the invasive Tap and Cr mosquitoes had a higher protein content than those of their Fo counterparts, suggesting optimized blood meal digestion to allocate resources for egg production. In the ovaries of Tap females, lipid increase was coordinated with protein accumulation, while lipid flux and protein deposition into the developing ovaries did not follow a similar trend in Fo females. Lipids appeared later in the ovaries of Tap females than in those of Fo females, resulting in a delay in oogenesis and egg deposition. However, the more efficient acquisition of nutrients allowed females from invasive Tap and Cr populations to sustain egg production longer than Fo, resulting in higher fecundity and fertility. The observed delay in the oogenesis of Tap and Cr mosquitoes indicates that maximization of allotment in the first clutch of eggs is associated with substantial benefits. Indeed, life history theory predicts that early reproduction is more likely to be successful, and therefore more valuable, than later reproduction because the chance of mortality increases with age [44]. We also verified that the delay in oviposition of Tap females does not extend to the second GC, probably because sufficient energetic resources are available in the fat body. We cannot exclude that the shift of nutrients from the blood meal to reproduction may result in a fitness cost in terms of longevity.
Aedes albopictus moved globally in less than 60 years, with recurrent incursions favouring genetic admixture [11]. This invasion dynamics and our results showing varying degree of heterosis in crosses involving invasive mosquitoes support the hypothesis that the observed differences in the reproductive capacity across populations are adaptive. The hypothesis that Ae. albopictus can undergo rapid adaptation is supported the results of a study on the distribution of mutations predictive of pyrethroid resistance, which have appeared rapidly and independently across different populations [45]. Moreover, there are frequent examples of invasive species adapting quickly to new environments. A well-studied example is the rapid evolution of Drosophila suboscura in the New World, which was demonstrated by the detection of rapidly evolving concordant patterns of chromosomal inversions across latitudinal clines in three different continents [46]. In addition to being facilitated by intraspecific hybridization in the introduced range, which can result in heterosis and lead to novel genotypes, rapid adaptation in Ae. albopictus reproductive capacity could be induced by severe changes in the selection regime imposed by new environments [47]. As an alternative explanation for the hypothesis of rapid adaptation, Ae. albopictus invasive populations may have a high reproductive capacity because they derive from regions prone to ecological disturbance. It has been hypothesized that fluctuating environments might favour organismal flexibility or the selection for evolvability by accumulation and maintenance of genetic variation [48]. In fact, recent population genomic studies showed that the invasive and native populations of Ae. albopictus retain similar levels of genomic diversity and that invasive populations can become sources of new founder populations promoting further invasions [12, 35].
Conclusions
The observed differences in the reproductive capacity of Ae. albopictus populations in this study are underpinned by physiological adjustments and hybrid vigour. Such traits can be associated with a successful invasion by enhancing the reproductive output and persistence in novel environments. More broadly, our findings emphasize that the success of biological invasions may be influenced by intraspecific variation across diverse populations.
Supplementary Information
Additional file 1.Additional file 2.Additional file 3.Additional file 4.Additional file 5.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Manni M, Guglielmino CR, Scolari F, et al. Genetic evidence for a worldwide chaotic dispersion pattern of the arbovirus vector, Aedes albopictus. P Lo S Negl Trop Dis. 2017;11:e 0005332. 10.1371/journal.pntd.0005332.10.1371/journal.pntd.0005332 PMC 530028028135274 · doi ↗ · pubmed ↗
- 2Geiser DL, Li W, Pham DQD, et al. Shotgun and TMT-labeled proteomic analysis of the ovarian proteins of an insect vector, Aedes aegypti (Diptera: Culicidae). J Insect Sci. 2022;22:7. 10.1093/jisesa/ieac 018.10.1093/jisesa/ieac 018PMC 893250535303100 · doi ↗ · pubmed ↗
- 3Andrews S. Fast QC: A quality control tool for high throughput sequence data. 2010. https://www.bioinformatics.babraham.ac.uk/projects/fastqc/. Accessed 1 Sep 2023.
- 4Czech L, Spence JP, Expósito-Alonso M. greendalf: population genetic statistics for the next generation of pool sequencing. ar Xiv. 2024. https://arxiv.org/abs/2306.11622.10.1093/bioinformatics/btae 508PMC 1135779439185959 · doi ↗ · pubmed ↗
- 5Moreau CA, Bhargav SP, Kumar H, et al. A unique profilin-actin interface is important for malaria parasite motility. P Lo S Pathog. 2017;13:e 1006412. 10.1371/journal.ppat.1006412.10.1371/journal.ppat.1006412 PMC 546467028552953 · doi ↗ · pubmed ↗
- 6Price DP, Nagarajan V, Churbanov A, et al. The fat body transcriptomes of the yellow fever mosquito Aedes aegypti, pre- and post- blood meal. P Lo S ONE. 2011;6:e 22573. 10.1371/journal.pone.0022573.10.1371/journal.pone.0022573 PMC 314491521818341 · doi ↗ · pubmed ↗