Role of miR-101a in targeting Cox-2 to attenuate chondrocyte hypertrophic differentiation and osteoarthritis progression
Rui Mi, Jinnan Chen, Tianxiang Zhu, Huiqin Bian, Rong Wei, Rushuang Deng, Tiaotiao Han, Qian Wang, Yaojuan Lu, Longwei Qiao, Yuting Liang, Qiping Zheng

TL;DR
This study shows that miR-101a reduces chondrocyte hypertrophy and osteoarthritis progression by targeting Cox-2, suggesting it could be a therapeutic target.
Contribution
The novel finding is that miR-101a regulates Col10a1 expression via Cox-2, offering a new therapeutic angle for osteoarthritis.
Findings
miR-101a overexpression decreases Col10a1 and Cox-2 in chondrocytes.
Inhibiting miR-101a increases Col10a1 and Cox-2 expression.
miR-101a mimics reduce articular damage in an osteoarthritis mouse model.
Abstract
MicroRNAs (miRNAs) are small non-coding RNAs that regulate gene expression post-transcriptionally, often playing critical roles in various biological processes. Recent studies have highlighted the involvement of miRNAs in chondrogenesis by targeting key marker genes. Among these, miR-101a has been identified as a significant regulator, previously reported to target cyclooxygenase-2 (Cox-2, ptgs2) in various contexts. Here, we investigate the role of miR-101a in chondrocyte hypertrophy and osteoarthritis (OA) progression, focusing on its regulation of Col10a1 expression. Using multiple web-based tools (TargetScan, PicTar, miRDB, and miRCODE), we identified miR-101a as a potential regulator of Col10a1. Our in vitro experiments demonstrated that miR-101a was down-regulated during chondrocyte hypertrophy in MCT and ATDC5 cells, while Col10a1 and Cox-2 expression levels were up-regulated.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
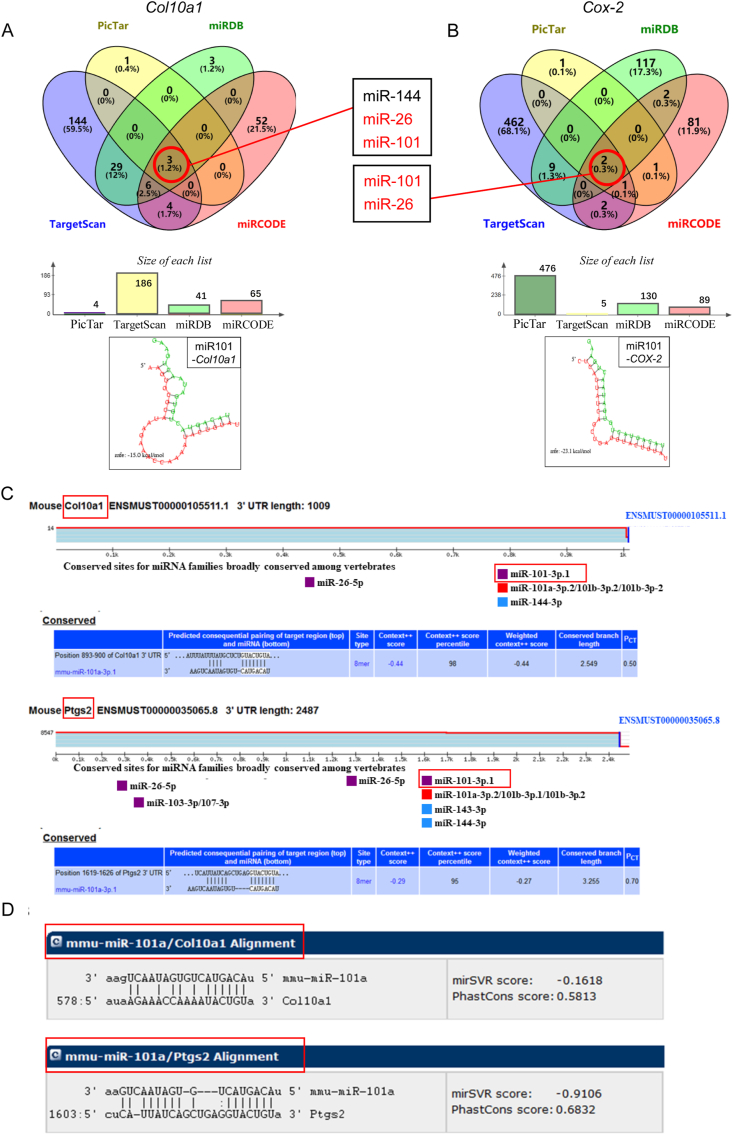 Figure 1
Figure 1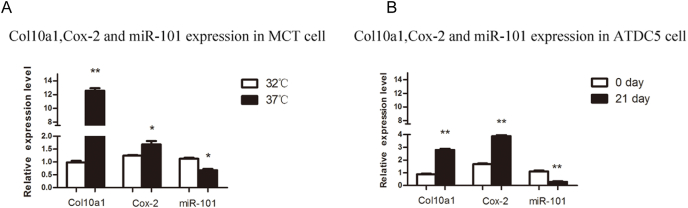 Figure 2
Figure 2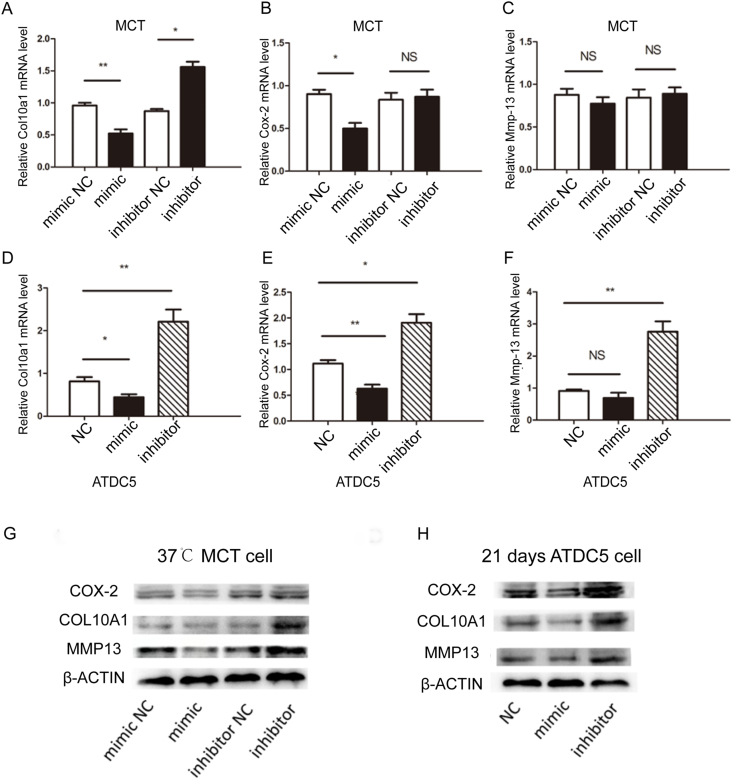 Figure 3
