Taxonomic revision of the freshwater mussel subtribe Cristariina (Bivalvia, Unionidae) in China: description of a new species and new synonyms

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
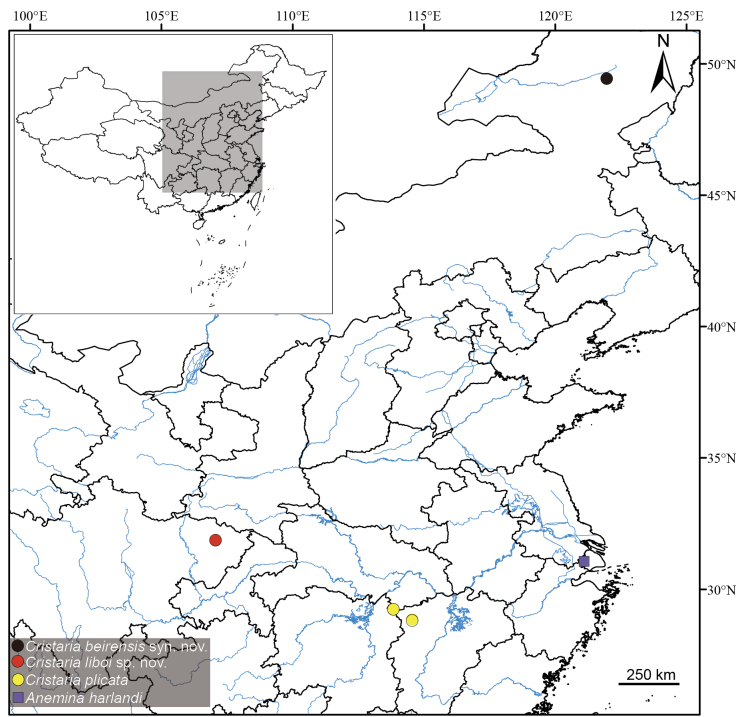 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3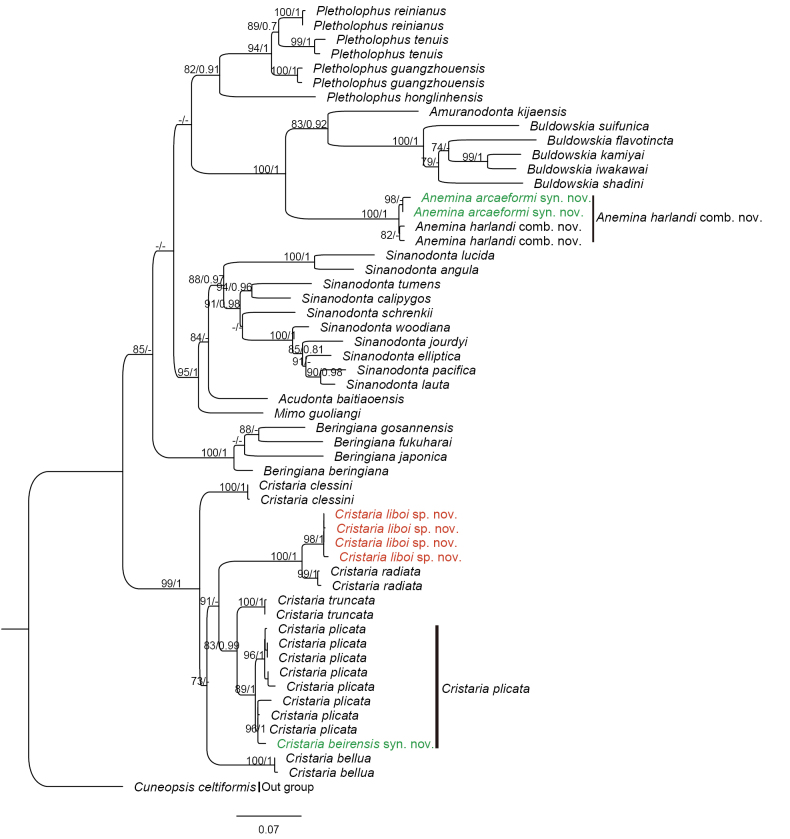 Figure 4
Figure 4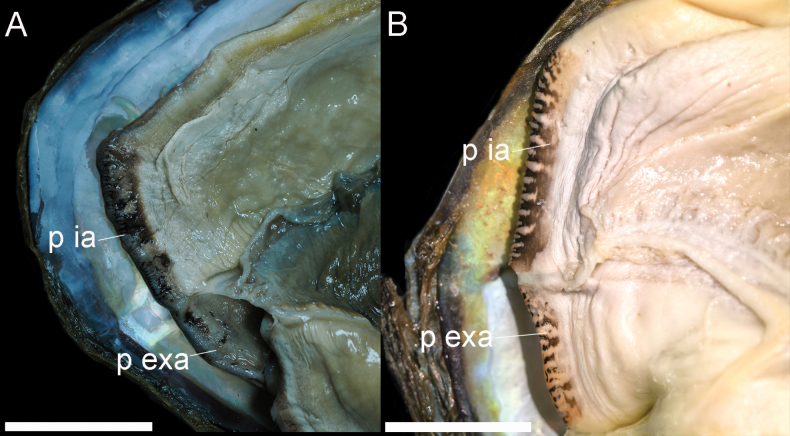 Figure 5
Figure 5| Species | NCBI | Locality |
|---|---|---|
|
|
| Laos: Mekong basin, Nam Ngum River |
|
| Laos: Mekong basin, Nam Ngum River | |
|
|
| Japan |
|
| Japan | |
|
| China: Sichuan | |
|
| China: Sichuan | |
| China: Sichuan, Mianyang, Furong Creek | ||
| China: Sichuan, Mianyang, Furong Creek | ||
|
|
| |
|
| ||
|
|
| Vietnam: Bang River |
|
| Vietnam: Bang River | |
|
|
| |
|
| ||
|
| ||
|
| ||
| China: Inner Mongolia, Hulunbuir City | ||
| China: Inner Mongolia, Hulunbuir City | ||
| China: Hunan, Yiyang | ||
| China: Hunan, Yiyang | ||
| China: Hunan, Changde | ||
|
|
| Japan: Fukuoka, Munakata |
|
| Japan: Fukuoka, Munakata | |
|
|
| Vietnam |
|
| Vietnam | |
|
|
| China: Guangdong, Ghuangzhou, Liuxi River |
|
| China: Guangdong, Ghuangzhou, Liuxi River | |
|
|
| Japan |
|
|
| Russia |
|
|
| Russia: Primorsky Krai, Soldatskoe Lake |
|
|
| South Korea |
|
|
| Japan |
|
|
| Japan |
|
|
| Russia: Primorsky Krai, Melgunovka River |
|
|
| China: Hunan, Dongting Lake |
|
| China: Hunan, Dongting Lake | |
| China: Shanghai City, Chenshan Botanical Garden | ||
| China: Shanghai City, Chenshan Botanical Garden | ||
|
|
| Japan |
|
|
| Japan |
|
|
| USA: Alaska, Eugumen Lake |
|
|
| Japan: Gifu, Ena City |
|
|
| China: Sichuan |
|
|
| China |
|
|
| China: Hunan, Anren County |
|
|
| Japan: Osaka, Yodo River |
|
|
| Japan |
|
|
| Russia: Primorsky Krai, Melgunovka River |
|
|
| China: Jiangxi, Gan River |
|
|
| Vietnam |
|
|
| China: Jiangxi, Gan River |
