The complete mitochondrial genome sequence of Trichoderma texanum (Hypocreales, Sordariomycetes)
Munkhgerel Dalantai, Seung-Yoon Oh

TL;DR
This paper reports the full mitochondrial genome of the fungus Trichoderma texanum, helping to understand its evolutionary relationships and genetic features.
Contribution
The study provides the first complete mitochondrial genome sequence for Trichoderma texanum.
Findings
The mitochondrial genome is 31,361 base pairs long and includes 15 protein-coding genes.
Phylogenetic analysis shows T. texanum is closely related to T. koningiopsis.
The genome includes 27 tRNA genes, two rRNA genes, and one open reading frame.
Abstract
Trichoderma texanum is a species of saprotrophic and endolichenic fungus in the genus Trichoderma. In this study, we sequenced and analyzed the complete mitochondrial genome of T. texanum strain CSC21B0438. The mitochondrial genome of T. texanum is 31,361 base pairs and comprises 15 protein-coding genes, 27 transfer RNA genes, two ribosomal RNA genes, and one open reading frames. Phylogenetic analysis using the maximum-likelihood method revealed that T. texanum is closely related to T. koningiopsis. The mitochondrial genome of T. texanum reported in this study extends the understanding of its phylogeny and genomic characteristics.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1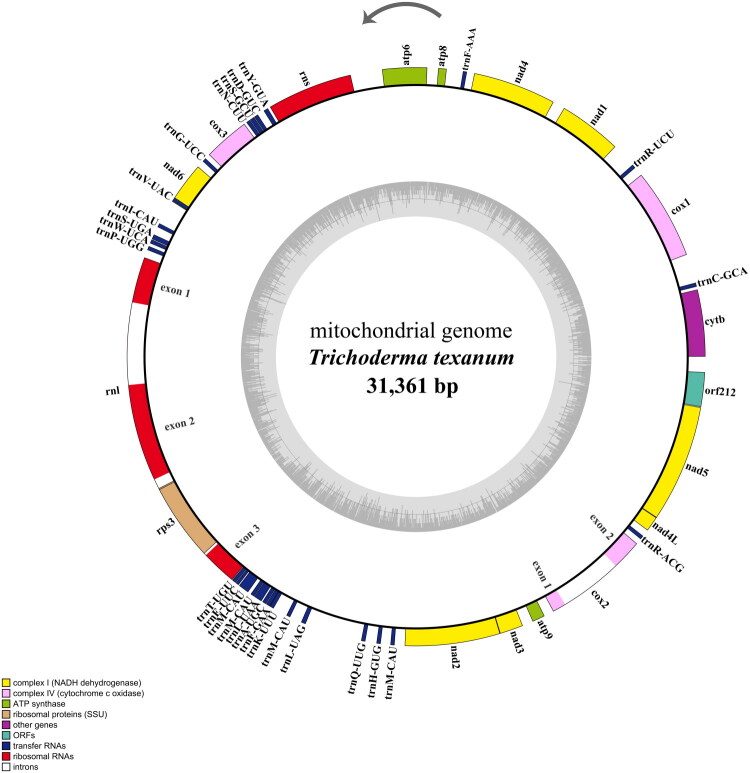 Figure 2
Figure 2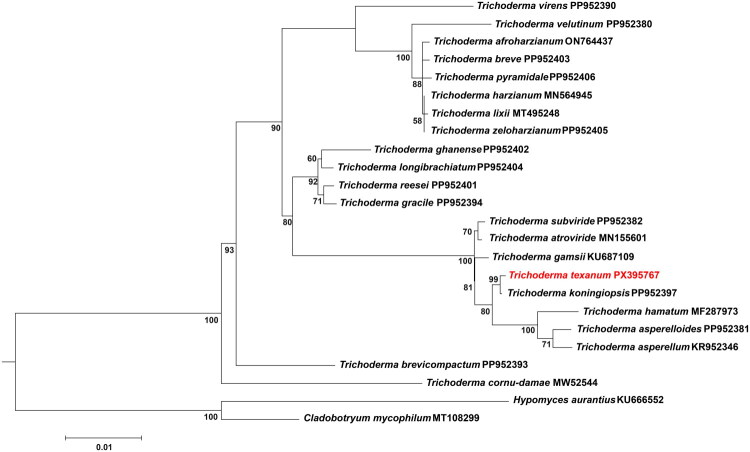 Figure 3
Figure 3- —National Research Foundation of Korea (NRF)10.13039/501100003725
- —Korea Government (Ministry of Science and ICT)
- —Korea Basic Science Institute (National Research Facilities and Equipment Center)10.13039/501100003716
- —Ministry of Education10.13039/100009122
- —‘Foundational and Protective Field of Studies Support Project’ at Changwon National University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Natural Products and Biosynthesis · Lichen and fungal ecology
Introduction
The genus Trichoderma Pers. 1794 is an ecologically important saprotrophic fungi that contributes to the degradation of organic matter in soil ecosystems and suppresses soil-borne plant diseases (Harman et al. 2004; Kubicek et al. 2011). Moreover, some Trichoderma species enhance plant growth and increase its nutrient uptake efficiency (Yao et al. 2023). Currently, the genus Trichoderma is known to have around 469 species among which 382 are currently widely distributed worldwide (Cai et al. 2022; Wang et al. 2024). Trichoderma texanum Q.V. Montoya, L.A. Meirelles, P. Chaverri & A. Rodrigues 2016 was first reported from the fungus garden of ant in Texas, U.S.A., and described as a new species based on three molecular markers (ITS, tef1 and rpb2) together with morphological characteristics (Montoya et al. 2016). T. texanum has the distinctive morphological characteristics, including globose to broadly green and smooth conidia and the elongated, relatively large chlamydospores with closely related species (Montoya et al. 2016).
According to a recent study, 47 mitochondrial genomes (mitogenomes) representing 22 species of Trichoderma have been reported (Wang et al. 2024). Although previous studies on T. texanum have included morphological examinations and nuclear gene-based molecular analyses, studies focusing on its complete mitochondrial genome remain limited (Montoya et al. 2016). In this study, we characterize mitochondrial genome of T. texanum strain CSC21B0438 and reconstruct phylogeny of Trichoderma based on mitogenome. To date, most mitogenomes of Trichoderma have been studied in isolates from soil or plants (Kwak 2020; Özkalekaya et al. 2022; Wang et al. 2024), whereas studies on species isolated from other organisms are scarce. This is the first study to analyze the mitogenome of an endolichenic Trichoderma strain, providing a useful resource for understanding the genomic characteristics of Trichoderma from diverse environment.
Materials and methods
The studied strain of T. texanum was isolated from lichen Parmotrema A. Massal. 1860 in Geoje-si, South Korea (34.998783 N, 128.704916 E) and cultured on potato dextrose agar (PDA, Difco Laboratories, Detroit, MI) at 25 °C (Figure 1). The strain CSC21B0438 was identified using the morphological and molecular from the previous study (Oh and Jang 2025) and deposited in the herbarium of Changwon National University (Changwon, South Korea). Genomic DNA was extracted by AccuPrep^®^ Genomic extraction kit (Bioneer, Daejeon, South Korea). The quantity and quality of extracted DNA were measured by Qubit 4.0 (Thermo Fisher Scientific Inc., Waltham, MA) and 2100 Bioanalyzer (Agilent, Santa Clara, CA). The complete mitogenome sequencing was performed using Illumina NovaSeq platform from Macrogen (Seoul, South Korea). The strain CSC21B0438 and its DNA are deposited in the herbarium of Changwon National University, South Korea (Seung-Yoon Oh; [email protected]).
Colony morphology of Trichoderma texanum strain CSC21B0438 grown on potato dextrose agar for seven days at 25 °C. Scale bar = 1 cm. Photograph by Yunhyeok Jang.