Figure 3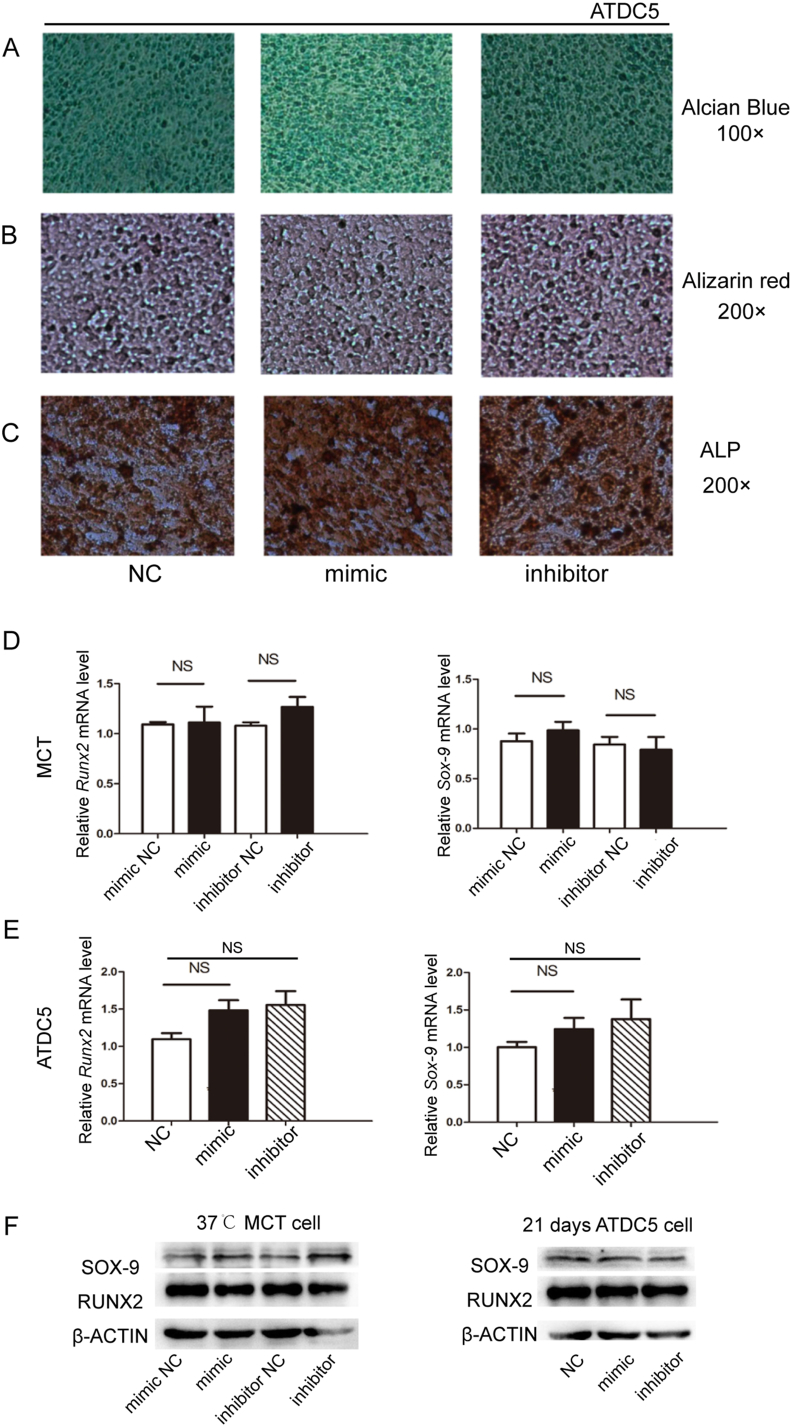 Figure 4
Figure 4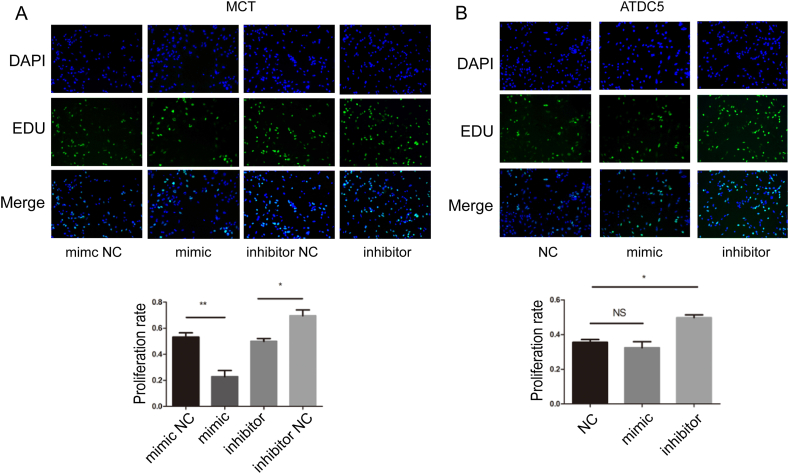 Figure 5
Figure 5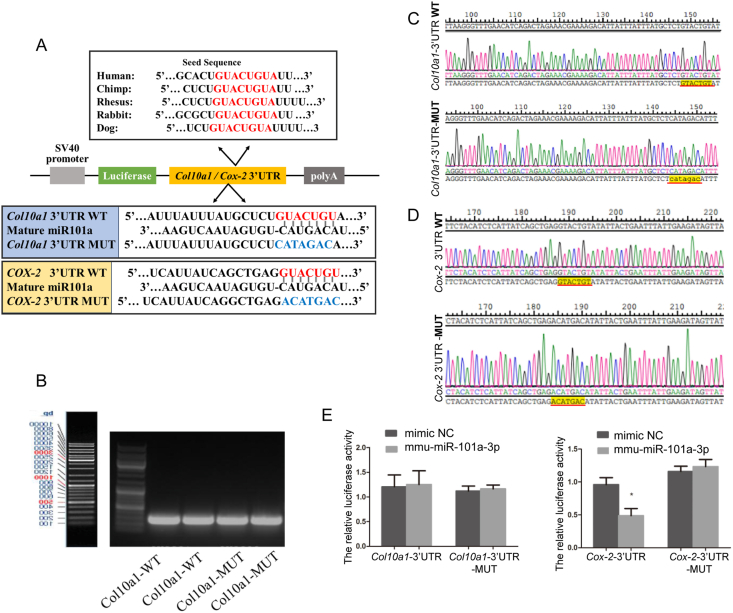 Figure 6
Figure 6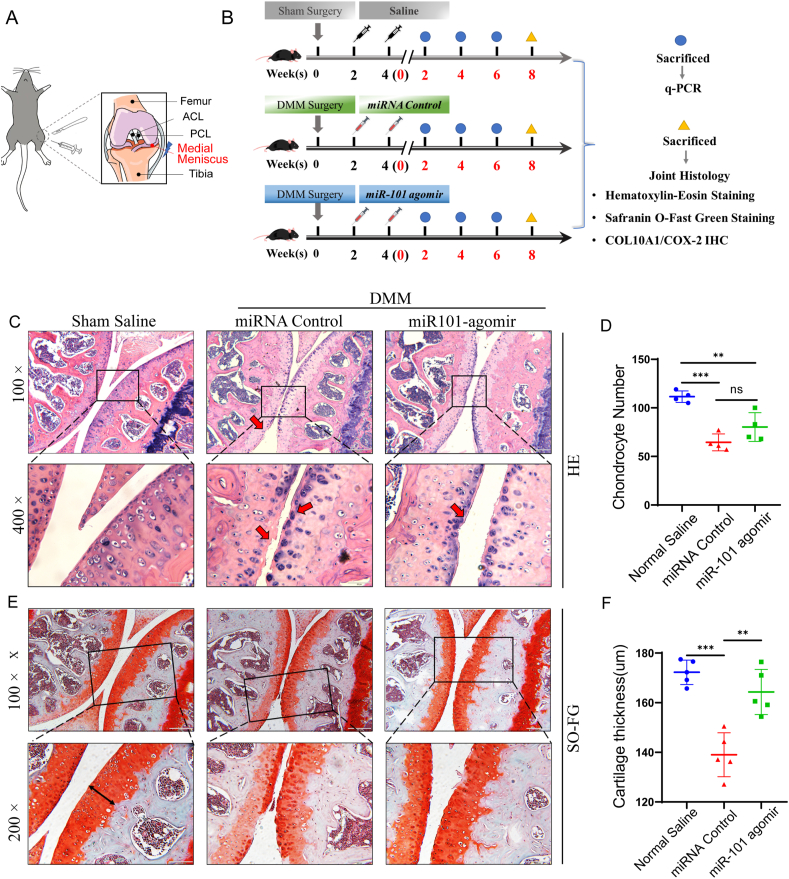 Figure 7
Figure 7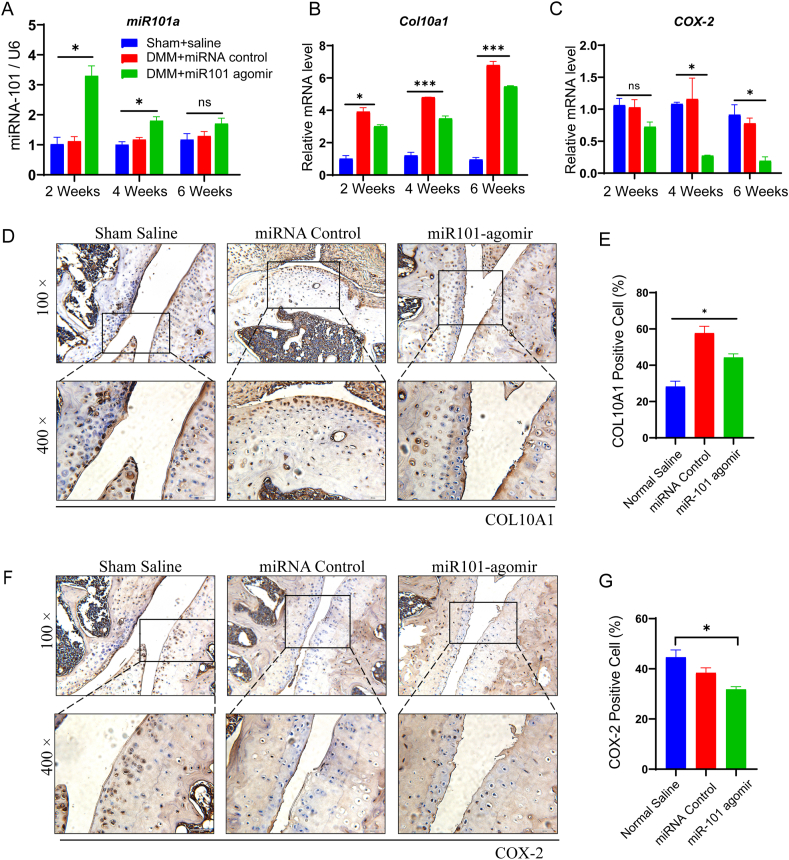 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOsteoarthritis Treatment and Mechanisms · Bone Metabolism and Diseases · Inflammatory mediators and NSAID effects
Introduction
Osteoarthritis (OA) is a prevalent chronic degenerative joint disease, primarily affecting middle-aged and elderly individuals. It is characterized by progressive cartilage degradation, subchondral bone remodeling, and synovial inflammation.1^,^2 A hallmark of OA pathology is the aberrant hypertrophic differentiation of chondrocytes, a process typically associated with the terminal stage of endochondral ossification.3^,^4 This phenotypic shift contributes to extracellular matrix breakdown and cartilage degeneration. Despite its pivotal role in OA progression, chondrocyte hypertrophy remains an under-targeted process in current therapeutic strategies, which predominantly focus on symptom management.5
Among the genes implicated in hypertrophic differentiation, collagen type X alpha 1 (Col10a1) is a well-established marker.6^,^7 Abnormal expression of Col10a1 is associated not only with OA but also with skeletal disorders such as Schmid-type metaphyseal chondrodysplasia, characterized by growth plate abnormalities and skeletal deformities.8 Mechanistic insights into the regulation of Col10a1 are critical not only for understanding OA pathogenesis but also for advancing regenerative approaches using in vitro cartilage models and organoids.9^,^10 Cyclooxygenase-2 (Cox-2, Ptgs2) is another key player in skeletal biology and OA pathology. As an inducible enzyme involved in prostaglandin synthesis, Cox-2 mediates inflammation, fracture healing, and endochondral ossification.11^,^12 