Angiopteris weimingii (Marattiaceae): a new endangered fern species from southern Yunnan, China, revealed by morphology and phylogeny

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
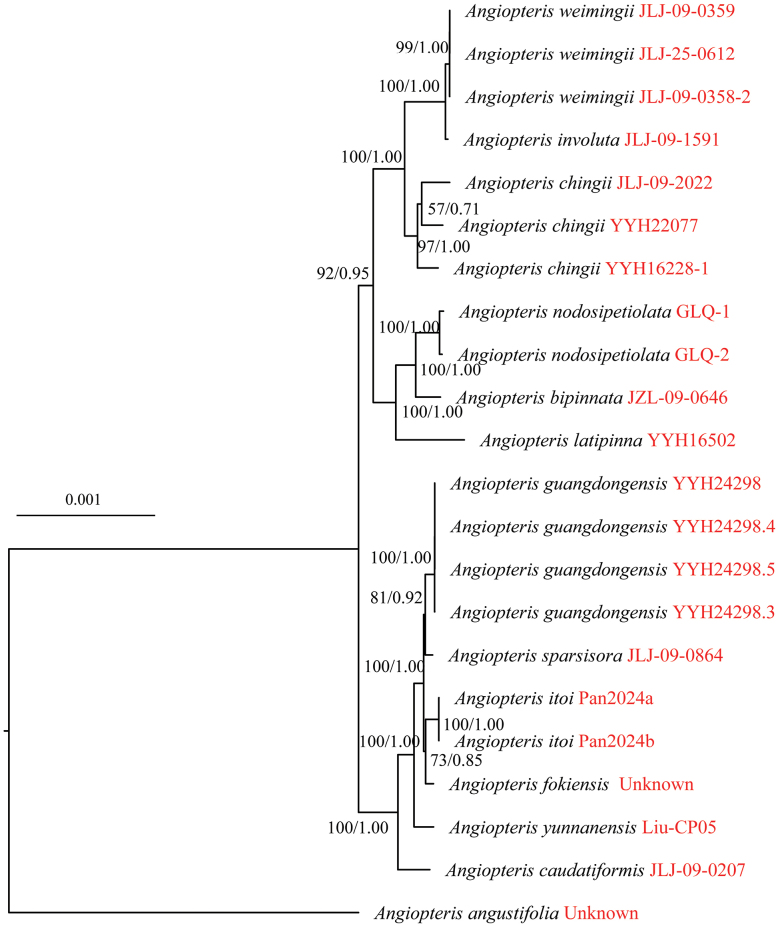 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Species | Plastome size (bp) | IR size (bp) | GC content (%) | Voucher | Location | GenBank ID | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| 153,596 | 89,708 | 20,536 | 21,676 | 35.50 | Unknown | Unknown |
|
| |
| 153,381 | 89,709 | 20,580 | 21,546 | 35.40 | JLJ-09-0646 (HITBC) | Yunnan, China |
|
| |
| 153,137 | 89,710 | 20,585 | 21,421 | 35.40 | JLJ-09-0207 (HITBC) | Yunnan, China |
|
| |
| 153,134 | 89,738 | 20,564 | 21,416 | 35.50 | JLJ-09-2022 (HITBC) | Yunnan, China |
|
| |
| 152,551 | 89,917 | 20,564 | 21,035 | 35.50 | YYH16228-1 | Yunnan, China |
|
| |
| 152,551 | 89,929 | 20,564 | 21,029 | 35.50 | YYH22077 | Yunnan, China |
|
| |
| 153,063 | 89,706 | 20,585 | 21,386 | 35.50 | Unknown | China |
| Unknown | |
| 153,069 | 89, 712 | 20,585 | 21,386 | 35.40 | YYH24298 (NOCC) | Guangdong, China | C_AA110714 |
| |
| 153,069 | 89, 712 | 20,585 | 21,386 | 35.40 | YYH24298.3(NOCC) | Guangdong, China | C_AA110715 |
| |
| 153,069 | 89, 712 | 20,585 | 21,386 | 35.40 | YYH24298.4 (NOCC) | Guangdong, China | C_AA110716 |
| |
| 153,069 | 89, 712 | 20,585 | 21,386 | 35.40 | YYH24298.5 (NOCC) | Guangdong, China | C_AA110717 |
| |
| 153,154 | 89,742 | 20,565 | 21,424 | 35.40 | JLJ-09-1591 ( | Yunnan, China |
|
| |
| 153,379 | 89,712 | 20,585 | 21,541 | 35.40 | Pan2024-a ( | Taiwan, China |
|
| |
| 153,379 | 89,712 | 20,585 | 21,541 | 35.40 | Pan2024-b ( | Taiwan, China |
|
| |
| 153,597 | 89,925 | 20,562 | 21,555 | 35.50 | YYH16502 | Yunnan, China |
|
| |
| 152,964 | 89,931 | 20,563 | 21,235 | 35.50 | GLQ-1 (SWFU) | Yunnan, China |
|
| |
| 152,963 | 89,932 | 20,561 | 21,235 | 35.50 | GLQ-2 (CSH) | Yunnan, China |
|
| |
| 153,067 | 89,709 | 20,586 | 21,386 | 35.40 | JLJ-09-0864 (HITBC) | Yunnan, China |
|
| |
| 153,168 | 89,736 | 20,564 | 21,434 | 35.40 | JLJ-0358-2 (HITBC) | Yunnan, China | C_AA132741 | This study | |
| 153,168 | 89,736 | 20,564 | 21,434 | 35.40 | JLJ-09-0359 ( | Yunnan, China | C_AA132740 | This study | |
| 153,168 | 89,736 | 20,564 | 21,434 | 35.40 | JLJ-25-0612 (HITBC) | Yunnan, China | C_AA132742 | This study | |
| 152,962 | 89,717 | 20,585 | 21,330 | 35.40 | Liu-CP05 (HITBC) | Yunnan (Cult.), China |
|
|
| Characters |
|
|
|
|
|---|---|---|---|---|
| Frond | 120–250 cm | 60–120 cm | 70–160 cm | 50–150 cm |
| Stipe | 45–140 cm, smooth | 35–70 cm, slightly tuberculate | 50–120 cm, smooth | 30–80 cm, smooth |
| Rhizome | erect | ascending | ascending | erect |
