Complete mitochondrial genome and phylogenetic analysis of the sea anemone Heteractis doreensis (Quoy & Gaimard, 1833)
Yanling Liao, Yirong Ye, Wenping Wang, Jinxing Fu, Xueying Zhang, Bingmiao Gao, Tianle Tang

TL;DR
This paper reports the first complete mitochondrial genome of Heteractis doreensis and its evolutionary relationships with other sea anemones.
Contribution
The study provides the first complete mitogenome of Heteractis doreensis and its phylogenetic placement within Hexacorallia.
Findings
The mitochondrial genome of H. doreensis is 20,720 bp long with 60.34% A+T content.
Phylogenetic analysis places H. doreensis in a strongly supported clade with Stichodactyla haddoni and Heteractis crispa.
The findings offer molecular insights into the evolution and phylogeography of H. doreensis.
Abstract
The first complete mitochondrial genome of Heteractis doreensis (Quoy & Gaimard, 1833) was sequenced to clarify its mitogenomic features and phylogenetic position. The circular genome is 20,720 bp long with 60.34% A+T content, comprising 13 protein-coding genes, two ribosomal RNA genes, two transfer RNA genes, and one putative open reading frame. Maximum-likelihood phylogenetic analysis of 19 Hexacorallia species revealed that H. doreensis clusters with Stichodactyla haddoni and Heteractis crispa in a strongly supported clade (bootstrap = 96%) within Stichodactylidae. These findings provide valuable molecular insights into the evolution and phylogeography of H. doreensis, supporting future research on its conservation and breeding.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1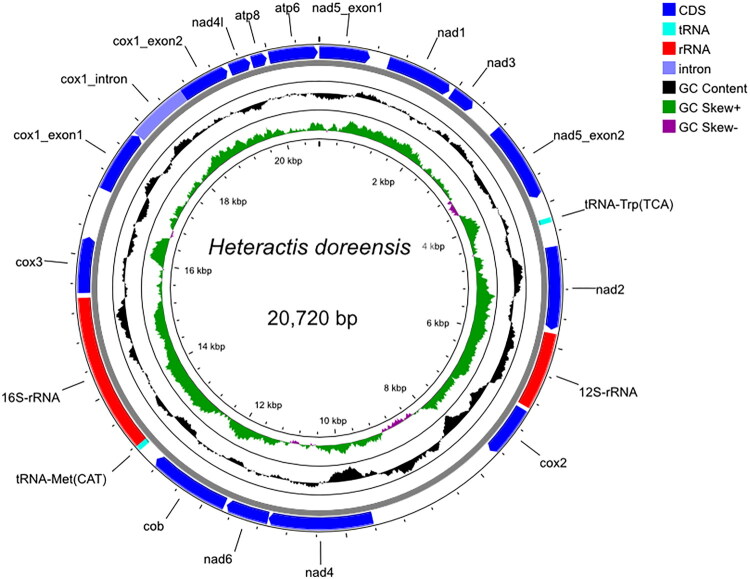 Figure 2
Figure 2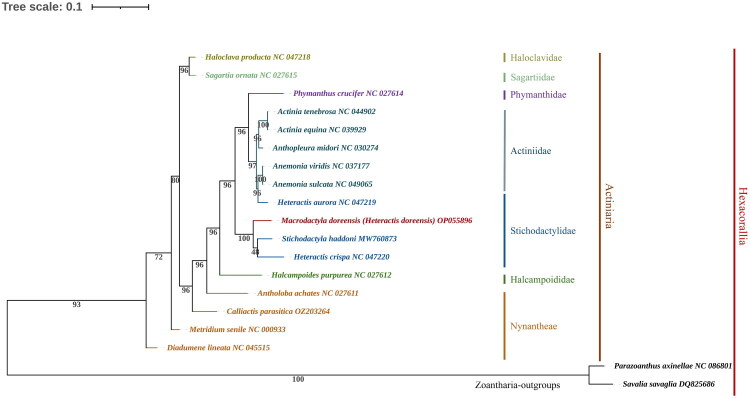 Figure 3
Figure 3- —Hainan Provincial Natural Science Foundation of China
- —Hainan Provincial Natural Science Foundation of China
- —Hainan Provincial Higher Education Institutions Scientific Research Project
- —National College Students Innovation and Entrepreneurship Training Program10.13039/501100013254
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Invertebrate Physiology and Ecology · Marine and coastal plant biology · Genomics and Phylogenetic Studies
Introduction
Heteractis doreensis (Figure 1) is a sea anemone belonging to the order Actiniaria, suborder Enthemonae, family Stichodactylidae, and genus Heteractis (Daly et al. 2016). Sea anemones are widely distributed from tropical to deep-sea environments, including depths exceeding 10,000 m, and produce a diverse array of peptide and protein toxins critical for prey capture, defense, and interspecific competition (Madio et al. 2017, 2018; Menezes and Thakur 2022). These bioactive compounds have emerging applications in psychopharmacology and medical drug development (O’Hara et al. 2021). High-throughput omics approaches have identified numerous sea anemone peptide neurotoxins (Kim et al. 2017). The mitochondrial genome is characterized by maternal inheritance, compact size, and relatively rapid evolution. Its replication is regulated by nuclear-encoded factors. Since mitochondrial DNA is abundant and amenable to PCR-based amplification, complete mitochondrial genomes are widely used in molecular systematics. Despite their ecological and pharmacological significance, genomic resources for H. doreensis are limited. Therefore, sequencing and comparative analysis of the mitochondrial genome of H. doreensis offers an opportunity to clarify the species’ evolutionary characteristics and phylogenetic position. Research on Actiniaria phylogeny has addressed taxonomy, nomenclature, identification, and biodiversity issues (Sinniger et al. 2007; Oliveira et al. 2012; Stampar et al. 2022). With advances in sequencing technologies, whole mitochondrial genomes have become an increasingly powerful tool for resolving phylogenetic relationships within Cnidaria (Kayal et al. 2015; Novosolov et al. 2022; González Muñoz et al. 2023; Li et al. 2023). This study determined the complete mitochondrial genome of H. doreensis and resolved its phylogenetic placement. These data enhance our understanding of the genetic diversity and evolutionary relationships within Stichodactylidae and provide a molecular reference for taxonomic resolution, comparative genomics, and conservation planning.
Reference image of H. doreensis taken by Bingmiao Gao.
Materials and methods
H. doreensis specimens were collected from the Xisha Islands, South China Sea (16.34° N, 111.49° E) on 17 May 2021. A specimen was deposited at the Key Laboratory of Tropical Translational Medicine, Ministry of Education, Hainan Medical University (Bingmiao Gao, [email protected]), Haikou, Hainan, China under voucher number CHMU0267. Species identity was determined based on morphological characteristics, including size, coloration, and basal column morphology, and was confirmed by BLAST analysis through comparison of the sequencing results with reference sequences in the NCBI database.
The complete mitochondrial genome was extracted using the Column mtDNAout kit (Tianda, Beijing, China). The sample concentration and purity were assessed using a NanoDrop™ Spectrophotometer (Thermo Fisher Scientific, Waltham, MA) and Qubit^®^ 2.0 Fluorometer (Life Technologies, Waltham, MA), respectively. Qualified DNA samples were then subjected to Illumina standard library preparation (San Diego, CA), and a paired-end sequencing library with an insert size of approximately 350 bp was constructed. Following library preparation, quality control was performed using quantitative PCR and an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA). Finally, DNA libraries that met the quality criteria were sequenced on an Illumina NovaSeq 6000 high-throughput platform (Illumina, San Diego, CA) using a PE150 sequencing strategy. Evaluation of the sequencing data quality and coverage revealed an average depth of 58.74× across the mitochondrial genome (Supplementary Figure 1).
The obtained clean reads were assembled de novo using SPAdes (version 3.5.0) (Bankevich et al. 2012). Genome annotation was performed using MITOS (Bernt et al. 2013) and open reading frame Finder (Cheng et al. 2013), while transfer RNA (tRNA) genes were specifically annotated using ARWEN (Laslett and Canbäck 2008) and tRNAscan-SE (Lowe and Chan 2016). Phylogenetic analyses were performed based on the complete mitochondrial genome of H. doreensis and 18 species across the subclass Hexacorallia (Beagley et al. 1998; Foox et al. 2016; Zhang and Zhu 2017; Chi et al. 2018; Surm et al. 2019; Fu et al. 2021; Johansen et al. 2021), including Actiniidae, Halcampoididae, Haloclavidae, Nyantheae, Phymanthidae, Sagartiidae, Stichodactylidae, and Zoantharia. Evolutionary analyses were conducted in MEGA11 (Tamura and Nei 1993), and the evolutionary history was inferred using the maximum-likelihood (ML) method with the Tamura–Nei substitution model (Tamura et al. 2021). Initial heuristic trees were generated automatically by applying the Neighbor-Joining and BioNJ algorithms to a pairwise distance matrix estimated under the Tamura–Nei model, followed by the selection of a topology with the highest log-likelihood value. Branch support was evaluated using 1000 bootstrap replicates (Barroso et al. 2025).