Elevated Cox-2 levels in OA cartilage promote extracellular matrix degradation and inflammation via prostaglandin E2 (PGE2) signaling,13^,^14 while Cox-2 knockout models exhibit impaired skeletal repair and reduced expression of transcription factors such as Runt-related transcription factor 2 (Runx2) and specificity protein 7 (Sp7).15
MicroRNAs (miRNAs), a class of ∼21-nucleotide non-coding RNAs, regulate gene expression at the post-transcriptional level and are essential for skeletal development and cartilage homeostasis.16^,^17 Dicer-knockout mice, which are deficient in miRNA biogenesis, show severe skeletal defects, underscoring the crucial role of miRNAs in cartilage formation.18^,^19 Several miRNAs have been identified as regulators of chondrocyte hypertrophy and OA-related pathways. For instance, miR-26a and miR-199a inhibit hypertrophic and inflammatory responses by targeting Col10a1 and Cox-2, respectively.20^,^21 Others, such as miR-140, miR-146a, miR-26b-5p, and miR-27b, modulate key OA-related genes, including matrix metallopeptidase 13 (MMP-13), COL2A1, RUNX2, interleukin-1beta (IL-1β), ADAM metallopeptidase with thrombospondin type 1 motif 5 (ADAMTS-5), and MMP-1.22, 23, 24, 25, 26 miR-101 has emerged as a multifunctional regulator in cartilage and bone biology. Previous studies have shown its involvement in cartilage degradation via the dual specificity phosphatase 1 (DUSP1) pathway in rheumatoid arthritis, modulation of senescence and extracellular matrix breakdown through the LINC00623/miR-101/HRAS axis, and inhibition of SRY-box transcription factor 9 (SOX9), affecting chondrocyte survival and differentiation.27, 28, 29, 30 Moreover, miR-101 influences cartilage integrity via epigenetic regulation of integrin-α1 and can prevent degradation when suppressed in OA models.31^,^32 However, its specific role in regulating chondrocyte hypertrophic differentiation has not been fully elucidated.