|
|
| Malaysia: Padas basin |
|
|
| South Korea |
|
|
| China: Jiangxi, Gan River |
|
|
| China: Yunnan, Luliang |
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|
|
| 0.2 | |||||
|
| 12.3 | 0.1 | ||||
|
| 11.8 | 3.3 | 0.1 | |||
|
| 7.9 | 11.3 | 11.1 | 0 | ||
|
| 8.2 | 10.6 | 9.8 | 4.9 | 1.1 | |
|
| 8.7 | 12.7 | 12.1 | 8.7 | 8.8 | 0.3 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquatic Invertebrate Ecology and Behavior · Marine Biology and Ecology Research · Marine Bivalve and Aquaculture Studies
Introduction
Freshwater mussels of the family Unionidae perform crucial ecological functions (Vaughn and Hakenkamp 2001; Aldridge et al. 2007), yet they rank among the globally most imperilled groups of freshwater organisms (Lopes-Lima et al. 2020). The Chinese unionid fauna is remarkably diverse, with numerous species recorded in the Yangtze River Basin (Heude 1885; Simpson 1900; He and Zhuang 2013; Graf and Cummings 2021; Guo 2022; Chen et al. 2023; Dai et al. 2024a, 2024b; Chen et al. 2025). Although recent studies (Wu et al. 2025; Xiang et al. 2025) have offered a preliminary framework for the Chinese species of the subtribe Cristariini (Klishko et al. 2016; Wu et al. 2025), a comprehensive revision is still needed to resolve persistent issues. For instance, the relationship between Anemina harlandi (Baird & H. Adams, 1867), comb. nov. and Anemina arcaeformis (Heude, 1877), syn. nov. requires clarification. Furthermore, the taxonomic status of Cristaria beirensis Y.-Y. Liu & W.-Z. Zhang, 1982, syn. nov. is questionable (Wu et al. 2025), as records from Sichuan Province are geographically disjunct from its type locality in Hulunbuir City, Inner Mongolia. To address these issues, this study re-collected the aforementioned species and re-assessed their relationships through integrated morphological and phylogenetic analyses. Additionally, we describe a new species Cristaria liboi sp. nov.
