Divergent volatile metabolomes and flavor attributes in rice fermented by Aspergillus oryzae and Aspergillus flavus
Colin O. McCarthy, Dasol Choi, Alissa A. Nolden, Eric A. Decker, Jae-Hyuk Yu, John G. Gibbons

TL;DR
This study compares how two types of fungi, Aspergillus oryzae and Aspergillus flavus, affect the flavor and chemical makeup of fermented rice.
Contribution
The study reveals how domestication has shaped the volatile compound production of A. oryzae to enhance sensory food qualities.
Findings
A. oryzae RIB40 produced more volatile compounds linked to pleasant flavors like fruity and umami.
A. oryzae RIB40 had higher alpha-amylase activity and lower bitterness and astringency in fermented rice.
Aflatoxin was detected only in A. flavus NPK13tox, not in A. oryzae.
Abstract
The generally recognized as safe (GRAS) fungus Aspergillus oryzae has been used for millennia in the production of traditional Asian fermented foods and beverages. Domestication has led to genomic and phenotypic adaptations that distinguish A. oryzae from its wild relative, Aspergillus flavus. While differences between these species have been partially characterized, their comparative production of volatile compounds during food fermentation remains poorly understood. We evaluated alpha-amylase activity, aflatoxin production, taste attributes using an electronic tongue, and volatile profiles using dynamic headspace gas chromatography-mass spectrometry. Analyses were conducted during rice fermentation using the food-grade strain A. oryzae RIB40 and two wild A. flavus strains, NPK13tox and AflaGuard. A. oryzae RIB40 exhibited significantly higher alpha-amylase activity during rice…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
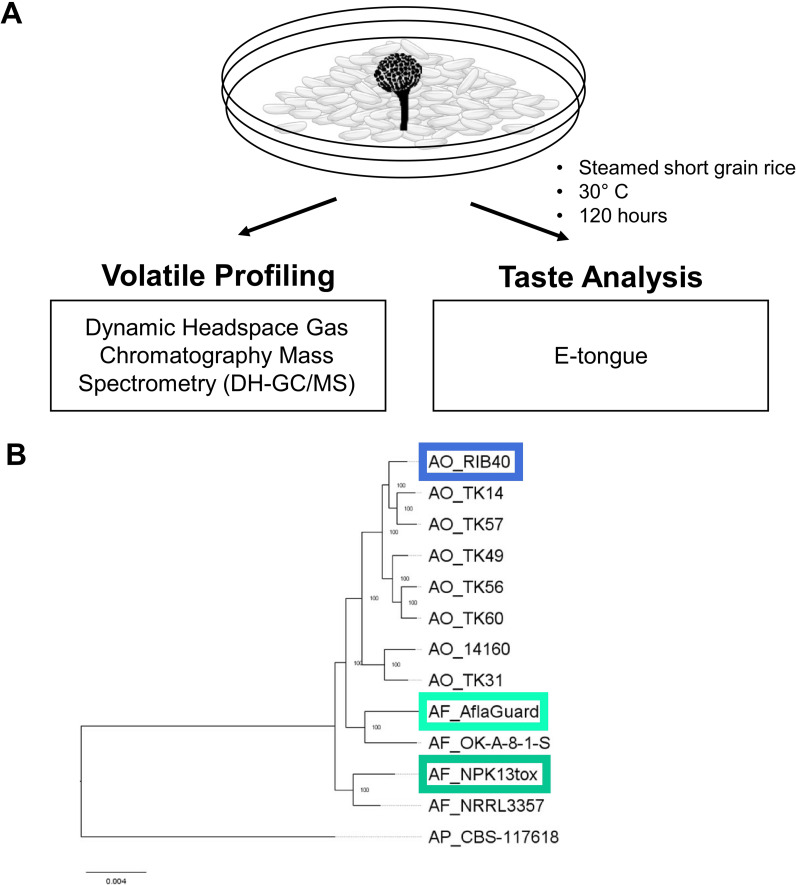 Figure 1
Figure 1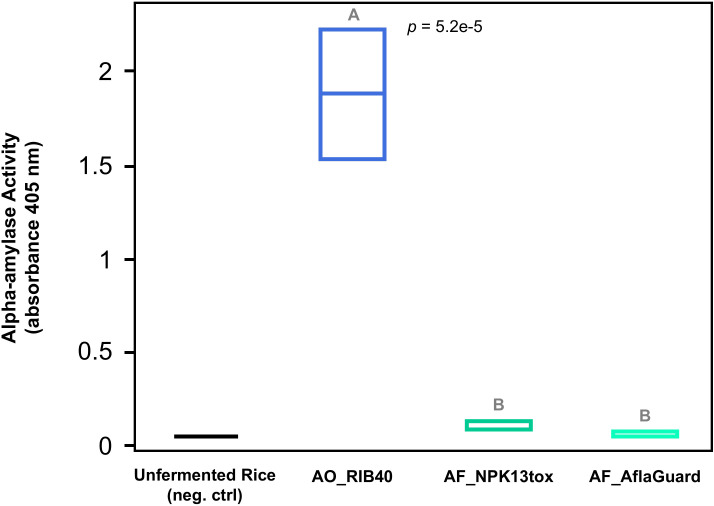 Figure 2
Figure 2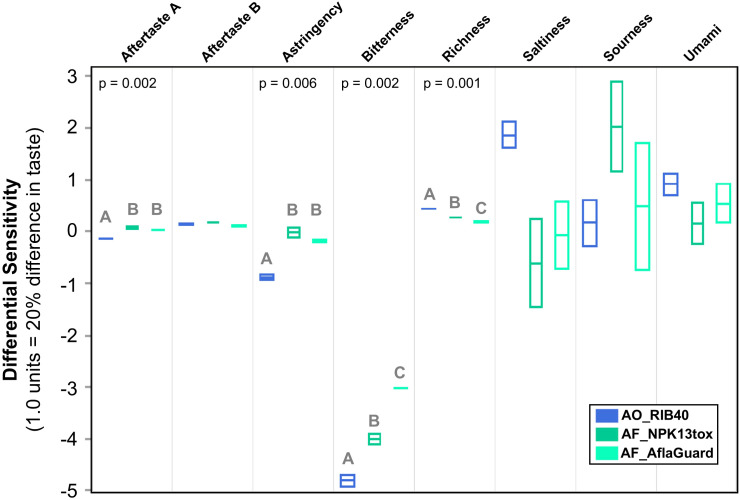 Figure 3
Figure 3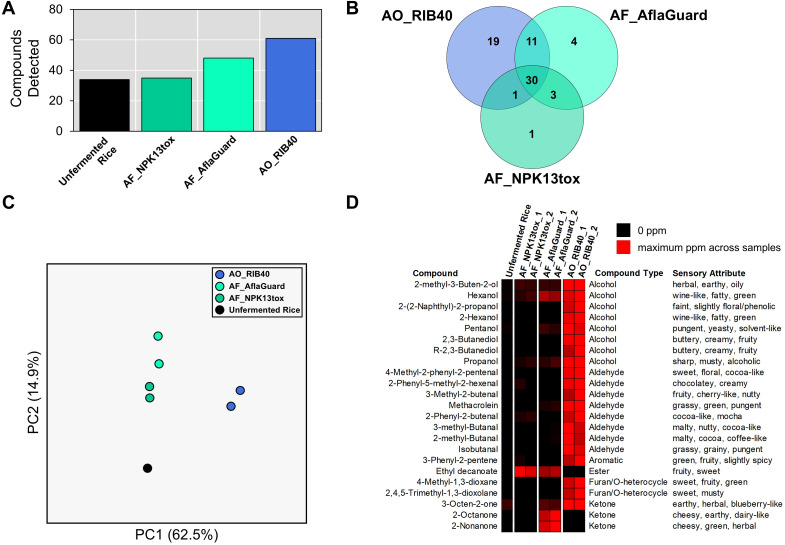 Figure 4