Raw reads were quality-filtered and trimmed by FASTP (Chen et al. 2018). The filtered reads were assembled de novo using NOVOPlasty v3.6 (Dierckxsens et al. 2017). Using the approach described by Ni et al. (2023), we generated a coverage depth map, which revealed an average sequencing depth of 5440× (Figure S1). MFannot (Lang et al. 2023) and GeSeq (Tillich et al. 2017) were used to annotate mitogenomes. tRNAscan-SE was used to identify transfer RNA (tRNA) genes (Lowe and Chan 2016) and manual curation was performed using Geneious Prime 2021.1 (Dotmatics, Boston, MA). Circular gene map visualized by OGDRAW version 1.3.1 (Greiner et al. 2019). The complete mitogenome sequence of T. texanum obtained in this study has been deposited in NCBI (GenBank) under accession number PX395767.
The conserved 14 protein-coding genes (PCGs) were used for phylogenetic analysis (atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, and nad6). Amino acid sequences of PCGs were aligned using Clustal Omega (Sievers et al. 2011) and concatenated by using Geneious Prime. The best substitution model for each mitochondrial PCG was determined using ModelTest-NG (Darriba et al. 2020). Hypomyces aurantius (Pers.) Fuckel 1870 (KU666552) and Cladobotryum mycophilum (Oudem.) W. Gams & Hooz. 1970 (MT108299) were used as outgroup species (Deng et al. 2016; Chen et al. 2020). Phylogenetic analysis was performed using RAxML (Stamatakis 2014) using 1000 bootstrap replicates through the graphical interface raxmlGUI v2.0 (Edler et al. 2021).
Results
The complete mitogenome of T. texanum strain CSC21B0438 was assembled in a single contig as a closed circular DNA. It was 31,361 bp in length, and overall base composition was A 31.6%, C 12.8%, G 15.5%, and T 35.5%. It comprised 15 PCGs, one open reading frame (orf), 27 tRNA, and two ribosomal RNA genes (Figure 2). Among the PCGs, there are 14 conserved PCGs and one ribosomal protein S3 (rps3). There was one PCG (cox2) with one intron and large subunit ribosomal RNA (rnl) with two introns (Figure S2). The rps3 gene was localized within an intron of the rnl gene (Figure 2). Our analysis revealed that the cox1 intron is absent from the mitogenome of T. texanum. All of the PCGs were found to have ATG as the predicted initiation codon and TAA as the termination codon, while orf212 was found to start with TTA.
Annotated circular map of the mitochondrial genome of Trichoderma texanum strain CSC21B0438. Gene clusters were shown with different color blocks. The complete mitochondrial genome comprised 15 protein coding genes (PCGs), one open reading frame (orf), 27 transfer RNA, and two ribosomal RNA genes. Arrows indicate the transcriptional direction of genes. The inner gray ring depicts the GC content distribution along the mitochondrial genome, illustrating local variation in nucleotide composition.
Phylogenetic analysis was performed on the conserved PCGs of mitogenomes of 22 Trichoderma species (Figure 3). For ML analysis, the model of substitution selected was JTT for cob, cox1, cox2, cox3, atp6, atp9, nad1, nad4, and nad3; WAG for atp8 and nad2; CPREV for nad6 and nad5; and BLOSUM62 for nad4L. The phylogenetic tree showed the Trichoderma genus formed a monophyletic clade. Among the genus, T. cornu-damae (Pat.) Z.X. Zhu & W.Y. Zhuang 2014 was early separated, and T. texanum is in close relation to T. koningiopsis Samuels, Carm. Suárez & H.C. Evans 2006 with a well-supported clade (bootstrap value = 99%).
Phylogenetic relationship of Trichoderma species based on 14 conserved protein-coding genes (atp6, atp8, atp9, cob, cox1, cox2, cox3, nad1, nad2, nad3, nad4, nad4L, nad5, and nad6). Numbers below branches indicate maximum-likelihood bootstrap support values. Only bootstrap values >50% are shown. The following mitochondrial genomes were used: T. koningiopsis, T. subviride, T. asperelloides, T. ghanense, T. longibrachiatum, T. velutinum, T. reesei, T. gracile, T. virens, T. zeloharzianum, T. brevicompactum, T. pyramidale, T. breve (Wang et al. 2024), T. asperellum, T. hamatum, T. atroviride, T. gamsii (Kwak 2020), T. harzianum (Kwak 2021), T. afroharzianum (Özkalekaya et al. 2022), T. lixii (Castrillo et al. 2023), T. cornu-damae (Lee et al. 2022), Cladobotryum mycophilum (Chen et al. 2020), and Hypomyces aurantius (Deng et al. 2016). GenBank accession numbers are provided following the species names.