| Laminae | Laminae once pinnate to bipinnate, pinnae 8–13 pairs, 8–45 × 2.5–5.5 cm, bases truncate or rounded, rarely cordate, margin crenulate, apex is abruptly acuminate with a short caudate tip (ca. 1 cm). | Laminae once pinnate to bipinnate, pinnae 5–7 pairs, 5–14 × 1.5–3 cm, bases cuneate, margin crenulate. The fertile leaf margins curl upward into a boat shape. | Laminae once pinnate to bipinnate, pinnae 5–12 pairs, 25–35 × 3–5.5 cm, bases cuneate, margin coarsely dentate. | Laminae once pinnate to bipinnate, pinnae 4–9 pairs, 26–34 × 3–5 cm, bases cuneate, margin undulate, apex is acuminate with a long caudate tip (2–4 cm) |
| False veins | present (0.5–2 mm) | absent (or obscure) | absent (or obscure) | obscure or present (longer than sori if present) |
| Sori | sori ca. 2 mm from margin, 0.2–1.5 cm, with 12–120 sporangia | sori ca. 1–2 mm from margin, ca. 0.2–0.8 cm, with 14–60 sporangia | sori ca. 1–2 mm from margin, ca. 0.2–1.2 cm, with 20–80 sporangia | sori medial, ca. 3–5 mm from margin, 0.2–1.2 cm, with 12–90 sporangia |
| Exospores | verrucose ornamentation | tuberculate-spinose ornamentation | spiculose ornamentation | clavate-echinate ornamentation |
| Distribution | Lvchun, Yunnan, China | Jiangcheng, Yunnan, China | Hekou, Yunnan, China | Lianhuachi and Wulai, Taiwan, China |
| References | This study |
|
|
| 1 | Sori short-linear, usually less than 5 mm long, marginal |
|
| – | Sori long-linear, usually more than 5 mm long, medial (on the veinlets) |
|
| 2 | Rhizome creeping. Sori ca. 3–4 mm long, positioned 3–4 mm from margin |
|
| – | Rhizome erect. Sori ca. 1 mm long, positioned 0.5–1 mm from margin |
|
| 3 | Stipe smooth; pinnules lanceolate |
|
| – | Stipe with transversely raised stripes; pinnules oblanceolate, apex abruptly acute with a short caudate tip |
|
| 4 | Rhizome erect |
|
| – | Rhizome ascending |
|
| 5 | Plants height not exceeding 150 cm; pinnae 4–9 pairs, pinnule apex attenuate, false veins absent or present; sori positioned 3–5 mm from margin, mature spores color is white with clavate-echinate ornamentation (Taiwan) |
|
| – | Plants height usually exceeding 150 cm, up to 250 cm; pinnae 8–13 pairs, pinnule apex acute, false veins short (ca. 0.5–2 mm, never reaching soral position); sori positioned 2 mm from margin, mature spores color is yellow with verrucose ornamentation (Yunnan) |
|
| 6 | Plants small, mature fronds usually < 1 m; veins sparse, 5–6 per cm; leaf margins coarsely serrate; sori ca. 4 mm from margin |
|
| – | Plants large, mature fronds > 1 m; veins dense, 8–10 per cm; leaf margins undulate or minutely serrate; sori ca. 1–2 mm from margin |
|
| 7 | Stipes smooth, petiole scales reddish brown; fertile pinnules flat; exospores with spiculose ornamentation |
|