Figure 4| Compound type | Compound | Unfermented rice (negative control) | AF_NPK13tox_R1 | AF_NPK13tox_R2 | AF_AflaGuard_R1 | AF_AflaGuard_R2 | AO_RIB40_R1 | AO_RIB40_R2 |
|---|---|---|---|---|---|---|---|---|
| Acetal | Acetal | 0 | 0 | 0 | 0 | 0 | 0.42 | 1.18 |
| Acid | Acetic Acid | 0.06 | 0.54 | 0 | 0.15 | 0.53 | 0 | 0.62 |
| Isobutanoic acid | 0 | 0.11 | 0.47 | 0.15 | 0.35 | 0.59 | 0.44 | |
| Isopentanoic acid | 0 | 0 | 0 | 0 | 0 | 0.23 | 1 | |
| Alcohol | 1-Hepten-3-ol | 0 | 0.06 | 0 | 0.12 | 0.07 | 0.12 | 0.14 |
| 2-(2-Naphthyl)-2-propanol | 0 | 0 | 0 | 0 | 0 | 0.17 | 0.21 | |
| 2,3-Butanediol | 0 | 0 | 0 | 0 | 0 | 5.79 | 6.15 | |
| 2-Hexanol | 0 | 0 | 0 | 0 | 0 | 2.28 | 2.49 | |
| 2-methyl-3-Buten-2-ol | 0.08 | 0.33 | 0.29 | 0.31 | 0.29 | 1.32 | 1.3 | |
| 2-Nonen-1-ol | 0.04 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Butanol | 0.14 | 0 | 0 | 0.15 | 0 | 1.6 | 0.99 | |
| Ethanol | 109.38 | 772.87 | 854.88 | 948.22 | 1608.26 | 2798.91 | 4449.95 | |
| Heptanol | 0 | 0 | 0 | 0 | 0 | 13.53 | 22.52 | |
| Hexanol | 0.09 | 0.23 | 0.37 | 0.94 | 0.82 | 1.22 | 1.39 | |
| Isoamyl Alcohol | 0.23 | 8.6 | 6.53 | 12.21 | 9.24 | 138.53 | 199.96 | |
| Isobutanol | 0.76 | 10.75 | 7.26 | 12.25 | 15.59 | 217.53 | 307.81 | |
| Isopropyl Alcohol | 0 | 2.71 | 0 | 7.84 | 16.68 | 10.97 | 10.94 | |
| Pentanol | 0.18 | 0 | 0 | 0.72 | 0.51 | 2.93 | 2.57 | |
| Phenylethyl Alcohol | 0 | 1.44 | 2.14 | 0 | 0 | 0 | 0 | |
| Propanol | 0.43 | 4.52 | 8.12 | 7.87 | 12.76 | 41.44 | 42.9 | |
| R-2,3-Butanediol | 0 | 0 | 0 | 0 | 0 | 7.83 | 10.87 | |
| Aldehyde | 2-Butyl-2-Octenal | 0.07 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2-Heptenal | 0.17 | 0.37 | 0.14 | 0.31 | 0.19 | 0.47 | 0.24 | |
| 2-methyl-Butanal | 0.16 | 0.44 | 0.95 | 4.75 | 10.07 | 137.67 | 104.25 | |
| 2-Octenal | 0.07 | 0.94 | 0.13 | 0.1 | 0.1 | 0.21 | 0.26 | |
| 2-Phenyl-2-butenal | 0 | 1.73 | 2.4 | 0.48 | 0.28 | 10.5 | 13.69 | |
| 2-Phenyl-5-methyl-2-hexenal | 0 | 0.15 | 0 | 0 | 0 | 0.87 | 0.94 | |
| 3-Methyl-2-butenal | 0 | 0 | 0 | 0 | 0 | 0.12 | 0.17 | |
