Euonymus maotaiensis M.T.An & Xu Wu, sp. nov., a new species of Euonymus (Celastraceae) from southwest China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
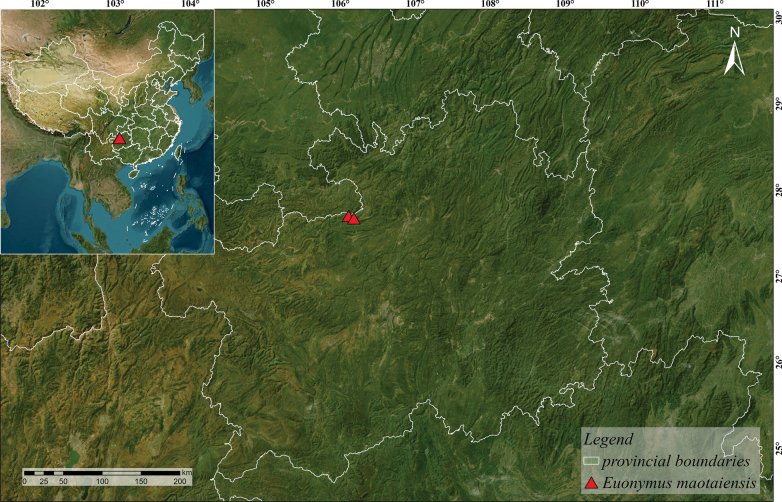 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3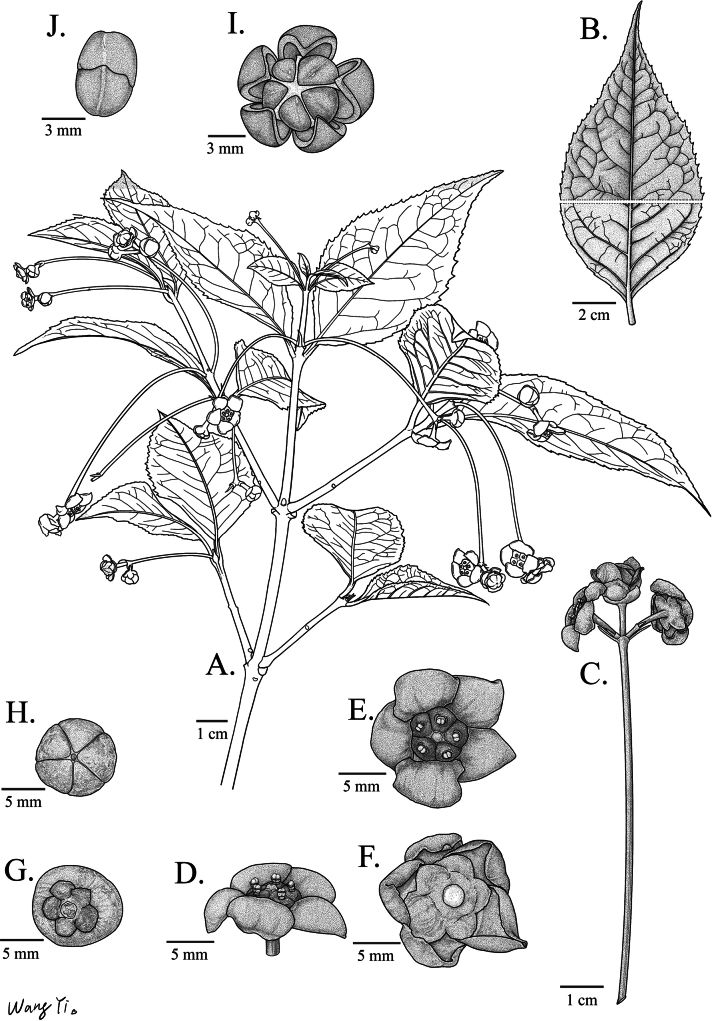 Figure 4
Figure 4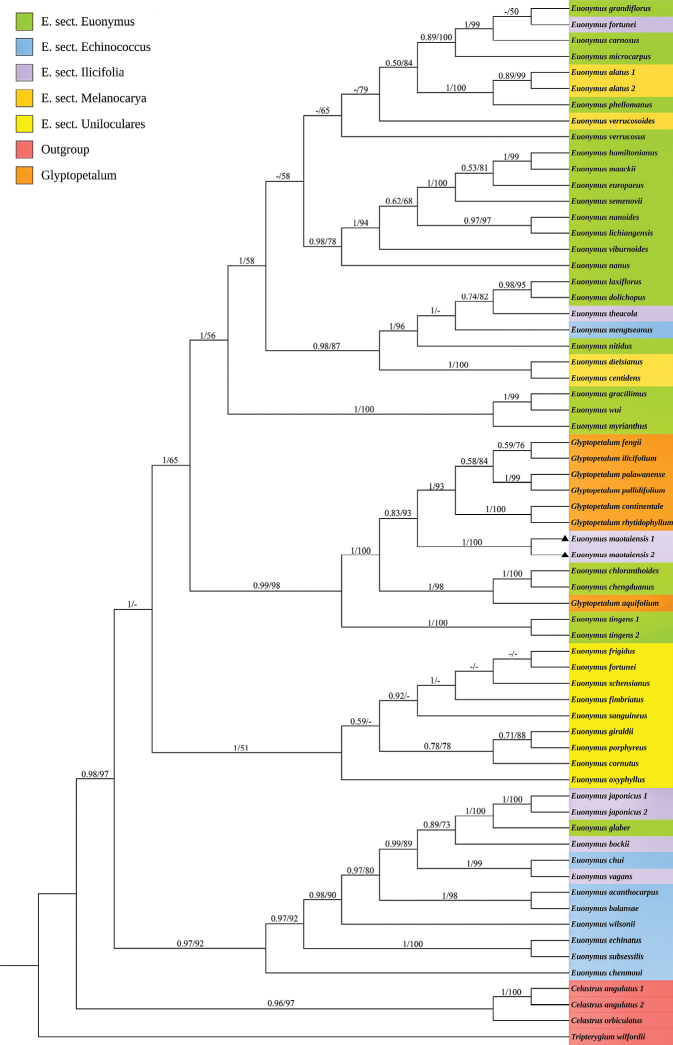 Figure 5
Figure 5| Character |
|
|
|
|
| Branchlets | Slightly 4-angled | With four narrow ridges | Initially 4-angled, becoming terete at maturity | Slightly 4-angled |
| Leaf texture | Rigidly leathery | Thinly leathery | Thinly leathery | Leathery |
| Leaf shape | Broadly lanceolate (rarely lanceolate or elliptic) | Obovate, narrowly oblong-obovate, or elliptic to narrowly elliptic | Narrowly ovate, ovate-elliptic, or ovate-lanceolate | Obovate, elliptic, or oblong-broadly lanceolate |
| Leaf margin | Irregularly sharp serrate | Regularly coarsely spinose-dentate | More or less regularly spinose-dentate | Subentire |
| Petiole length | 5–12 mm | 1–2 mm | 2–3 mm | 6–10 mm |
| Inflorescence | 2–3-flowered (rarely > 10) | 3-flowered | 3–7-flowered | 3–15-flowered |
| Peduncle length | 4–9 cm | 1–2 cm | 1.5–4.5 cm | 2–4 cm |
| Flower merosity | Usually 5-merous (rarely four) | 5-merous | 4-merous | 4-merous |
| Flower color | Purple-black | Purple-black | Deep red or dark red | White or yellowish-green |