In our previous work, we demonstrated that Cox-2 promoted Col10a1 expression and contributed to chondrocyte hypertrophy.33 Given their similar expression profiles in hypertrophic chondrocytes, we hypothesized that both Cox-2 and Col10a1 may be co-regulated by a common miRNA. Through bioinformatics analysis and experimental validation, we identified miR-101a as a direct regulator of Cox-2, indirectly of Col10a1. In vitro, miR-101a levels decreased during hypertrophic differentiation of MCT and ATDC5 cells, while overexpression of miR-101a reduced Cox-2, Col10a1, and MMP-13 expression, without significantly affecting SOX9 and RUNX2. Luciferase reporter assays confirmed Cox-2 as a direct target of miR-101a. Importantly, intra-articular injection of miR-101a mimics in a destabilization of the medial meniscus (DMM)-induced OA mouse model led to reduced Col10a1 expression and attenuation of articular cartilage damage. These findings establish miR-101a as a novel modulator of chondrocyte hypertrophy via direct targeting of Cox-2 and suggest its therapeutic potential in OA intervention.
Materials and methods
Cell lines and cell culture
The mouse cartilage-derived MCT cell line was kindly provided by Dr. B. de Crombrugghe's laboratory at MD Anderson Cancer Center (Houston, Texas, USA), and the mouse cartilage-like ATDC5 was originally from the department of orthopedic surgery at New York University Medical Center. The MCT cells were maintained in Dulbecco's Modified Eagle Medium (DMEM, BI, Israel), supplemented with 8% fetal bovine serum (BI, Israel). The temperature-sensitive MCT cells were cultured at 32 °C (proliferative) in a humidified incubator containing 5% CO_2_ and acquired hypertrophy-like performance when the temperature was up to 37 °C. Mouse cartilage-like ATDC5 cells were cultured in DMEM/F12 (HAM) (1:1) with 5% fetal bovine serum (proliferative) and were treated with 1 × insulin-transferrin-sodium selenite (ITS, Sigma I3146, Germany) to induce hypertrophy.
Overexpression and down-regulation of miRNA
The 5′ FAM chemically modified miR-101a mimics, miR-101a mimics negative control (mimic NC), miR-101a inhibitor, and miR-101a inhibitor negative control (inhibitor NC) were synthesized by GenePharma (Shanghai, China). MCT cells were transfected with 50 nM of either miR-101a mimics, miR-101a inhibitors, or their respective negative controls (mimic NC and inhibitor NC), in 24-well plates using 2 μL Lipofectamine™ 2000 (Invitrogen, USA) according to the manufacturer's protocol. The transfected cells were subjected to 37 °C after 6 h and then cultured in the complete medium for 48 h. To optimize transfection conditions, additional experiments were performed using 40 nM and 80 nM of miR-101a mimics and 80 nM and 120 nM of miR-101a inhibitors. After 6 h of transfection at 37 °C, cells were incubated in complete medium and harvested at 24, 48, or 72 h post-transfection for quantitative reverse transcription PCR or western blotting analysis. Lentiviral vectors containing miR-101a mimics, miR-101a inhibitors, and negative controls were all commercially synthesized by GenePharma (Shanghai, China). ATDC5 cells were transduced with L/miR-101a mimic, L/miR-101a inhibitor, and L/C at a multiplicity of infection (MOI) of 0.6 × 10^8^ TU/mL for 24 h. Cells were maintained in DMEM/F12 medium supplemented with 5% fetal bovine serum and 5 μg/mL streptomycin (Sigma, USA). Colonies were picked up from survival colonies after puromycin selection for 2 weeks. It was confirmed to have integrated with miR-101a mimic and miR-101a inhibitor expression plasmid, and used for subsequent experiments. miR-101a mimic and miR-101a inhibitor stable expression ATDC5 cell line was serially cultured with an optimized concentration of ITS. For time-course validation, stable ATDC5 lines were collected on day 7 and day 21 of ITS-induced differentiation, and subjected to analysis of miR-101a expression and downstream gene regulation.
RNA isolation and quantitative PCR
The total RNA of proliferative and hypertrophic MCT and ATDC5 cells was isolated with TRIzol® reagent (Invitrogen, Thermo Fisher Scientific, Inc., USA) following the manufacturer's instructions. After spectrophotometric quantification, 1 μg of total RNA in a final volume of 20 μL was used for reverse transcription with a PrimeScript™ RT reagent Kit with gDNA Eraser (Perfect Real Time) (TaKaRa, Japan) according to the manufacturer's protocols. Hairpin-it™ miRNAs qPCR Quantitation Kit (GenePharma, Shanghai, China) was used to examine the expression of miR-101a. Reverse transcription quantitative real-time PCR was performed with ultra-SYBR Premix (CWBIO, China) by the Applied Biosystems QuantStudio 5 PCR instrument (ThermoFisher Scientific, USA) according to the thermocycling conditions. β-actin was used as an internal control of mRNA, and U6 was used for miRNA. The 2^–ΔΔCt^ method was used to calculate the fold change of the target genes. Further details of primer sequences used in our study are shown in Table 1.Table 1. Primers for PCR and miRNA fragment.Table 1. List of oligonucleotide sequencesGeneSequence (5′–3′)GapdhACCCAGAAGACTGTGGATGGForwardGapdhCACATTGGGGGTAGGAACACReverseCox-2TGCAGAATTGAAAGCCCTCTForwardCox-2CCCCAAAGATAGCATCTGGAReverseCol10a1TCTGTGAGCTCCATGATTGCForwardCol10a1GCAGCATTACGACCCAAGATCReverseMmp13CTTCTTCTTGTTGAGCTGGACTCForwardMmp13CTGTGGAGGTCACTGTAGACTReverseRunx2ACCCAGCCACCTTTACCTACForwardRunx2TATGGAGTGCTGCTGGTCTGReverseSox9TTCATGAAGATGACCGACGAForwardSox9ATGCACACGGGGAACTTATCReverseU6CTCGCTTCGGCAGCACAForwardU6AACGCTTCACGAATTTGCGTReversemiR-101a-3pCATCGCACGTACAGTACTGTGATAForwardmiR-101a-3pGTGCAGGGTCCGAGGTReversemiRNA agomir fragmentsagomir Negative control (NC)UUUGUACUACACAAAAGUACUGSenseagomir Negative control (NC)CAGUACUUUUGUGUAGUACAAAAnti-senseMmu-miR-101a-3p agomirUACAGUACUGUGAUAACUGAASenseMmu-miR-101a-3p agomirUUCAGUUAUCACAGUACUGUAAnti-sense
Western blotting
Total cellular protein was extracted with radioimmunoprecipitation assay buffer (Beyotime, China) supplemented with 1 × phenylmethylsulfonyl fluoride (PMSF, Beyotime, China), and the supernatant was collected and quantified by a spectrophotometer. An equal amount of protein extracts (100 μL) was separated by 10% SDS-PAGE gel at 100 V for 1.5–2 h and then transferred to the 0.22 μm PVDF membrane (Millipore, Billerica, Massachusetts, USA) at 350 mA for 90 min. The membranes were incubated with specific primary antibodies at 4 °C overnight after blocking with 5% fat-free milk at room temperature for 2 h. The next day, after washing the membrane three times with 1 × Tris-buffered saline with Tween 20 for 15 min, the membrane was incubated with the corroding horseradish peroxidase-labelled secondary antibodies at room temperature for 1 h. Finally, the target bands were visualized by chemiluminescence using a GE Amersham Imager 600. The primary antibodies included COL10A1 (ab182563, Abcam, Massachusetts, USA), COX-2 (D223097, Biotechnology, Shanghai, China), MMP13 (ab51072, Abcam, Cambridge, Massachusetts, USA), RUNX2 (#12556, CST, USA), and SOX9 (#82630, CST, USA), which were prepared with primary antibody dilution (Yeasen Biotechnology, China). β-ACTIN was used as an internal control.