| 3-methyl-Butanal | 0.41 | 1.34 | 1.53 | 6.45 | 15.5 | 278.73 | 226.3 | |
| 4-Methyl-2-phenyl-2-pentenal | 0 | 0 | 0 | 0 | 0 | 0.88 | 0.95 | |
| 4-Methyl-3-pentenal | 0 | 0 | 0 | 0 | 0 | 1.45 | 2.15 | |
| Acetaldehyde | 0.44 | 0.55 | 0.9 | 3.54 | 3.11 | 8.92 | 12.02 | |
| Benzaldehyde | 0.15 | 0.4 | 1.52 | 0.57 | 0.43 | 2.13 | 1.71 | |
| Benzeneacetaldehyde | 0 | 2.82 | 8.79 | 1.9 | 2.26 | 20.96 | 14.63 | |
| Butanal | 0.06 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Decanal | 0.07 | 0 | 0 | 0.1 | 0.05 | 0.16 | 0.13 | |
| Heptanal | 0.26 | 0 | 0 | 0 | 0 | 0.28 | 0.19 | |
| Hexanal | 4.05 | 0.94 | 2.01 | 2.71 | 2.65 | 4.15 | 8.34 | |
| Isobutanal | 0.34 | 0.75 | 0.57 | 8.4 | 10.18 | 660.61 | 577.84 | |
| Methacrolein | 0.09 | 0.25 | 0.22 | 0.98 | 1.32 | 7.96 | 8.24 | |
| Methional | 0 | 0.09 | 0 | 0.06 | 0.12 | 2.71 | 1.32 | |
| Nonanal | 0.54 | 0.89 | 0.62 | 0.71 | 0.64 | 0.74 | 0.87 | |
| Octanal | 0.28 | 0.07 | 0.09 | 0.22 | 0.11 | 0.3 | 0.23 | |
| Pentanal | 0.46 | 0.33 | 0.6 | 3.26 | 6.02 | 0.81 | 0.6 | |
| Aromatic | 1,3-Di-tert-butylbenzene | 0 | 0 | 0 | 0 | 0 | 0.13 | 0 |
| 3-Phenyl-2-pentene | 0 | 0.1 | 0 | 0 | 0 | 0.65 | 0.9 | |
| Aromatic/Phenolic | 2-Methoxy-4-vinylphenol | 0 | 0.18 | 0.19 | 0.28 | 0.27 | 0.29 | 0.39 |
| Ester | Ethyl 2-methylbutyrate | 0 | 0 | 0 | 0 | 0 | 0.25 | 0.48 |
| Ethyl Acetate | 0.08 | 0.32 | 0.22 | 0.27 | 0.83 | 0.78 | 1.39 | |
| Ethyl decanoate | 0 | 0.4 | 0.34 | 0.25 | 0.28 | 0 | 0 | |
| Ethyl hexanoate | 0 | 0.05 | 0.06 | 0.34 | 0.51 | 0.34 | 0.26 | |
| Ethyl isobutyrate | 0 | 0 | 0 | 0.14 | 0.5 | 0.89 | 2.52 | |
| Ethyl nonanoate | 0 | 0.1 | 0.09 | 0.09 | 0.19 | 0.38 | 0.56 | |
| Ethyl octanoate | 0 | 0 | 0 | 0.05 | 0.12 | 0.14 | 0.15 | |
| Ethyl phenylacetate | 0 | 0 | 0 | 0 | 0 | 0.2 | 0.38 | |
| Ethyl-2-methyl-2-butenoate | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Hexyl isobutyrate | 0 | 0.26 | 0.15 | 0.12 | 0.12 | 0 | 0 | |