| Petals | Broadly ovate, margin recurved, white-membranous, and irregularly undulate | Obovate, margin recurved with shallow irregular teeth | Ovate to broadly ovate, margin revolute, entire | Orbicular or obovate, margin flat, entire |
| Capsule | Globose, indehiscent, shallowly sulcate | 5-lobed, dehiscent to near midpoint | Subglobose, indehiscent, shallowly sulcate | Triangular-ovoid, dehiscent, deeply 4-lobed, or broadly 4-angled |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNatural Compounds in Disease Treatment · Plant Diversity and Evolution · Pharmacological Effects of Medicinal Plants
Introduction
The genus Euonymus Linnaeus (Linnaeus 1753a) belongs to Celastraceae and comprises approximately 130 species that are primarily distributed in the Northern Hemisphere, with centers of diversity in East Asia, the Himalayan Region, and the northern part of South Asia (Blakelock 1951; Ma 2001). China hosts about 98 species that are widely distributed across the country, with more than 54 species endemic to China (Xiong et al. 2024; The Biodiversity Committee of the Chinese Academy of Sciences 2025; Hu et al. 2025; Ma et al. 2025; Qin et al. 2025). At present, Guizhou Province harbors approximately 58 species of the genus Euonymus, exhibiting remarkable diversity (Zhang et al. 2022).