EdU proliferation assay
Proliferative MCT and ATDC5 cells were transfected with miRNA mimics, inhibitors, and controls in 96-well plates. The cells were collected after 48 h and then were detected according to the manufacturer's instructions (Cell Light EdU DNA imaging Kit, Guangzhou, RiboBio, China). Briefly, cells were cultured with 10 μM EdU for 2 h before fixation, permeabilization, and staining.
Alcian blue, alizarin red, and alkaline phosphatase staining
ATDC5 cells from the miR-101a stable line and controls undergoing differentiation were measured by Alcian blue staining to assess the deposition of cartilage matrix proteoglycans. The cells used for Alcian blue staining were fixed with methanol at −20 °C for 2 min. Then, they were stained with 0.1% Alcian blue (Biotechnology, Shanghai, China) overnight. The induced ATDC5 cells were stained with alizarin red to detect the Calcium deposition. Cells were fixed with 95% ethanol for 10 min and then stained with 1% alizarin red. The induced ATDC5 cells were also collected to analyze calcium deposition. Cells were stained according to the manufacturer's instructions for the CAKP kit (Jiancheng, Nanjing, China).
Dual-luciferase reporter gene assay
293T cells were co-transfected with 0.5 μg of either the pirGLO-Col10a1 3′ UTR-WT, pirGLO-Col10a1 3′ UTR-MUT, and 50 nM of either miRNA mimics negative control or miR-101a mimics. Cox-2 plasmids were treated in the same way. After the transfection for 24 h, the obtained cells were used for the luciferase assay and subsequent luminescence calculations. Then, the Firefly and Renilla luciferase were measured with a Dual-Glo luciferase reporter assay system (Promega, E1960, Wisconsin, USA).
Induction of mouse knee OA model and intra-articular injection
Animal experiments were performed under the approval of the Animal Experimentation of Jiangsu University and according to the guidelines of the National Institutes of Health Guide for the Care and Use of Laboratory Animals. C57BL/6 male mice at 4 weeks old were purchased from Changzhou Cavens Laboratory Animal Co., Ltd., Jiangsu, China.
The mouse knee OA model was induced by DMM surgery according to the previously described procedure.34^,^35^,^36 Briefly, the mouse was anesthetized via intraperitoneal injection of sodium pentobarbital (Sigma–Aldrich, St. Louis, Missouri, USA) at a dose of 50 mg/kg, and the surgical procedure was conducted for DMM in their left knee joints. Postoperatively, chlortetracycline ointment was coated on the incision to prevent infection. The timeline for these studies is depicted in Figure 7A. Two weeks after surgery, mice in the DMM group were randomly divided into groups treated with intra-articular injections of miR101a agomir and negative control. The miR101a agomir and negative control are synthesized and shipped as powders, which should be diluted with 15 μL (5 nmol miRNA agomir) saline and completely solubilized in a 37 °C water bath before use.37^,^38 Two intra-articular injections were performed at 2 and 4 weeks postoperatively, the mice were sacrificed 8 weeks after the completion of the injections, and the left knee joint samples were collected and embedded in paraffin for histological analysis.Figure 1. Bioinformatics prediction of interactions between microRNAs and target genes. (A, B) The Venn diagram displaying miR-101a-3p computationally predicted to target Col10a1 and Cox-2 by four different prediction algorithms: TargetScan, PicTar, miRDB, and miRCODE. The structure of the predicted duplex by PicTar was shown. (C) TargetScan bioinformatics prediction result of Col10al and Ptgs2 (Cox-2) was performed. (D) miRanda Bioinformatics prediction result of Col10a1 and Ptgs2 (Cox-2).Figure 1. Figure 2Col10a1, Cox-2, and miR-101a expression in cell lines of chondrocyte hypertrophy. (A) The levels of Col10a1, Cox-2, and miR-101a were analyzed in proliferative (32 °C) and hypertrophic (37 °C) MCT cells. The expression levels of Col10a1 and Cox-2 significantly increased compared with proliferative MCT cells, whereas miR-101a exhibited weak expression during chondrocyte hypertrophy. (B) The expression levels of Col10a1, Cox-2, and miR-101 were shown in the ATDC5 cell line. Both Col10a1 and Cox-2 levels were up-regulated during 21-day 1% ITS induction. However, miR-101 had lower expression in chondrocyte hypertrophy. ∗P < 0.05 and ∗∗P < 0.01.Figure 2. Figure 3miR-101a inhibits hypertrophic differentiation of chondrocytes in vitro. (A–C) Relative mRNA levels of Col10a1, Cox-2, and Mmp13 after transfected with miR101a mimic in ATDC5 cells. (D–F) Relative mRNA levels of Col10a1, Cox-2, and Mmp13 after transfected with miR101a mimic in MCT cells. (G) The protein levels of COL10A1, COX-2, and MMP13 after miR-101a interference in MCT cells were investigated by western blotting. (H) The protein levels of COL10A1, COX-2, and MMP13 after miR-101a interference in hypertrophy-induced ATDC5 cells were detected by western blotting.Figure 3. Figure 4The effects of miR-101a on chondrogenic differentiation in ATDC5 cells. (A) Representative pictures of Alcian blue staining of ATDC5 cells after transfected with miR101a mimic and inhibitor. Magnification, 100×. (B, C) Representative pictures of Alizarin red staining and alkaline phosphatase (ALP) staining of ATDC5 cells after being transfected with miR-101a mimic and inhibitor. Magnification, 200×. (D, E) Reverse transcription quantitative real-time PCR analysis was utilized to evaluate the mRNA expression of Runx2 and Sox-9 in MCT and ATDC5 cells following miR-101a interference. (F) The protein levels of RUNX2 and SOX-9 were assessed by western blotting.Figure 4. Figure 5Overexpression of miR-101a inhibits the proliferation ability of chondrocytes. (A) The representative images of fluorescence and the percentage of EDU-positive cells following the transfection of MCT with miR101a mimic and inhibitor. (B) The representative fluorescent images of ATDC5 cells transfected with miR-101a mimic and inhibitor. The nucleus was labeled with Hoechst 33342 (blue fluorescence), while the proliferative cell was labeled with EdU (green fluorescence).Figure 5. Figure 6Validation of the targeting effect of miR-101a on Col10a1 and Cox-2.(A) Schematic diagram of the WT and MUT luciferase reporter plasmid constructs. The mutation sequences were generated in the Col10a1 and Cox-2 3′ UTR sequence in the complementary site for the seed region of miR-101a, as indicated. The potential binding sequences are indicated in red, while the sites of mutation are shown in blue. (B) The amplification products of Col10a1-miR-101a-WT and Col10a1-miR-101a-MUT were obtained by conventional PCR, and the results of agarose gel electrophoresis showed that the fragment sizes were around 230 bp. (C) Sanger sequencing was utilized to identify the precision of the mutation site on the Col10a1 reporter plasmid. (D) Sanger sequencing was utilized to identify the precision of the mutation site on the Cox-2 reporter plasmid. (E) Luciferase activity of the Col10a1 and Cox-2 3′ UTR reporter was analyzed in 293T cells. ∗P < 0.05.Figure 6. Figure 7Intra-articular injection of miR-101a attenuated OA progression. (A) The diagram illustrates the destabilization of the medial meniscus (DMM) surgical process. Six-week-old male C57BL/6J mice underwent DMM surgery on the medial meniscus of the left hind limb. The medial meniscus tibial ligament, with a portion of the medial meniscus, was dissected by microsurgery. PCL, posterior cruciate ligament; ACL, anterior cruciate ligament. DMM surgery is indicated by blue lightning. (B) The graphic demonstrates the timeline of the postoperative injection procedure of the three groups of mice. Thirty-three model mice were randomly allocated into three groups (n = 11 in each). After three weeks of surgery, mice were treated with equal amounts (15 μL) of normal saline (sham/saline), miRNA control (5 nmol), and miRNA-101 agomir (5 nmol) via intra-articular injection administered through a medial parapatellar approach (the mice in the sham group underwent sham surgery on the right knee). Mice were sacrificed at 2, 4, 6, and 8 weeks after injection for corresponding molecular biological and histological analyses. (C) The hematoxylin and eosin staining of the knee joint coronal sections. Magnification = 100× and scale bar = 200 μm; magnification = 400× and scale bar = 50 μm. (E) The safranin O/fast green staining of the knee joint coronal sections. Magnification = 100× and scale bar = 200 μm; magnification = 200× and scale bar = 100 μm. (D) The statistical count of the chondrocyte number was determined by hematoxylin and eosin staining (thick red arrow: from the cartilage surface to the deep zone). (F) The knee joint cartilage thickness was determined by safranin O/fast green staining (black double arrow: from the cartilage surface to the deep zone). ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001.Figure 7
Histological staining and assessment
The paraffin blocks were cut into 5 μm serial frontal plane sections for the following staining with either hematoxylin, fast green/safranin O, or hematoxylin/eosin. Five different fields of view were used to quantify chondrocytes. The width of cartilage in safranin O/fast green staining slides was measured to operationalize knee joint damage. All the evaluations were independently performed by two researchers.