| Furan/O-heterocycle | 2,3-Dihydrofuran | 0 | 0.36 | 0.28 | 0.78 | 0.46 | 15.77 | 25.83 |
| 2,4,5-Trimethyl-1,3-dioxolane | 0 | 0 | 0 | 0 | 0 | 0.63 | 0.83 | |
| 2-pentyl-Furan | 0.19 | 0.11 | 0 | 0.16 | 0.09 | 0.28 | 0.28 | |
| 4-Methyl-1,3-dioxane | 0 | 0 | 0 | 0 | 0 | 1.01 | 1.24 | |
| Coumaran | 0.07 | 0.07 | 0.11 | 0.25 | 0.38 | 0.25 | 0.38 | |
| Furfural | 0 | 0 | 0 | 0 | 0 | 0.23 | 0 | |
| Pantolactone | 0 | 0.42 | 0.29 | 0.23 | 0.23 | 0.13 | 0.22 | |
| Ketone | 1-Hepten-3-one | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2,3-Pentanedione | 0 | 0 | 0 | 0.17 | 0.25 | 0.56 | 0.25 | |
| 2-Butanone | 0.07 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 2-Decanone | 0 | 0 | 0 | 2.52 | 3.84 | 0 | 0 | |
| 2-Heptanone | 0.09 | 0 | 0 | 4.28 | 8.45 | 0.16 | 0.17 | |
| 2-Nonanone | 0 | 0 | 0 | 4.11 | 4.93 | 0 | 0 | |
| 2-Octanone | 0 | 0 | 0 | 0.46 | 0.61 | 0 | 0 | |
| 3-methyl-2-Butanone | 0.21 | 2.62 | 2.11 | 2.16 | 1 | 6.63 | 4.3 | |
| 3-Methyl-5-heptanone | 0 | 0 | 0 | 0 | 0 | 0.71 | 0.35 | |
| 3-Octen-2-one | 0.05 | 0 | 0 | 0.07 | 0.06 | 0.2 | 0.2 | |
| Acetoin | 0 | 1.5 | 1.76 | 2.66 | 0.87 | 2.83 | 1.9 | |
| Acetone | 3.96 | 14.59 | 13.47 | 14.07 | 20.7 | 0 | 0 | |
| Terpene | beta-Elemene | 0 | 0.16 | 0.14 | 0 | 0 | 0.7 | 2.62 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Fermentation and Sensory Analysis · Microbial Metabolism and Applications
Introduction
Traditional fermented foods and beverages are a staple of many Asian diets. For instance, nearly 20% of the total weight and energy of Japanese dietary intake is from fermented foods (Fujihashi and Sasaki, 2024). Aspergillus oryzae is a filamentous mold and one of the most used microbes for fermented food production in Asia (e.g. miso, soy sauce, sake and amazake). In soy-based fermentations, A. oryzae is used primarily for its high proteolytic activity, while in rice-based fermentations A. oryzae is used for both proteolytic and amylolytic activity (Machida et al., 2005; Machida et al., 2008). A. oryzae was domesticated