The genus Euonymus is chiefly characterized by opposite leaves, axillary or terminal inflorescences, 4- or 5-merous flowers, stamens inserted on a fleshy annular disk, loculicidally dehiscent capsules, and arillate seeds. However, recent reports of cauliflory in E. fangdingianus (Xiong et al. 2024) have expanded the morphological understanding of the genus. A comprehensive taxonomic system for the genus Euonymus, comprising two subgenera, seven sections, and 14 series, was established by Blakelock (1951) based on characters such as winter bud morphology, aril morphology, and degree of fruit dehiscence. This system has been widely adopted by subsequent researchers (Cheng et al. 1999; Savinov and Baikov 2007). Morphologically, the genus Euonymus shares many traits with the genus Glyptopetalum Thwaites (Thwaites 1856). Phylogenetic analyses also suggest that these two genera form a monophyletic group. Nevertheless, because of limited taxon sampling and the short sequence lengths used in constructing these phylogenies, the results require further validation (Simmons et al. 2012; Li et al. 2014; Hu et al. 2022, 2025).
Renhuai City is located in northwest Guizhou Province, in the middle reaches of the Chishui River and on the northern flank of the West Dalou Mountain (Cheng et al. 2024). Notably, it is the first area in the province to have completed a comprehensive, jurisdiction-wide biodiversity survey. During a botanical survey conducted in October 2024 in Maoba Town and Maotai Town of Renhuai City, we collected this species at the fruiting stage. Its opposite leaves and capsules led to a preliminary identification as a member of Celastraceae. We revisited the site in June of the following year to observe the flowering phase and documented key diagnostic characters. Based on integrated evidence from field observations and detailed examination of voucher specimens, we propose this taxon as a new species of the genus Euonymus.
Materials and methods
Morphological characteristics
Specimens of this species were collected for morphological description and deposited as voucher specimens. Morphological characteristics were observed and measured from living plants. Comparisons with morphologically similar species were based on their type specimens and on morphological descriptions, photographs, and other specimens obtained through the following sources: Flora of China (http://www.efloras.org/), PPBC (https://ppbc.iplant.cn/), CVH (https://www.cvh.ac.cn/), JSTOR Global Plants (https://plants.jstor.org/), and iNaturalist (https://www.inaturalist.org/).
Taxon sampling and phylogenetic inference
The phylogenetic analysis included 64 accessions, representing 59 taxa, including two species of Celastrus Linnaeus (Linnaeus 1753b) and one species of Tripterygium Hook.f. (Bentham and Hooker 1862) as outgroups. Seven species among the 56 entities included in the ingroup were documented as belonging to the genus Glyptopetalum. Information on taxa and GenBank accessions was summarized in Suppl. material 1. The internal transcribed spacer (ITS) sequences were selected for phylogenetic tree analysis, and both maximum likelihood (ML) and Bayesian inference (BI) methods were employed for tree construction. All analyses were performed using PhyloSuite (Zhang et al. 2020). For the BI analysis, the nucleotide substitution model for the data matrix was estimated using ModelFinder (Kalyaanamoorthy et al. 2017) under the corrected Akaike Information Criterion (AICc), and the GTR + I + G model was selected as the best-fit model. The ML analysis was conducted using IQ-TREE v1.4.2 (Nguyen et al. 2014), with the nucleotide substitution model selected via AIC in ModelFinder, resulting in the GTR + G model for the ITS region. The resulting phylogenetic trees were visualized using iTOL (https://itol.embl.de/).
Taxonomic treatment
Euonymus
maotaiensis
Taxon classificationPlantaeCelastralesCelastraceae
M.T.An & Xu Wu sp. nov.
17559297-DB24-5033-AD24-BECB84CAA617
urn:lsid:ipni.org:names:77376161-1
Type.
China. • Guizhou Province: Renhuai City, Maotai, 27°43'N, 106°13'E, alt. 500 m, 26 October 2024, Ming-tai An, Feng Liu, Xu Wu, Jiang-hong Yu, GZAC-WX-2594 (holotype: GZAC!).
Geographical distribution of Euonymus maotaiensis.
Euonymus maotaiensis. A. Habitat; B, C. Plant; D. Karst habitat.
Euonymus maotaiensis. A, D, E. Branches with flowers; B. Branches with fruits; C. Lenticels on the old branches; F. Lenticels on the young branches; G. Serrate of leaf margin; H. Ovules; I. Buds; J. Number of perianth segments; K. Side view of flowers; L. Top view of flowers; M. Bottom view of flowers; N. Immature fruits; O. Top view of mature fruits; P. Bottom view of mature fruits; Q. Anatomical diagram of mature fruit; R. Seeds.