| – | Stipes sparsely tuberculate, petiole scales black or dark brown; fertile pinnules with revolute margins (boat-shaped); exospores with densely tuberculate-spinose ornamentation |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFern and Epiphyte Biology · Plant Diversity and Evolution · Ecology and Vegetation Dynamics Studies
Introduction
Marattiaceae are one of the most ancient lineages of eusporangiate ferns, with extant genera including Angiopteris Hoffm., Christensenia Maxon, Danaea Sm., Eupodium J. Sm., Marattia Sw., and Ptisana Murdock (PPG I 2016). Among these, Angiopteris is one of the most species-rich genera and is widely distributed in the Paleotropics, from Madagascar to the South Pacific islands (He and Christenhusz 2013; PPG I 2016; Zhao et al. 2023; POWO 2025). The taxonomy of this genus has undergone significant revisions, particularly regarding the incorporation of the formerly independent genus Archangiopteris (Li and Lu 2006; Murdock 2008a, b; He and Christenhusz 2013; PPG I 2016; Zhao et al. 2023). Archangiopteris was established by Christ and Giesenhagen (1899) based on diagnostic characters such as simply pinnate fronds, pulvinate stipe articulations, and elongate-linear sori. Despite some uncertainty regarding the robustness of early phylogenetic analyses, molecular phylogenetic studies have consistently confirmed that Archangiopteris is nested within Angiopteris (Li and Lu 2006; Murdock 2008a, b; He 2009). In addition, putative hybrid species—including Angiopteris itoi, A. involuta, A. sparsisora, and A. sugongii—provide critical evidence supporting the broadly defined genus (Ching and Wang 1982; Wu 2002; Wang et al. 2021; Jiang et al. 2025).
Globally, Angiopteris comprises approximately 60 species (POWO 2025), with China serving as a center of diversity and hosting more than 30 species (He and Christenhusz 2013). In China, Yunnan Province harbors 19 recorded species (He and Christenhusz 2013; Wang et al. 2020; Wang et al. 2021; Wang et al. 2024; Jiang et al. 2025). This study describes a new species, Angiopteris weimingii, discovered in Lvchun County, Honghe Prefecture, Yunnan Province, and named in honor of Professor Wei-Ming Chu for his seminal contributions to the study of lycophytes and ferns. This discovery further enriches the species diversity of Angiopteris in Yunnan Province.
Materials and methods
Sample collection and morphological analysis
Living material was first collected from Lvchun County in 2008 (collectors: Guo-Da Tao and Jian-Wu Li; XTBG accession number 0359) and is maintained in the nursery of Xishuangbanna Tropical Botanical Garden (XTBG). Based on Tao’s records and description, we conducted field surveys in May 2024 and June 2025, during which one population comprising approximately 30 plants was recorded.