over thousands of years of use in food fermentation from its wild progenitor Aspergillus flavus (Gibbons et al., 2012; Watarai et al., 2019), which is a major agricultural pest and the main producer of aflatoxin contamination in stored seeds and grains (Amaike and Keller, 2011). As a result of domestication, A. oryzae lost its ability to produce some toxic secondary metabolites [including aflatoxin, cyclopiazonic acid, and aflatrem (Machida et al., 2005; Tominaga et al., 2006; Gibbons et al., 2012; Rank et al., 2012)] and adapted to the high starch environment of rice by increasing alpha-amylase production through gene duplications of the alpha-amylase encoding gene (Hunter et al., 2011; Chacon-Vargas et al., 2021). While the genetic differences between A. oryzae and A. flavus are well documented, far less is understood about how domestication shaped the sensory characteristics of A. oryzae.
During fermentation, A. oryzae produces a variety of extracellular enzymes to digest proteins, carbohydrates, and lipids into sugars, amino acids and free fatty acids that are subsequently reabsorbed and used as energy. These metabolites also significantly influence the volatile profile and sensory characteristics of the fermented food product. For instance, A. oryzae produces lipases during doenjang (soy paste) fermentation, and lipase activity shows strong associations with the formation of long−chain fatty acid esters that are essential for the final aroma and flavor (Kum et al., 2015). Additionally, in soy sauce fermentation, A. oryzae increases certain volatile compounds that impart “musty” and “soy-sauce-like” odors (Feng et al., 2013). Importantly, Zhao et al. (2015) showed substantial variation in volatile compound profiles (e.g. levels of ketones, aldehydes, alcohols and esters) between two A. oryzae strains during soy fermentation, which indicates that genetic differences within A. oryzae can directly result in distinct volatile profiles (Zhao et al., 2015).