Materials and methods
Specimen sampling and morphological observations
Cristaria beirensis Y.-Y. Liu & W.-Z. Zhang, 1982 were collected in Hexin Park (49.2083°N, 119.7722°E), Hailar District, Hulunbuir City, Inner Mongolia, China, in August 2025; Cristaria plicata (Leach, 1814) were collected in Yiyang City (28.5969°N, 112.3581°E) and Changde City (28.9823°N, 111.6459°E), Hunan Province, China, in August 2025; Cristaria liboi sp. nov. were collected in Furong Creek (31.6296°N, 104.8649°E), Mianyang City, Sichuan Province, China, in August 2025; Anemina harlandi (Baird & H. Adams, 1867), comb. nov. were collected in Chenshan Botanical Garden (31.08°N, 121.1849°E), Shanghai City, China, in August 2025 (Figs 1, 2, 3). The specimens were preserved in 95% ethanol, and deposited at the Museum of Biology, Nanchang University (NCUMB), China. Vernier callipers were used to measure shell length (L), width (W), and height (H).
Sampling localities of specimens utilized in this study. The locality of Cristaria beirensis, which is treated herein as a junior synonym of C. plicata (syn. nov.), is distinctly marked. Symbols: black circle, C. beirensis (syn. nov. of C. plicata); red circle, C. liboi sp. nov.; yellow circle, C. plicata; purple square, Anemina harlandi.
A–H. Cristaria plicata (Leach, 1814); A–D. C. beirensis syn. nov., NCUMB_CB251001, from Hexin Park, Hailar District, Hulunbuir City, Inner Mongolia, China; E, F. NCUMB_CB251002, from Yiyang City, Hunan Province, China; G, H. NCUMB_CB251003 from Changde City, Hunan Province; I–P. C. liboi Chen, Xiang, He & Guo, sp. nov. from Furong Creek, Mianyang City, Sichuan Province, China; I–L. Holotype, NCUMB_CL250901; M–P. Paratype: NCUMB_CL250902. Scale bar: 50 mm.
Anemina harlandi (Baird & H. Adams, 1867), NCUMB_AH251004, from Shanghai, China. Scale bar: 20 mm.