Both cultivated and wild adult individuals were examined. Voucher specimens are deposited in the Pteridophyte Herbarium of Yunnan University (PYU) and the Xishuangbanna Tropical Botanical Garden (XTBG) Herbarium (HITBC) (herbarium acronyms follow Index Herbariorum by Thiers 2024). Gross morphology was examined, and photographs were taken using an SMZ1270 stereo microscope (Nikon, Japan). Spore materials were attached to carbon adhesive tape under an anatomical lens, after which samples were coated with gold using a BAL-TEC SCD 005 Cool Sputter Coater (BAL-TEC AG., Liechtenstein) and visualized using a QUANTA 200 scanning electron microscope (FEI Co., USA) at 25 kV at Yunnan University, Kunming, China.
Taxonomic sampling, DNA extraction, and sequencing
Three individuals of the new species were sequenced to determine its phylogenetic position. Total genomic DNA was extracted from silica-dried leaf material using the TIANGEN plant genomic DNA extraction kit (TIANGEN Biotech., Beijing, China), following the manufacturer’s protocols, and sequenced at Biomaker Technology Co., Ltd. (Beijing, China) for Illumina sequencing. Paired-end reads of 2 × 150 bp were generated using an Illumina NovaSeq 6000 platform (2G data for each sample). In addition, all 19 available plastomes of Angiopteris from GenBank and GenBase were included. Accession numbers for these sequences are provided in Table 1.
Plastome assembly, annotation, and phylogenetic analyses
Complete plastomes were assembled using GetOrganelle v1.7.1 (Jin et al. 2020). Assembled plastomes were initially annotated using PGA (Qu et al. 2019) to detect and annotate all genes. All tRNAs were confirmed using tRNAscan-SE v2.0.7 (Chan and Lowe 2019). For further validation, the positions of start codons, stop codons, and introns were manually adjusted in Geneious Prime 2019.2.1. Sequences downloaded from GenBank and newly generated data were aligned in Geneious using the MAFFT v7.450 plugin (Katoh and Standley 2013) with the E-INS strategy. Poorly aligned regions were removed using Gblocks v0.91b (Talavera and Castresana 2007). ModelFinder was used to infer the most appropriate nucleotide substitution model based on the corrected Akaike Information Criterion (AICc). Maximum likelihood (ML) bootstrapping was conducted with 5,000 rapid bootstrap (BS) replicates in IQ-TREE v2.1.3 (Nguyen et al. 2015) in a single run. Bayesian inference (BI) was performed using MrBayes v3.2.2 (Ronquist et al. 2012) with two runs of four Markov chain Monte Carlo chains, each beginning with a random tree and sampling one tree every 1,000 generations over 2,000,000 generations. The standard deviation of split frequencies was below 0.001, and the Markov chain Monte Carlo output was examined to confirm convergence and ensure that all effective sample size (ESS) values exceeded 200. Maximum likelihood bootstrap support (ML-BS) values and Bayesian inference posterior probabilities (BI-PP) were generated and visualized using FigTree v1.4.3 (Rambaut 2017).
Results
The complete plastome of the new species displayed a typical quadripartite structure, consisting of a pair of inverted repeats (IRA and IRB), separated by the large single-copy region (LSC) and the small single-copy region (SSC) (Table 1). The overall GC content of the plastome was 35.40%, and the lengths of the LSC, SSC, and IR regions were 89,736 bp, 20,564 bp, and 21,434 bp, respectively (Table 1). The concatenated dataset was 152,766 bp in length, of which 370 sites were parsimony informative. GTR+F+I was selected as the best model of evolution in both ML and BI analyses.
Phylogenetic analyses based on complete plastome datasets revealed generally congruent topologies (Fig. 1). Except for Angiopteris angustifolia, the remaining species of Angiopteris could be divided into two clades: the Angiopteris clade (including Angiopteris caudatiformis, A. fokiensis, A. guangdongensis, A. itoi, A. sparsisora, and A. yunnanensis) and the Archangiopteris clade (including A. bipinnata, A. involuta, A. latipinna, A. nodosipetiolata, and A. weimingii). Within the Angiopteris clade, A. caudatiformis was resolved as sister to the remaining five species in our analysis. Within the Archangiopteris clade, A. involuta and A. weimingii formed a strongly supported monophyletic clade (ML-BS = 99, BI-PP = 1.0) and were sister to A. chingii. These three species together were sister to the clade comprising A. bipinnata, A. latipinna, and A. nodosipetiolata, with strong support (ML-BS = 92, BI-PP = 0.95).