Euonymus maotaiensis. A. Branches with flowers; B. Leaf (adaxial surface above, abaxial surface below); C. Inflorescence; D. Side view of flowers; E. Top view of flowers; F. Bottom view of flowers; G. Top view of fruits; H. Bottom view of fruits; I. Anatomical diagram of fruit; J. Seeds. [Drawn by Yi Wang].
Diagnosis.
E. maotaiensis is morphologically similar to E. chloranthoides but can be distinguished by several key characteristics (Table 1): its broadly lanceolate leaves (as opposed to elliptic to oblong – elliptic), longer petioles (5–12 mm vs. 1–2 mm), longer peduncles (4–9 cm vs. 1–2 cm), 4- or 5-merous flowers (vs. strictly 4-merous), suborbicular petals with the upper half completely recurved abaxially (vs. planar petals), and indehiscent, 4–5 shallowly sulcate globose capsules (vs. 5-lobed capsules dehiscent to near the midpoint).
Table 1.: Morphological comparison between E. maotaiensis and related species, including those distributed in the same area.
Description.
Evergreen shrubs, 1.5–4 m tall, glabrous throughout. Trunk and branches with conspicuous longitudinal lenticels; branchlets slightly 4-angled, green; Leaves opposite, rigidly coriaceous, extremely brittle when dry, usually broadly lanceolate, rarely lanceolate or elliptic, 6–12 × 2–6 cm, apex acuminate, base cuneate to broadly cuneate, margin irregularly sharp serrate; venation conspicuous on both surfaces, midvein raised abaxially, flush to slightly raised adaxially, lateral veins 6–9 pairs, impressed adaxially, prominent abaxially, anastomosing near the margin, reticulate with tertiary veinlets; petiole 0.5–1.2 cm long. Inflorescences mostly cymes with 2–3 flowers, rarely compound thyrses to > 10 flowers, extra-axillary or occasionally axillary; Peduncle elongate, 4–9 cm long; Flowers usually 5-merous, rarely 4-merous, purple-black, ca. 1 cm in diam; Pedicel usually purple-black (occasionally green), 0.5–0.9 cm long; Bracts usually longer than bracteoles, both linear, 0.4–1.1 cm long; Sepals suborbicular, purple-black or green, ca. half as long as petals, margin membranous; Petals suborbicular, purple-black, the upper half of the petal is completely reflexed and obscured from view, margin white-membranous and undulate, irregularly undulate; Staminal filaments Ca. 1 mm long; Pistil styles absent, stigma shortly columnar, disk flat, 4- 5-lobed. ovules erect to pendulous, 2 per locule. Capsule globose, red, 1–2 cm in diam, indehiscent with only 4–5 shallowly sulcate, usually 1 developed per inflorescence; fruiting pedicel slender, green; Seeds 4–5, bright orange-red in vivo.
Distribution and habitat.
This species is distributed in Maotai Town and Maoba Town, Renhuai City, Zunyi City, Guizhou Province (Fig. 1), at an elevation of ca. 500 m. It may be distributed in Sichuan Province and Chongqing City from the PPBC picture. It grows in well-lit broad-leaved forests along river valleys.
Phenology.
Flowering May – June; fruiting October – December.
Etymology.
“maotaiensis” refers to the discovery site of this species.
Local name.
Simplified Chinese: 茅台卫矛.
Conservation status.
We investigated potential habitats in the surrounding area and identified two populations totaling approximately 42 individuals. Euonymus maotaiensis primarily grows along forest edges, particularly on roadside margins, but was not observed in denser understory areas. In addition, a review of specimens and image databases from regions with similar habitats and elevations revealed that photographs taken by Li Xiaodong in Wuhan City, Hubei Province (PPBC id: 22545059–22545070) and those taken by Zhang Hua’an in Xuyong County, Luzhou City, Sichuan Province (PPBC id: 12794935, 12794949) represent E. maotaiensis. However, because a comprehensive investigation of the population status of E. maotaiensis has not yet been conducted, we recommend provisionally classifying it as Data Deficient (DD) according to IUCN criteria (IUCN 2022).
Phylogenetic position analysis
Based on a dataset of ITS sequences, both Bayesian inference (BI) and maximum likelihood (ML) analyses indicate that sequences of the new species form a strongly supported monophyletic clade (BI = 1, ML = 100%). This new species is sister to a supported subclade (BI = 0.83, ML = 93%) containing six species of the genus Glyptopetalum (BI = 1, ML = 93%) (Fig. 5).