DNA extraction, PCR, sequencing, and phylogenetic analyses
Total genomic DNA was extracted on foot tissues using TIANamp Marine Animals DNA Kit (Tiangen Biotech) following the manufacturer’s protocol. Freshwater mussels exhibit doubly uniparental inheritance (DUI), a system in which highly differentiated maternal (F-type) and paternal (M-type) mitogenomes coexist. In males, the M-type genome is primarily localized in the germ line, whereas somatic tissues such as the foot are dominated by the F-type (Breton et al. 2007). As the majority of existing unionid COI barcode references are derived from somatic tissues (primarily the F-type), we targeted the F-type mitochondrial COI gene to ensure direct comparability. Amplification was performed using the primers LCO22me2 (GGTCAACAAAYCATAARGATATTGG) and HCO700dy2 (TCAGGGTGACCAAAAAAYCA) (Walker et al. 2007). Polymerase chain reaction (PCR) amplifications of COI were performed in a final 25 μL volume mixture containing 1 μL of template DNA, 1 μL of each pair of primers, 12.5 μL of Green Taq Mix (Vazyme, China), and 9.5 μL ddH2O. Thermal cycling began with one cycle at 98 °C for 10 s, followed by 35 cycles of denaturation at 94 °C for 1 min, 50 °C for 1 min, and 72 °C for 1 min, with a final extension step at 72 °C for 7 min. The PCR products were purified and sequenced using an ABI 3730XL analyzer. The newly obtained sequences were deposited in GenBank (Table 1). Sequences were aligned using MAFFT v. 7 based on the Q-INS-i method (Katoh et al. 2019). Pairwise distances between species were calculated using MEGA X (Kumar et al. 2018). The best substitution model was selected using the corrected Bayesian Information Criterion (BIC) in MODELFINDER v. 2.2.0 (Kalyaanamoorthy et al. 2017). Bayesian inference (BI) and maximum-likelihood (ML) analysis were performed using MrBayes v. 3.2.6 (Ronquist et al. 2012) and IQ-TREE v. 2.2 (Minh et al. 2020), respectively, with reference to the selected model of sequence evolution. For Bayesian analysis, two runs were performed simultaneously with four Markov chains starting from a random tree. Bayesian posterior probabilities (BPPs) of nodes were determined using Metropolis-coupled Markov chains (one cold chain) for 2 million generations, with sampling every 1000 generations. The first 25% of sampled trees were discarded as burn-in when the standard deviation of split frequencies of the two runs was less than 0.01; the remaining trees were then used to create a 50% majority-rule consensus tree and to estimate BPPs. Node support for maximum-likelihood analysis was determined using 1000 rapid bootstrap (BS) replicates.
Results
The GTR+F+I+R4 model was selected as the best-fit model of nucleotide substitution by the AIC criterion. The phylogenetic analyses showed that the genus Cristaria formed a monophyletic clade with high support (BS = 99%, BPP = 1). The new species Cristaria liboi sp. nov. collected from Mianyang, Sichuan, China, together with Cristaria radiata, formed a strongly supported monophyletic clade (BS = 100%, BPP = 1) (Fig. 4). The uncorrected p-distances within the genus Cristaria based on COI sequences ranged from 3.3% to 12.7%. The smallest interspecific distance (3.3%) was observed between the new species C. liboi sp. nov. and its closest relative, C. radiata, while the largest distance (12.7%) was found between C. liboi and C. bellua (Table 2). These distances as well as the distinct morphological characteristics provide compelling evidence for its classification as a new species C. liboi sp. nov. in the genus Cristaria.
Maximum-likelihood (ML) phylogenetic tree based on cytochrome c oxidase subunit I (COI) gene sequences. Nodal support is indicated as ML bootstrap values (BS) and Bayesian posterior probabilities (PP). Only values ≥ 70% (BS) and ≥ 0.80 (PP) are shown. Species names are colored to highlight taxonomic findings: red for the new species (Cristaria liboi sp. nov.), and green for the junior synonyms (C. beirensis and Anemina arcaeformis).
Systematics
Family Unionidae Rafinesque, 1820
Subfamily Unioninae Rafinesque, 1820
Tribe Anodontini Rafinesque, 1820
Subtribe Cristariini Lopes-Lima, Bogan & Froufe, 2017
Cristaria
Taxon classificationAnimaliaUnionidaUnionidae
Genus
Schumacher, 1817
6FC71910-C201-5C99-822F-009482989FAC
Taxonomic note.
Based on morphological and molecular evidence presented in this study, Cristaria beirensis (Xiong & Liu, 1994) is herein treated as a junior synonym of Cristaria plicata (Leach, 1814), (syn. nov.). The following description and discussion of C. plicata encompass the characteristics of both nominal forms.