Maximum likelihood tree of Angiopteris weimingii and its putative closely related taxa based on complete plastome sequences. Maximum likelihood bootstrap support (ML-BS) and Bayesian inference posterior probability (BI-PP) values are shown above the branches.
Angiopteris weimingii is morphologically intermediate between the Archangiopteris clade and the Angiopteris clade, characterized by fertile once-pinnate and bipinnate fronds and long linear sori (Fig. 2). It is most morphologically similar to A. itoi from Taiwan Province, which was inferred as a hybrid between A. lygodiifolia and A. somae (Wu 2002), but differs by its taller plants, pinnae apices that are abruptly acuminate with a short caudate tip, sori positioned closer to the leaf margin, and shorter false veins (Fig. 3; Table 2). Despite their morphological similarity, plastid-based phylogenetic analyses revealed that A. weimingii and A. itoi reside in two distinct clades, with A. weimingii placed within the Archangiopteris clade and A. itoi within the Angiopteris clade. Spore morphology also provides a key diagnostic feature for distinguishing A. weimingii from A. itoi (Fig. 2K, M, N, O). The mature spores of A. weimingii are yellow and possess densely verrucate ornamentation, whereas those of A. itoi are white with clavate-echinate ornamentation. This distinct difference in both ornamentation type and spore color constitutes an important diagnostic characteristic. Morphological and molecular evidence collectively supports A. weimingii as a new species.
Angiopteris weimingii L.J.Jiang & Z.R.He. A. Cultivated individual (in XTBG). The red arrow indicates the pulvinate articulation at the stipe in pinnate fronds; B. Pinnate and bipinnate fertile fronds; C. Habitat; D. Pinna of a bipinnate frond; E. Pinnule; F. Leaf margin and retrograde false veins; G. Elongate linear sori; H. Mature sporangia; I. Erect rhizome; J. Pulvinate articulation at the middle-lower part of the stipe in pinnate fronds; K. Spores with densely verrucate ornamentation; L. Black scales on the stipe; M. A. weimingii mature sporangia releasing yellow spores; N. A. itoi spores with clavate-echinate ornamentation; O. A. itoi mature sporangia releasing white spores (materials from Wulai, Taiwan Province).
The type specimens of Angiopteris weimingii (left, PYU) and A. itoi (right, TAIF) clearly exhibit the acute pinna tips of the former, in contrast to the attenuate pinna tips of the latter.
Table 2.: Morphological comparison of Angiopteris weimingii, A. involuta, A. sugongii, and A. itoi
Discussion
Angiopteris weimingii offers novel insights into the taxonomy of Angiopteris. Molecular phylogenetic studies have consistently placed the Archangiopteris clade within Angiopteris (Li and Lu 2006; Wang et al. 2024). Although A. weimingii is morphologically allied with A. itoi, which belongs to the Angiopteris clade, it is nested within the Archangiopteris clade (Fig. 1). This finding supports the taxonomic merger of the two genera and underscores the remarkable evolutionary complexity within Angiopteris. We further hypothesize that A. weimingii may represent a hybrid between Angiopteris and a typical Archangiopteris lineage. Given that both A. sparsisora and A. itoi are presumed to be hybrids (Ching and Wang 1982; Wu 2002), this hypothesis warrants validation through future integrative studies.
Taxonomy
Angiopteris
weimingii
Taxon classificationPlantaeMarattialesMarattiaceae
L.J.Jiang & Z.R.He sp. nov.
6574177C-CBDE-569C-9B18-EE49FDE804A1
urn:lsid:ipni.org:names:77376164-1
Type.
China • Yunnan: Lvchun County, Banpo Township, 26 May 2025, L.J. Jiang & J.P. Shi JLJ-09-0359 (Holotype: PYU JLJ-250526!; Isotype: HITBC C090370!).
Diagnosis.
Angiopteris weimingii (endemic to Yunnan) is most similar morphologically to A. itoi (endemic to Taiwan), but differs in the following key characters: taller plants (up to 2.5 m), pinnule apex with a short caudate tip (ca. 1 cm), sori positioned 2 mm from margin, mature spore color is yellow and possess a densely verrucate ornamentation. In contrast, A. itoi is shorter (ca. 1.5 m), pinnule apex with a long caudate tip (2–4 cm), mature spore color is white with clavate-echinate ornamentation. Phylogenetic analysis revealed that A. weimingii is most closely related to A. involuta, yet A. weimingii is notably larger (often exceeding 1.2 m), with a smooth stipe and retrograde false veins. However, A. involuta is typically under 1.2 m, has tuberculate stipes, lacks retrograde false veins, and possesses consistently enrolled fertile pinnae. Comparative morphology with related species is provided in Table 2.