Statistical analysis
All detections were based on at least three individual experiments. Differences between the groups were determined using the Student's t-test. A one-way analysis of variance (ANOVA) test was used to evaluate whether a gross statistically significant change existed. Data were represented as mean ± standard error of the mean, and P values < 0.05 or < 0.01 were considered statistically significant. All the data were analyzed using GraphPad Prism software version 8.0.
Results
Bioinformatics prediction of a single miRNA that targets Col10a1 and Cox-2
To find a single miRNA that regulates the expression of Col10a1 and Cox-2, four kinds of online bioinformatics approaches were used to screen the miRNAs that could interact with the 3′ UTR of both mRNAs separately (TargetScan, PicTar, miRDB, and miRCODE). Through screening the 3′ UTR region of Col10a1 and Cox-2 from four miRNA target databases, we identified two overlapping candidate miRNAs, miR101 and miR26, and the structure of the predicted duplex by PicTar was shown (Fig. 1A and B). miR-26a was proven to regulate Col10a1 gene expression in previous studies. miR-26a could directly bind to and inhibit the expression of Col10a1 in primary epiphyseal chondrocytes and manipulate chondrocyte hypertrophy and the cartilage extracellular matrix composition.20 We further confirmed the binding site of miR101 with the 3′ UTR of Col10a1 and *Cox-*2 by TargetScan and miRanda, and the results were consistent with the predicted site of Pictar, and there existed complementary sites in the seed region of miR-101a with Cox-2 and Col10a1 (Fig. 1C and D).
Expression levels of miR-101a, Col10a1, and Cox-2 in proliferative and hypertrophic chondrocyte models
The MCT cells were mouse chondrocytes that were immortalized with a temperature-sensitive simian virus 40 large tumor antigen.39 The MCT cells proliferated continuously when cultured at 32 °C, while the culture temperature was raised to 37 °C, the cell proliferation rate decreased, and the cells underwent a hypertrophic differentiation phase with specific expression of the collagen type X gene. In response to changes in culture temperature, MCT cells can mimic the differentiation process of chondrocytes in vitro and are a classic cell model for in vitro studies of chondrocyte differentiation and maturation. The expression changes of miR-101a, Col10a1, and Cox-2 both in the proliferative (32 °C) and hypertrophic (37 °C) stages were detected by quantitative PCR. The results showed that the Col10a1 mRNA level was higher in hypertrophic differentiation compared with proliferative MCT cells. Furthermore, *Cox-*2 mRNA is two-fold higher in hypertrophic than in proliferative. On the contrary, miR-101a approximately falls by half in the hypertrophic period (Fig. 2A).
The ATDC5 cells, derived from the teratocarcinoma, are a common mouse chondrocyte cell line that could mimic the sequential process of endochondral ossification under the prolonged simulation culture of chondrogenic-induced medium containing ITS (insulin, transferrin, and sodium selenite).34^,^40^,^41 We further detect the expression of miR-101a, Col10a1, and Cox-2 in ATDC5 cells with (ITS supplement, cultured for 21 days) or without (cultured for 0 day) hypertrophic-induced medium. The results indicated that the mRNA levels of Col10a1 and Cox-2 were both up-regulated 3-to-4-fold in hypertrophic ATDC5 cells compared with the proliferative ATDC5 cells. Meanwhile, the level of miR-101a was significantly decreased in hypertrophic chondrocytes (Fig. 2B). Collectively, these data suggest that miR-101a could be a potential negative regulator of Col10a1 and Cox-2.
miR-101a inhibits the expression of Col10a1 and Cox-2 both in MCT and ATDC5 cells
To confirm the effect of miR-101a on chondrocyte hypertrophy, three hypertrophic chondrocyte-related markers, Col10a1, Cox-2, and Mmp-13, were analyzed by quantitative PCR and western blotting after transfection with miR-101a mimic, miR-101a inhibitor, or corresponding negative controls. The results showed that the levels of Cox-2 and Col10a1 were significantly reduced after overexpression of miR-101a compared with the negative control in MCT and ATDC5 cells (Fig. 3A, B, D, E). However, there was no significant change in Mmp-13 in MCT and ATDC5 cells (Fig. 3C–F). Meanwhile, silencing miR-101a in cell models of chondrocyte hypertrophy up-regulated Col10a1 expression in MCT and ATDC5 cells. We also found that Cox-2 and Mmp-13 had high expression in ATDC5 cells, but no significant changes were found in Cox-2 and Mmp-13 in MCT cells (Fig. 3B–E). Western blotting results confirmed that miR-101a overexpression down-regulated the level of COL10A1, COX-2, and MMP13 (Fig. 3G and H). To support the reliability of these transfection experiments, we performed optimization of transfection concentrations for miR-101a mimic/inhibitor and a time-course analysis of miR-101a expression, as shown in Figure S1–S3. Together with the mRNA level, the results suggest that miR-101a could suppress the expression of Col10a1, Cox-2, and Mmp-13 during chondrocyte hypertrophy.
The effects of miR-101a on chondrogenic differentiation in ATDC5 cells
To further explore the influence of miR-101a on chondrocyte development, we performed three kinds of staining experiments on ATDC5 cells and quantitative reverse transcription PCR on several relevant marker genes during chondrocyte differentiation in MCT and ATDC5 cells.