Type species.
Cristaria plicata (Leach, 1814).
Cristaria
liboi
Taxon classificationAnimaliaUnionidaUnionidae
Chen, Xiang, He & Guo sp. nov.
B12DFFA4-544C-5E7B-BB36-7F5AA4BC3F01
https://zoobank.org/5220F1DA-BDEA-45BD-8938-E526A7D8E5F7
Material examined.
Holotype: • NCUMB_CL250901, shell length 185.21 mm, shell width 65.94 mm, shell height 111.25 mm (Fig. 2I–L), Furong Creek (31.6296°N, 104.8649°E), Mianyang City, Sichuan Province, China, in August 2025.
Paratypes: • 1 specimen, NCUMB_CL250902, shell length 185.13 mm, shell width 66.74 mm, shell height 113.85 mm (Fig. 2M–P), locality and habitat same as holotype.
Diagnosis.
Shell winged, large, oval, thick, solid; wing not obvious, posterior ridges high and obtuse. Umbo often eroded. Muscle scar obvious. Incurrent aperture without papillae arranged. Nacre lavender or white.
Description.
Shell winged, large, oval, black, thick, solid; wing not obvious. Posterior ridges high and obtuse; ventral margin curved; posterior slope prominent; periostracum sculptured with thick and obvious concentric growth lines. Right and left valve with an almost completely degraded pseudocardinal teeth and a short lateral tooth. Hinge well developed. Umbo often eroded. Muscle scar obvious. Nacre lavender or white (Fig. 2I–P). Incurrent aperture without papillae arranged and excurrent aperture with obvious papillae arranged in one row; incurrent aperture papillae are more than twice the size of the excurrent aperture, and both have black pigmentation (Fig. 5A).
Internal anatomy of the mantle cavity of Cristaria species. A. C. liboi sp. nov.; B. C. plicata. Abbreviations: p ia = papillae of the incurrent aperture; p exa = papillae of the excurrent aperture. Scale bars: 30 mm.
Etymology.
The name of this species is in honour of Mr Li Bo, a conchologist who collected the specimens.
Vernacular name.
李氏冠蚌 (li shi guan bang).
Habitat and distribution.
This species is currently only known to be distributed in the Furong Creek, Mianyang City, Sichuan Province (Fig. 1).
Remarks.
The species can be distinguished from other species in the genus by its characteristics, such as the incurrent aperture without arranged papillae and the wings being not obvious (Fig. 5A, B).
Cristaria
plicata
Taxon classificationAnimaliaUnionidaUnionidae
(Leach, 1814)
446B3527-5FB5-55E1-8BC7-B349947FD58F
Anodon herculeus G.B. Sowerby II, 1867: pl. 3, fig. 7.Anodonta (Dipsas) herculea von Middendorff, 1848: 302–304.Anodonta cristata Blainville, 1825: 631.Anodonta dipsas Blainville, 1825: 538, pl. 66, fig. 3.Anodonta spatiosa Clessin, 1876: 173, pl. 57, fig. 2.Craspedodonta smaragdina (Anton, 1838)—Clessin 1876: 93.Cristaria beirensis Y.-Y. Liu & W.-Z. Zhang, 1982: 35. Syn. nov.Cristaria (Cristaria) plicata (Leach, 1814)—Haas 1969: 387.Cristaria herculea (Middendorff, 1847)—Vinarski & Kantor, 2016: 45.Cristaria tuberculata Schumacher, 1817: 22.Dianisotis chinensis Rafinesque, 1831: 7.Dipsas bialatus (I. Lea, 1829)—Morle 1877: 266.Dipsas occidentalis Heude, 1885: pl. 66, fig. 129.Dipsas plicata Leach, 1814: 120, pl. 53.Dipsas plicatus Leach, 1814: 120, pl. 53.Symphynota bialata I. Lea, 1829: 445, pl. 14, fig. 24.Symphynota magnifica I. Lea, 1834 sensu Mabille & le Mesle, 1866—Brandt 1974: 278.Unio (Anodonta) smaragdinus Anton, 1838: 16.Unio smaragdina Anton, 1838: 16.