Bayesian inference and maximum likelihood phylogenetic tree based on nuclear internal transcribed spacer (ITS) sequences of 64 accessions representing 59 taxa of Euonymus and Glyptopetalum. Values above branches indicate Bayesian posterior probabilities (>0.50) / maximum likelihood bootstrap percentages (>50). Terminal nodes are shown in seven colors corresponding to the five sections of the genus Euonymus and Glyptopetalum described in “Flora of China” (Ma and Funston 2008), as well as the outgroup. Black triangle indicates E. maotaiensis.
Discussion
Morphologically, E. maotaiensis is clearly distinguishable from its close relatives, a conclusion further supported by phylogenetic analyses. The occasional misidentification of E. maotaiensis as E. chloranthoides in the PPBC database underscores their morphological similarity. However, key diagnostic characters evident in PPBC images – such as leaf shape, petiole length, degree of petal recurvature, and degree of capsule dehiscence – allow reliable differentiation between the two species. Nevertheless, it is often difficult to observe all of these distinguishing features simultaneously under field conditions, which can result in errors in identification.
Hu et al. (2022) proposed transferring G. aquifolium (Loes. & Rehder) C.Y. Cheng & Q.S. Ma to the genus Euonymus, reasoning that the genus Glyptopetalum is morphologically close to the genus Euonymus. The original distinction of the genus Glyptopetalum was based on its exclusively 4-merous flowers, a single pendulous ovule per locule of the ovary, and seeds with a branched raphe (Thwaites 1856; Ding Hou 1963; Simmons 2004; Ma and Funston 2008). However, other studies have suggested that Glyptopetalum should be included within a broadly defined genus, Euonymus (Li et al. 2014). Therefore, based on the diagnostic characteristics of E. maotaiensis – including 4- or 5-merous flowers, a flattened disk with non-rolled margins that do not enclose the ovary, and the presence of two ovules per locule – we classify it within the genus Euonymus in this study. Furthermore, based on features such as smooth branchlets without tubercles, 4- or 5-merous flowers, 2-locular anthers, and indehiscent capsules bearing 4–5 shallowly sulcate ribs, we assign it to sect. Euonymus.
Supplementary Material
XML Treatment for Euonymus maotaiensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blakelock RA (1951) A synopsis of the genus Euonymus L. Kew Bulletin 6: 210–290. 10.2307/4120601 · doi ↗
- 2Chen Q, Li DS, An MT, Yu JH, Liu F (2024) Study on the pattern and Influence of species richness of seed plants in Renhuai city, Guizhou Province. Journal of Mountain Agriculture and Biology 43(1): 180–27. 10.15958/j.cnki.sdnyswxb.2024.01.003 · doi ↗
- 3Cheng CY, Ma JS, Huang PH (1999) Euonymus. In: Cheng CY, Huang PH (Еds) Flora Reipublicae Popularis Sinicae (Vol. 45). Science Press, 3–85.
- 4Hou D (1963) Two additional Asiatic species of Glyptopetalum (Celastraceae). Blumea 12: 57–60.
- 5Hu J, Zhang JY, Yu DX, Jiang H, Xu B, Liu Q, He H (2022) Euonymus aquifolium (Celastraceae): Rediscovered in flowering with respect to its taxonomy, nomenclature, and rarity. Phyto Keys 201: 139–151. 10.3897/phytokeys.201.86180 PMC 984888536762312 · doi ↗ · pubmed ↗
- 6Hu J, Huang X, Luo Y, Liu QR, He H, Liu Q (2025) Euonymus chengduanus (Celastraceae), a new species unexpectedly discovered in an urban forest park in the Megacity of Chengdu, West China. Ecology and Evolution 15: e 71234. 10.1002/ece 3.71234 PMC 1198535640225884 · doi ↗ · pubmed ↗
- 7Kalyaanamoorthy S, Minh BQ, Wong TKF, Haeseler A, Jermiin LS (2017) Model Finder: Fast model selection for accurate phylogenetic estimates. Nature Methods 14: 587–589. 10.1038/nmeth.4285 PMC 545324528481363 · doi ↗ · pubmed ↗
- 8Li YN, Xie L, Li JY, Zhang ZX (2014) Phylogeny of Euonymus inferred from molecular and morphological data. Journal of Systematics and Evolution 52(2): 149–160. 10.1111/jse.12068 · doi ↗