As illustrated in Figure 4A, the intensity of Alcian blue staining was weaker in the miR-101a overexpression group than in the NC group, but no significant difference was observed in miR-101a-silencing ATDC5 cells, implying the partially positive role of miR-101a in chondrocyte differentiation and maturation. Nevertheless, no clear difference was observed in the intensity of Alizarin red staining and alkaline phosphatase staining between the experimental and control groups, indicating a limited role of miR-101a in chondrocyte proliferation at the early stage and matrix mineralization at the late stage of endochondral ossification (Fig. 4B and C). To further characterize the effect of miR-101a on chondrogenesis, we detected the expression of related transcription factors Runx2 and Sox9 by quantitative reverse transcription PCR and western blotting. However, there was no significant effect of miR-101a on Runx2 and Sox9 in hypertrophic MCT and ATDC5 cells induced by ITS for 21 days (Fig. 4D–F).
Overexpression of miR-101a inhibits the proliferation ability of chondrocytes
We performed EdU assays to investigate the effect of miR-101a on the proliferation of chondrocytes. MCT and ATDC5 cells were seeded in 96-well plates at a density of 2.5 × 10^3^ cells per well. After 24 h, the cells were transfected with miR-101a mimics, miR-101a inhibitors, and negative controls separately. Transfection reagents were removed after 6 h, and cells were cultured for another 24 h in DMEM with 8% fetal bovine serum. The fluorescence results after merging EDU with DAPI indicated that forced miR-101a expression in MCT cells suppressed their proliferation, while inhibition of miR-101a expression facilitated the proliferation ability in chondrocytes (Fig. 5A), which is consistent with the results of Alcian blue staining. In addition, we also examined EdU cell proliferation in ATDC5 cells. After transient transfection with miR-101a inhibitor, it was able to partially promote the proliferation rate of ATDC5 cells, with no significant change after transfection with miR-101a mimic (Fig. 5B). Taken together, we found that miR-101a may have a role in inhibiting chondrocyte proliferation in vitro.
Analysis and verification of miR-101a target sites
To investigate the specific mechanism of miR-101a between Col10a1 and Cox-2, the dual-luciferase reporter assay was conducted. The potential binding site of miR101a with Col10a1 and Cox-2 predicted by TargetScan and the corresponding mutation sequences are shown in Figure 6A. Agarose gel electrophoresis was used to determine the size of the DNA fragments after PCR, and the fragment sizes were all located at 230 bp (Fig. 6B). After digesting and ligating the wild-type (WT) and mutation fragments to the pmirGLO vector, Sanger sequencing was used to confirm that the Col10a1 and Cox-2 vectors were constructed correctly and used for subsequent experiments (Fig. 6C and D). The reporter plasmid of pmirGLO-Col10a1-3′ UTR-WT and pmirGLO-Col10a1-3′ UTR-MUT was co-transfected with miR-101a mimic and mimic NC, respectively. The activity of luciferase of 293T cells was significantly reduced after being co-transfected with pmirGLO-Cox-2-3′ UTR-WT and miR101a mimic compared with the mutation vector. However, there was no significant difference in luciferase activity after transfection of pmirGLO-Col10a1-3′ UTR-WT and miR101a mimic (Fig. 6E). Together, the reporter assay showed that miR-101 could directly target the 3′ UTR region of Cox-2.
Intra-articular injection of miR-101a attenuated OA progression
miRNA agomir and antagomir are chemically modified miRNA agonists and antagonists, respectively, which can be used to regulate target gene mRNA expression in vivo. Compared with common miRNA mimics and inhibitors, miRNA agomir and antagomir have higher stability and miRNA activity in animals, and are more likely to be enriched in target cells through cell membranes and tissue interstitial spaces. They can be administered systemically or locally, and their effects can last for several weeks.37^,^38
Figure 7A and B show the schematic and the timeline of the DMM surgery. The surgery was performed on the left knee joint of four-week-old C57BL/6 male mice, followed by intra-articular injections of 15 μL miR101a-agomir and control solution every 2 weeks after surgery. The mouse knee joint was collected at 8 weeks after the completion of the second injection for subsequent histological and molecular biological analysis. The hematoxylin and eosin staining of knee joint slides showed that the width of joint space became narrowed in the DMM group (Fig. 7C). The number of chondrocytes was significantly reduced in the DMM group (Fig. 7D). The safranin O/fast green staining and the statistical analysis of cartilage thickness illustrated that the cartilage damage and loss were most severe in the DMM group compared with the miRNA control group, and miR101-agomir supplement could partially alleviate cartilage damage (Fig. 7E and F).
miR-101a inhibits Col10a1 and Cox-2 expression in the knee joints of DMM-induced OA mice
According to the surgical timeline settings shown in Figure 7A, the knee joint tissues were harvested at 2 weeks, 4 weeks, and 6 weeks after the second injection, respectively. Quantitative PCR analysis and immunohistochemical staining were conducted to assess the expression level of Col10a1 and Cox-2. The mRNA level of miR101a was increased in the first 4 weeks after injection (Fig. 8A). The mRNA expression of Col10a1 in the DMM group was extremely evaluated and the addition of miR101-agomir partially decreased the level of Col10a1 (Fig. 8B). The mRNA of Cox-2 did not show significant differences between the DMM and sham groups, however, miR101-agomir could inhibit the expression of Cox-2 significantly (Fig. 8C). Meanwhile, the immunohistochemical staining of COL10A1 showed an increased protein level in the DMM group compared with the sham group, and the proportion of positive cells in the superficial layer of articular cartilage in the group with intra-articular injection of miR101-agomir was decreased than that in the miRNA-agomir control group (Fig. 8D and E). However, compared with the sham group, there was a decrease in the protein level of COX-2 in the DMM group, and the injection of agomir had no significance between the miR-101a and negative control groups (Fig. 8F and G). The immunohistochemical staining and the proportion of the positive cells in articular cartilage tissues of OA mice suggest that miR-101a could inhibit the degradation of type X collagen.Figure 8. Col10a1, COX-2, and miRNA-101a expression after intra-articular injection of miRNA-101 agomir in DMM-induced OA mice. (A–C) The relative mRNA expression levels of miRNA-101, Col10a1, and Cox-2 in mouse cartilage at 2, 4, and 6 weeks after intra-articular injection of miRNA-101 agomir (n = 2 in each group). β-actin and U6 were used as endogenous controls. (D) The immunohistochemical staining of COL10A1 in the articular cartilage tissue of the knee joint. (E) Quantification of the percentage of COL10A1-positive cells in the three groups as determined by the staining results. (F) The immunohistochemical staining of COX-2 in the articular cartilage tissue of the knee joint (n = 4 in each group). Magnification, 100 × or 400 × . (G) Quantification of the percentage of COX-2 positive cells in the three groups as determined by the staining results. ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001.Figure 8
Discussion
The process of cartilage development and homeostasis is tightly regulated by a coordinated network of transcription factors, signaling molecules, and epigenetic regulators. Among these, Cox-2 and Col10a1 play essential roles during chondrocyte hypertrophy.24 While the epigenetic regulation of hypertrophy-related genes by miRNAs has been increasingly recognized, specific interactions remain underexplored. For instance, although miR-101a has previously been shown to regulate extracellular matrix-degrading genes and Sox9 in cartilage,32 its role in modulating hypertrophic differentiation has not been well defined. In this study, we explored how miR-101a influenced hypertrophic chondrocyte marker expression. We observed that miR-101a expression was down-regulated during the hypertrophic stage in MCT and ATDC5 cells, while Cox-2 and Col10a1 were significantly up-regulated. Overexpression of miR-101a suppressed both Cox-2 and Col10a1, along with Mmp-13, an extracellular matrix-degrading enzyme involved in OA progression.42 These results suggest that miR-101a acts as a negative regulator of chondrocyte hypertrophy.