Material examined.
• Cristaria beirensis syn. nov., NCUMB_CB251001, Hexin Park, Hailar District, Hulunbuir City, Inner Mongolia, China (49.2083°N, 119.7722°E) (Fig. 2A–D); • C. plicata, NCUMB_CB251002, collected in Yiyang City (28.5969°N, 112.3581°E) (Fig. 2E, F), NCUMB_CB251003, collected in Changde City (28.9823°N, 111.6459°E), Hunan Province, China (Fig. 2G, H).
Diagnosis.
Shell winged, large, oval, solid; wing obvious. Muscle scar obvious. Nacre lavender or white.
Habitat and distribution.
This species is widespread in China, Russia, and Japan where it lives in freshwater ponds, streams, reservoirs, and lakes, typically on soft substrates composed of sand and mud.
Remarks.
Our specimens of C. beirensis were collected from near its type locality. Both morphological comparison (Fig. 2A–H) and molecular evidence from the phylogenetic tree (Fig. 4) demonstrate a lack of significant differences. Therefore, we formally regard Cristaria beirensis as a junior synonym of Cristaria plicata.
Anemina
Taxon classificationAnimaliaUnionidaUnionidae
Genus
F. Haas, 1969
FE4DF734-067D-537A-AC04-59006C0CF0D7
Type species.
Anodon arcaeformis Heude, 1877 accepted as Anemina harlandi (Baird & H. Adams, 1867), comb. nov.
Anemina harlandi (Baird & H. Adams, 1867), comb. nov.
Anemina arcaeformis (Heude, 1877)—Wu et al. 2025: 39. Syn. nov.
Anemina euscaphys (Heude, 1879)—He and Zhuang 2013: 31.
Anemina fluminea (Heude, 1877)—He and Zhuang 2013: 32.
Anemina globosula (Heude, 1878)—He and Zhuang 2013: 33.
Anodon arcaeformis Heude, 1877: pl. 19, fig. 40.
Anodon euscaphys Heude, 1879: pl. 35, fig. 68.
Anodon fluminea Heude, 1877: pl. 20, fig. 42.
Anodon globosula Heude, 1878: pl. 25, fig. 54.
Anodonta harlandi Baird & H. Adams, 1867: 492, pl. 26, fig. 3a.
Anodon torrentis Heude, 1878: pl. 29, fig. 61.
Anodonta (Haasiella) arcaeformis (Heude, 1877)—Korobkov 1954: 80.
Anodonta (Haasiella) euscaphys (Heude, 1879)—Korobkov 1954: 80.
Anodonta arcaeformis (Heude, 1877)—Lindholm 1925: 137.
Material examined.
• Anemina harlandi comb. nov., NCUMB_AH251004, 05, were collected in Chenshan Botanical Garden (31.0800°N, 121.1849°E), Shanghai City, China (Fig. 3).
Diagnosis.
Shell medium-sized, expanded, oval, thin, sub-glossy, opaque. Umbo inflated. Without pseudocardinal teeth.
Habitat and distribution.
This species is widespread in China where it lives in freshwater ponds, streams, reservoirs, and lakes.
Remarks.
Specimens of Anemina harlandi comb. nov. were collected near the type locality (Figs 1, 3) and correspond well to the illustrations provided in the original description. Morphological comparisons reveal no significant differences in shell shape between Anemina harlandi comb. nov. and Anemina arcaeformis. This conclusion is further supported by the phylogenetic analysis (Fig. 4). Therefore, Anemina arcaeformis syn. nov. is herein proposed as a junior synonym of Anemina harlandi comb. nov. Currently, this species represents the only extant member of the genus Anemina F. Haas, 1969.