Discussion and conclusions
We report, for the first time, the sequencing and characterization of the complete mitogenome of T. texanum. Many of the mitochondrial genomic features identified in T. texanum were consistent with those reported for other Trichoderma species. The mitogenomes of Trichoderma species range from 26,276 to 94,608 bp in length (Lee et al. 2022; Wang et al. 2024), and T. texanum falls within this range (31,361 bp). In most Trichoderma species, the stop codon of nad4L (TAA) and the start codon of nad5 (ATG) overlap by a single nucleotide, with the shared adenine (A) functioning as both the terminal base of nad4L and the initial base of nad5 (Kwak 2021; Özkalekaya et al. 2022). This single base overlap was also observed in T. texanum genome. In fungal mitogenomes, the rps3 gene is often located within the intron of the rnl gene (Özkalekaya et al. 2022; Oh 2024). Consistent with previous reports, this study also detected the rps3 gene within the intron of the rnl gene. In contrast to these shared features, one notable difference was observed in the cox1 gene. The cox1 gene contains intron in some Trichoderma species (Wang et al. 2024); however, in our studies, the intron of cox1 gene is not detected in T. texanum. In the context of the phylogenetic distribution of this intron, its absence in T. texanum suggests a possible lineage-specific intron loss. The results of the phylogenetic analysis confirmed that the genus Trichoderma forms a well-supported monophyletic clade and that T. texanum is closely related to T. koningiopsis, in agreement with previous reports (Montoya et al. 2016; Kullnig-Gradinger et al. 2002). In conclusion, this study expands our knowledge of the complete mitogenome of T. texanum and provides a foundation for future research to fully explore its potential applications.
Supplementary Material
Supplemental Material
Supplemental Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castrillo ML et al. 2023. Trichoderma koningiopsis (Hypocreaceae) has the smallest mitogenome of the genus Trichoderma. Front Microbiol. 14:1141087. 10.3389/fmicb.2023.114108737383640 PMC 10294050 · doi ↗ · pubmed ↗
- 2Chen C et al. 2020. The complete mitochondrial genome of Cladobotryum mycophilum (Hypocreales: Sordariomycetes). Mitochondrial DNA B Resour. 5(3):2595–2596. 10.1080/23802359.2020.174260033457872 PMC 7782004 · doi ↗ · pubmed ↗
- 3Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. 10.1093/bioinformatics/bty 56030423086 PMC 6129281 · doi ↗ · pubmed ↗
- 4Darriba D et al. 2020. Model Test-NG: a new and scalable tool for the selection of DNA and protein evolutionary models. Mol Biol Evol. 37(1):291–294. 10.1093/molbev/msz 18931432070 PMC 6984357 · doi ↗ · pubmed ↗
- 5Deng Y et al. 2016. Analysis of the mitochondrial genome in Hypomyces aurantius reveals a novel twintron complex in fungi. Int J Mol Sci. 17(7):1049. 10.3390/ijms 1707104927376282 PMC 4964425 · doi ↗ · pubmed ↗
- 6Dierckxsens N, Mardulyn P, Smits G. 2017. NOVO Plasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e 18. 10.1093/nar/gkw 95528204566 PMC 5389512 · doi ↗ · pubmed ↗
- 7Edler D, Klein J, Antonelli A, Silvestro D. 2021. R Ax MLGUI 2.0: a graphical interface and toolkit for phylogenetic analyses using R Ax ML. Methods Ecol Evol. 12(2):373–377. 10.1111/2041-210X.13512 · doi ↗
- 8Greiner S, Lehwark P, Bock R. 2019. Organellar Genome DRAW (OGDRAW) version 1.3.1: expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 47(W 1):W 59–W 64. 10.1093/nar/gkz 23830949694 PMC 6602502 · doi ↗ · pubmed ↗