Description.
Plants terrestrial, 120–250 cm tall. Rhizome stout, erect. Fronds clustered, once pinnate to bipinnate. Stipes 45–140 cm long, terete, 1–1.5 cm in diameter, smooth; densely covered with brown filamentous hairs when young (caducous), sparsely scaly with black, narrowly lanceolate scales (ca. 0.05–0.1 × 0.5–2 cm). Lamina up to 250 cm long. Pinnae once pinnate to bipinnate, pinna bases truncate or rounded, rarely cordate, margins crenate–serrulate and undulate, pinnule apex acute with a short caudate tip (ca. 1 cm). Venation: Veins simple or forked, bearing acroscopic false veins (ca. 0.5–2 mm long, never extending to sori position). Texture herbaceous, adaxially green, abaxially pale green when dry. Sori linear, 0.2–1.5 cm long, composed of 12–120 sporangia, positioned 2 mm from margin. Spores yellow, trilete; exospores densely tuberculate. The spore matures from May to July.
Etymology.
The specific epithet “weimingii” is designated in honor of Prof. Wei-Ming Chu, an esteemed Chinese pteridologist, in recognition of his contributions to the genus Angiopteris. The Chinese name is suggested as ‘维明观音座莲 (wei ming guan yin zuo lian)’.
Distribution and habitat.
Endemic to southern Yunnan (Lvchun County) and grows in rubber plantations understory along streams, alt. 270–350 m.
Conservation status.
The species inhabits the understory of rubber plantations, where anthropogenic disturbance is extremely severe. Only one population (consisting of approximately 30 plants) is found in the wild. In accordance with the IUCN (2024) standards, the species is tentatively designated as Critically Endangered (CR), and further investigations in similar forests are still needed.
Additional specimens examined.
China • Yunnan: Lvchun County, Banpo Township, 26 June 2025, JLJ-09-0358-2 (HITBC!); JLJ-25-0612 (HITBC!).
Keys to the species with fertile fronds in both once-pinnate and bipinnate laminae of Angiopteris in China and surrounding regions
**: **
Supplementary Material
XML Treatment for Angiopteris weimingii
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chan P-P, Lowe TM (2019) t RN Ascan-SE: Searching for t RNA genes in genomic sequences. Methods in Molecular Biology 1962: 1–14. 10.1007/978-1-4939-9173-0_1PMC 676840931020551 · doi ↗ · pubmed ↗
- 2Chen W-F, Sun W-Y, Chen L-J, Shu J-P, Jiang JJ, Zheng Y-B, Yan Y-H (2025) Angiopteris guangdongensis (Marattiaceae): A new species from Guangdong, China. Ecology and Evolution 15(11): e 72447. 10.1002/ece 3.72447 PMC 1259760441221248 · doi ↗ · pubmed ↗
- 3Ching R-C, Wang Z-R (1982) Angiopteris sparsisora Ching, sp. nov.—A putative bigeneric hybrid. Acta Phytotaxonomica Sinica 320: 347–350.
- 4Christ H, Giesenhagen K (1899) Pteridographische Notizen. 1. Archangiopteris nov. gen. Marattiacearum. Flora 86: 72–79.
- 5He Z-R (2009) Systematic study of Angiopteris from China. Yunnan University, 1–106.
- 6He Z-R, Christenhusz MJM (2013) Marattiaceae. In: Wu ZY, Raven PH, Hong DY. (Eds) Flora of China Vol. 2–3. Science Press, Beijing, Missouri Botanical Garden Press, St. Louis, 82–89.
- 7Hsieh TH, Chang YH, Chiou WL, Huang YM (2008) Chromosome numbers of five species of the Marattiaceae in Taiwan. Taiwan Journal of Forest Science 234: 335–339.
- 8IUCN (2024) Guidelines for using the IUCN red list categories and criteria. version 16. https://www.iucnredlist.org/documents/Red List Guidelines.pdf [accessed 17.03. 2025]