Discussion
This study involved sampling near the type locality of Cristaria beirensis (Fig. 2A–D), and through phylogenetic tree analysis (Fig. 4), it was confirmed that C. beirensis is a junior synonym of Cristaria plicata. Currently, there are four species of Cristaria recorded in China: C. truncata Dang, 198, C. plicata, C. liboi sp. nov., and C. radiata (Simpson, 1900). The species diversity of Cristaria in China is mainly concentrated in the Yangtze river basin, with C. plicata being most widespread. In northern China, Cristaria species are very limited, with only C. plicata present. Its shell morphology exhibits certain plasticity in different regions (Klishko et al. 2016). For C. plicata in different regions, there are slight variations in the incurrent and excurrent apertures. The taxonomic basis for recent records of C. radiata (e.g. Wu et al. 2025) is unreliable, as the source of the COI sequence (Jia et al. 2009) did not provide voucher imagery or a morphological identification to confirm the species assignment. Therefore, future investigations should involve collecting specimens from the type locality to determine the true identity of C. radiata.
A recent study proposed Anemina arcaeformis as the only species within the genus Anemina. However, the authors of that study overlooked the taxon historically known as Anodonta harlandi. Although A. harlandi had been treated as a junior synonym of Sinanodonta woodiana (I. Lea, 1834), its shell morphology is congruent with specimens identified as A. arcaeformis by Wu et al. (2025). To clarify this taxonomic confusion, we collected new topotypic specimens (from the type locality of A. harlandi, Figs 1, 3) matching the original description. Our molecular phylogenetic analyses (Fig. 4) conclusively demonstrate that A. arcaeformis is not a distinct species but is conspecific with A. harlandi. Therefore, we herein establish the revised classification: Anemina harlandi (Heude, 1877), comb. nov., with A. arcaeformis as its junior synonym (syn. nov.).
Based on the current research, species in the subtribe Cristariini that are endemic to China often have very narrow distributions, making them highly susceptible to human activities, leading to their potential endangerment. Some species may already be extinct (Xiang et al. 2025), so further systematic investigations of their distribution are necessary, along with the development of corresponding conservation measures.
Supplementary Material
XML Treatment for Cristaria
XML Treatment for Cristaria liboi
XML Treatment for Cristaria plicata
XML Treatment for Anemina
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aldridge DC, Fayle TM, Jackson N (2007) Freshwater mussel abundance predicts biodiversity in UK lowland rivers. Aquatic Conservation 17(6): 554–564. 10.1002/aqc.815 · doi ↗
- 2Anton HE (1838) Verzeichniss der Conchylien welche sich in der Sammlung von Herrmann Eduard Anton befinden. Herausgegeben von dem Besitzer. Anton, Halle, [xvi +] 110 pp. 10.5962/bhl.title.11509 · doi ↗
- 3Baird W, Adams H (1867) Notes upon some interesting Chinese shells, with a description of two or three new species of Unionidæ, collected at Shanghai by Jones Lamprey M.D., 67th Regiment, C.M.Z.S. Proceedings of the Zoological Society of London 1867: 489–492[, pl. 26].
- 4Brandt RAM (1974) The non-marine aquatic Mollusca of Thailand. Archiv für Molluskenkunde 105: [i–iv,] 1–423.
- 5Breton S, Beaupré HD, Stewart DT, Hoeh WR, Blier PU (2007) The unusual system of doubly uniparental inheritance of mt DNA: isn’t one enough? Trends in Genetics 23: 465–474. 10.1016/j.tig.2007.05.01117681397 · doi ↗ · pubmed ↗
- 6Chen ZG, Dai YT, Ouyang S, Huang XC, Wu XP (2023) Unveiling the identity of Diaurora Cockerell, 1903 (Bivalvia, Unionidae): Morphology, molecular phylogenetics, and the description of a new species. Zoo Keys 1173: 131–144. 10.3897/zookeys.1173.106148 PMC 1041589837577154 · doi ↗ · pubmed ↗
- 7Chen H, Xiang HQ, Dai YT, Chen ZG, He YM, Guo G, Wu XP, Huang XC (2025) A new genus and two new species of freshwater mussels (Bivalvia, Unionidae) from Sichuan, China: Overlooked cryptic endemism in the upper Yangtze River Basin. Zoosystematics and Evolution 101(4): 1459–1470. 10.3897/zse.101.161018 · doi ↗
- 8Clessin S (1876) Die Gattung Anodonta nebst den übrigen Najaden mit unvolkommenem Schloss. Systematisches Conchylien-Cabinet 9(1): 65–288.