Results
The mitochondrial genome of H. doreensis is a circular molecule of 20,720 bp in length (GenBank accession number: OP055896) (Supplementary Figure 2). Its overall nucleotide composition is 26.64% A, 17.84% C, 21.82% G, and 33.71% T, corresponding to GC and AT contents of 39.66% and 60.34%, respectively. The genome contains 13 protein-coding genes (PCGs), two tRNA genes (tRNA^Trp^ and tRNA^Met^), two ribosomal RNA (rRNA) genes (12S rRNA and 16S rRNA), and several non-coding regions (Supplementary Table 1). These genomic features collectively reflect the typical mitogenome architecture observed in Hexacorallia, with 13 PCGs spanning 11,847 bp, accounting for 57.18% of the total genome length. The 12S rRNA gene (1081 bp) is located between nad1 and cox2, whereas the 16S rRNA gene (1714 bp) is positioned between tRNA^Met^ and cox3. All PCGs, except nad1, initiate with the ATG start codon. Termination codons vary, with nad2, nad5, and nad4l ending with TAG, whereas the remaining PCGs terminate with TAA (Figure 2). Furthermore, assembly metrics and inspection of terminal overlaps confirmed that the sequence was complete and circular, without any gaps or unresolved regions. The continuity between the terminal region (cox1-atp6) and the start region (nad5) further supports the circular organization of the mitogenome.
Mitochondrial genome map of H. doreensis. The complete mitochondrial genome of H. doreensis was 20,720 bp long and included 13 PCGs, two tRNA genes, and two rRNA genes. The overall base composition was as follows: 26.64% for A, 17.84% for C, 21.82% for G, and 33.71% for T. The outermost circle represents the gene name, direction of the encoding gene, and transcription; tRNA and rRNA have no direction. Inside the black circle is the GC proportion, and further inside is the GC offset. The innermost circle represents the length coordinate.
To investigate the evolutionary implications of the newly sequenced mitogenome, we performed a phylogenetic analysis. Phylogenetic reconstructions based on homologous mitochondrial markers indicate that the genus Macrodactyla comprises multiple divergent evolutionary lineages. Consequently, Macrodactyla doreensis was recently reclassified within the genus Heteractis based on strong nodal support (bootstrap > 90%) and shared synapomorphic characters (Hua et al. 2024). Accordingly, the specimen previously labeled as M. doreensis in Figure 3 is referred to as H. doreensis. We further analyzed the mitochondrial genome of H. doreensis, together with those of 18 additional Hexacorallia species, to infer relationships within the subclass. The resulting ML phylogeny revealed that H. doreensis clustered with other members of the genus Heteractis, which was supported by robust node support.
ML phylogenetic tree for H. doreensis and other Zoantharia species based on the complete mitochondrial genome. Bootstrap support values are indicated at each node. Parazoanthus axinellae and Savalia savaglia, both belonging to the order Zoantharia, were used as the outgroups. The dataset includes the following mitochondrial genomes: Macrodactyla doreensis (OP055896; this study name is Heteractis doreensis), Actinia tenebrosa (NC044902; Surm et al. 2019), Actinia equina (NC039929; Wilding and Weedall 2019), Anthopleura midori (NC030274; Zhang and Zhu 2017), Anemonia sulcata (NC049065; unpublished), Anemonia viridis (NC037177; Chi et al. 2018), Halcampoides purpurea (NC027612; Foox et al. 2016), Haloclava producta (NC047218; unpublished), Metridium senile (NC000933; Beagley et al. 1998), Diadumene lineata (NC045515; unpublished), Calliactis parasitica (OZ203264; unpublished), Antholoba achates (NC027611; Foox et al. 2016), Phymanthus crucifer (NC027614; Foox et al. 2016), Sagartia ornata (NC027615; Foox et al. 2016), Stichodactyla haddoni (MW760873; Johansen et al. 2021), Heteractis crispa (NC047220; unpublished), Heteractis aurora (NC047219; unpublished), Savalia savaglia (DQ825686; Sinniger et al. 2007), and Parazoanthus axinellae (NC086801; unpublished).