Interestingly, miR-101a did not significantly affect Runx2 or Sox9 expression in our system, suggesting that it may act upstream of or parallel to these master transcription factors. Given that each miRNA can target hundreds of mRNAs,43 the context-specific regulation of miR-101a is not surprising. Previous studies in OA models showed that miR-101a suppressed chondrocyte proliferation,32^,^44 consistent with our incorporation results of Alcian blue and EdU stainings, which indicate reduced proliferation upon miR-101a overexpression. These findings support a model in which miR-101a may exert distinct regulatory effects at different stages of chondrocyte development. To determine direct molecular interactions, we conducted luciferase reporter assays and confirmed Cox-2 as a direct target of miR-101a. Previous work in other cell types has shown that miR-101a represses Cox-2 expression, including in mammary gland epithelial cells, colon cancer, and esophageal squamous-cell carcinoma cell lines.45^,^46 Our findings extend this regulatory relationship to chondrocytes and further link it to hypertrophy.
Mechanistically, our data suggest that miR-101a may delay chondrocyte hypertrophy by targeting Cox-2, which plays a key role in prostaglandin E2 (PGE2) synthesis. PGE2 signaling has been implicated in promoting hypertrophic differentiation via Runx2 activation. Notably, Cox-2 has been shown to act as a co-activator of Runx2, enhancing its transcriptional activity and thereby up-regulating Col10a1 expression.47^,^48 Moreover, Cox-2 activity is essential for bone morphogenetic protein 2 (BMP-2)-mediated hypertrophy, further supporting its role in this pathway. Therefore, miR-101a-mediated suppression of Cox-2 could reduce PGE2 levels, inhibit Runx2 activity, and subsequently down-regulate Col10a1. This highlights a potential miR-101a–Cox-2–PGE2–Runx2–Col10a1 axis that regulates hypertrophic differentiation. Future studies using pathway inhibitors or genetic manipulation of downstream targets will be important to confirm this regulatory network. Although our findings support the therapeutic potential of miR-101a in OA, the clinical translation of miRNA-based therapies remains challenging. A recent study showed that intra-articular delivery of miR-15a promoted chondrocyte apoptosis in OA mice by targeting B-cell lymphoma 2 (BCL-2), illustrating both the potential and complexity of miRNA-based interventions.49 In our model, intra-articular injection of miR-101a mimics resulted in reduced Col10a1 expression and cartilage degradation, suggesting a protective role in OA.
We acknowledge several limitations in our study. First, the lack of a positive control drug precludes a direct assessment of miR-101a′s therapeutic efficacy. Second, although micro–computed tomography (micro-CT) offers a non-destructive method to quantify cartilage volume, subchondral bone remodeling, and osteophyte formation in OA models,50 we were unable to perform this analysis because the original animal studies had already concluded and the samples had subsequently degraded. To address these limitations, future studies will include both micro-CT imaging and a positive drug control to rigorously validate the therapeutic potential of miR-101a. In parallel, we are investigating the mechanistic role of miR-101a in BMP-2-induced C2C12 cells, a well-established model of osteochondral differentiation. These complementary studies will clarify whether miR-101a exerts broader regulatory functions across distinct stages and models of cartilage development and degeneration (supplemental data and data not shown).
Conclusions
In summary, our findings identify miR-101a as a novel post-transcriptional regulator of chondrocyte hypertrophic differentiation through direct targeting of Cox-2, leading to downstream suppression of Col10a1 expression. This miR-101a–Cox-2–Col10a1 axis may play a critical role in delaying hypertrophy and attenuating OA cartilage degeneration. While further studies are needed to delineate the precise downstream signaling mechanisms and validate therapeutic efficacy in more comprehensive models, our study provides new insight into the epigenetic regulation of OA progression and highlights miR-101a as a promising target for future therapeutic development.
CRediT authorship contribution statement
Rui Mi: Writing – original draft, Conceptualization, Data curation. Jinnan Chen: Methodology, Data curation, Writing – original draft, Formal analysis, Conceptualization. Tianxiang Zhu: Formal analysis, Methodology. Huiqin Bian: Data curation, Methodology. Rong Wei: Writing – review & editing. Rushuang Deng: Writing – review & editing. Tiaotiao Han: Methodology. Qian Wang: Data curation. Yaojuan Lu: Writing – review & editing. Longwei Qiao: Writing – review & editing. Yuting Liang: Writing – original draft. Qiping Zheng: Writing – review & editing, Investigation, Conceptualization, Supervision, Funding acquisition.
Data availability
All data generated or analyzed during this study are included in this published article.
Ethics declaration
All experiments and methods were performed according to relevant guidelines and regulations. All animal procedures were approved by the Institutional Animal Care and Use Committee of Jiangsu University.
Funding
This work was supported by the 10.13039/501100013058Jiangsu Provincial Key Research and Development Program (No. BE2020679 to Q.Z.), the Innovation Team (leader) of Jiangsu Province, China (2017, to Q.Z.), Science Foundation of Jiangsu Province (China) (No. BK20240371 to Y.L.), and the National Science Foundation of China (No. 81901632 to Y.L.; 82001576 to L.Q.).
Conflict of interests
The authors declared no conflict of interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abramoff B.Caldera F.E.Osteoarthritis: pathology, diagnosis, and treatment options Med Clin North Am 104220202933113203557010.1016/j.mcna.2019.10.007 · doi ↗ · pubmed ↗
- 2Allen K.D.Huffman K.Cleveland R.J.Evaluating osteoarthritis management programs: outcome domain recommendations from the OARSI joint effort initiative Osteoarthr Cartil 317202395496510.1016/j.joca.2023.02.078PMC 1056583936893979 · doi ↗ · pubmed ↗
- 3Cancedda R.Cancedda F.D.Castagnola P.Chondrocyte differentiation International Review of Cytology 1995 Elsevier Amsterdam 265358
- 4Bijlsma J.W.J.Berenbaum F.Lafeber F.P.J.G.Osteoarthritis: an update with relevance for clinical practice Lancet 37797832011211521262168438210.1016/S 0140-6736(11)60243-2 · doi ↗ · pubmed ↗
- 5Bálint G.Szebenyi B.Diagnosis of osteoarthritis. Guidelines and current pitfalls Drugs 52Suppl 3199611310.2165/00003495-199600523-000038911794 · doi ↗ · pubmed ↗
- 6Kawaguchi H.Endochondral ossification signals in cartilage degradation during osteoarthritis progression in experimental mouse models Mol Cells 25120081618319608 · pubmed ↗
- 7Bonaventure J.Chaminade F.Maroteaux P.Mutations in three subdomains of the carboxy-terminal region of collagen type X account for most of the Schmid metaphyseal dysplasias Hum Genet 96119955864760765510.1007/BF 00214187 · doi ↗ · pubmed ↗
- 8Allen D.B.Cuttler L.Clinical practice. Short stature in childhood: challenges and choices N Engl J Med 368132013122012282353456110.1056/NEJ Mcp 1213178 PMC 5754004 · doi ↗ · pubmed ↗