Discussion and conclusions
This study presents the first complete mitochondrial genome of H. doreensis, revealing the conservation of mitogenome structure and molecular characteristics. Previous research (Yap et al. 2023) demonstrated that the genus Macrodactyla is not monophyletic, as evidenced by mitochondrial (12S, 16S, cox3) and nuclear (28S) markers. Based on the complete mitochondrial genome of H. doreensis, our phylogenetic analysis aligns with these findings and further supports the reassignment of M. doreensis to the genus Heteractis. The ML tree reconstructed using MEGA11 places H. doreensis in a strongly supported clade (bootstrap = 96%) with Stichodactyla haddoni and Heteractis crispa, reinforcing its position within the family Stichodactylidae. This finding confirms the evolutionary distinction of H. doreensis and resolves long-standing taxonomic uncertainties surrounding this species (Figure 3). Currently, mitogenomic data for Actiniaria are limited, with substantial gaps in taxonomic representation across different families. This uneven sampling may challenge the accuracy of the phylogenetic framework by affecting branch support and topological stability at the deeper nodes of the tree. Therefore, expanding the mitogenomic dataset through further sequencing of underrepresented groups is essential for testing the robustness of the current phylogeny and uncovering broader evolutionary patterns in sea anemones. Our findings clarify the evolutionary position of H. doreensis within Stichodactylidae and contribute to the expansion of the mitochondrial genomic resources available for sea anemones. These resources not only enhance our understanding of the evolutionary history of this diverse group but also provide a more comprehensive foundation for future taxonomic and evolutionary studies within Actiniaria, facilitating comparative genomic research and advancing our understanding of the processes that shape their diversities.
Supplementary Material
Supplementary Figure 1.jpg
Supplementary table 1.xls
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bankevich A et al. 2012. SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 19(5):455–477. 10.1089/cmb.2012.002122506599 PMC 3342519 · doi ↗ · pubmed ↗
- 2Barroso RA et al. 2025. Proteomic diversity of the sea anemone Actinia fragacea: comparative analysis of nematocyst venom, mucus, and tissue-specific profiles. Mar Drugs. 23(2):79. 10.3390/md 2302007939997203 PMC 11857728 · doi ↗ · pubmed ↗
- 3Beagley CT, Okimoto R, Wolstenholme DR. 1998. The mitochondrial genome of the sea anemone Metridium senile (Cnidaria): introns, a paucity of t RNA genes, and a near-standard genetic code. Genetics. 148(3):1091–1108. 10.1093/genetics/148.3.10919539427 PMC 1460033 · doi ↗ · pubmed ↗
- 4Bernt M et al. 2013. MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69(2):313–319. 10.1016/j.ympev.2012.08.02322982435 · doi ↗ · pubmed ↗
- 5Cheng JK, Zeng X, Ren GM, Liu ZH. 2013. CGAP: a new comprehensive platform for the comparative analysis of chloroplast genomes. BMC Bioinformatics. 14(1):95. 10.1186/1471-2105-14-9523496817 PMC 3636126 · doi ↗ · pubmed ↗
- 6Chi SI, Urbarova I, Johansen SD. 2018. Expression of homing endonuclease gene and insertion-like element in sea anemone mitochondrial genomes: lesson learned from Anemonia viridis. Gene. 652:78–86. 10.1016/j.gene.2018.01.06729366757 · doi ↗ · pubmed ↗
- 7Daly M et al. 2016. The phylum Cnidaria: a review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa. 1668(1):127–182. 10.5281/zenodo.180149 · doi ↗
- 8Foox J, Brugler M, Siddall ME, Rodríguez E. 2016. Multiplexed pyrosequencing of nine sea anemone (Cnidaria: Anthozoa: Hexacorallia: Actiniaria) mitochondrial genomes. Mitochondrial DNA A DNA Mapp Seq Anal. 27(4):2826–2832. 10.3109/19401736.2015.105311426104159 · doi ↗ · pubmed ↗