Less is understood regarding the volatile, sensory, and enzymatic differences of foods fermented by these two strains, A. oryzae and A. flavus. Addressing these differences could provide insight into metabolic shifts impacted by A. oryzae’s long-term adaptation to food substrates. Rank et al. (2012) performed perhaps the only direct comparative metabolic study of the A. oryzae and A. flavus reference strains (RIB40 and NRRL 3357, respectively) using an LC-MS/MS, and found species-specific chemical profiles, including the absence of secondary metabolite end-products in A. oryzae (Rank et al., 2012). This result suggests A. oryzae and A. flavus have distinct metabolomic profiles, which are likely impacting their volatile profiles and sensory characteristics.
Here, we measured aflatoxin production, alpha-amylase activity, volatile profiles (via Dynamic Headspace Gas Chromatography Mass Spectrometry (DH-GC-MS)), and taste attributes (via E-tongue) of the fermentation starter culture A. oryzae RIB40 (AO_RIB40), the aflatoxin producer A. flavus NPK13tox (AF_NPK13tox), and the non-aflatoxin producer and biocontrol strain A. flavus AflaGuard (AF_AflaGuard). The goal of this study was to understand the differences in volatile and flavor characteristics between the domesticated food grade starter culture A. oryzae and its wild progenitor A. flavus.
Materials and methods
Aspergillus flavus 13tox genome sequencing and relationship of strains analyzed
We sequenced the genome of AF_NPK13tox to understand its relationship to AO_RIB40 and AF_AflaGuard. A. flavus 13tox was grown on potato dextrose agar (PDA) at 30 °C for 48 hours. DNA was extracted directly from the spores using the method outlined by (Zhao et al., 2019). DNA concentrations were measured for each extraction using a Qubit fluorometer. PCR-free 150-bp paired-end libraries were prepared and sequenced by Novogene (https://en.novogene.com/) on an Illumina NovaSeq 6000 platform. The raw whole-genome sequencing data for A. flavus 13tox can be accessed through the BioProject accession number PRJNA1277813. Raw paired-end fastq files were adapter and quality trimmed using trim_galore (Martin, 2011) with the following parameters: “-q 30”, “--stringency 1”, and “--length 80”. Adapter and quality trimmed reads were assembled using SPAdes (Bankevich et al., 2012) using the “--careful” option and K-mer sizes of 55, 75, 95, 105, and 115.
Eleven genome assemblies spanning the diversity of A. oryzae and A. flavus populations were obtained from NCBI and analyzed with the AF_NPK13tox assembly. These assemblies were A. flavus AflaGuard (GCA_012896875.1), OK-A-8-1-S (GCA_024676875.1), NRRL 3357 (GCF_009017415), A. oryzae TK14 (GCA_009685055.1), TK31 (GCA_009686725.1), TK49 (GCA_009687085.1), TK56 (GCA_009687225.1), TK57 (GCA_009687245.1), TK60 (GCA_009687305.1), RIB40 (GCF_000184455.2), 14160 (GCA_019097755.1) (Machida et al., 2005; Weaver et al., 2017; Watarai et al., 2019; Drott et al., 2020; Chacon-Vargas et al., 2021), and Aspergillus parasiticus CBS-117618 (GCA_009176385.1). For this analysis, gene models for all 11 genomes were predicted using Augustus v3.4.0 with the following parameters: “--species=aspergillus_oryzae,” “--strand=both,” “--gff3=on,” “--uniqueGeneId=true,” and “--protein=on” (Stanke and Waack, 2003). We used Augustus for gene prediction even for genomes with existing annotations to avoid bias from differences in gene model predictions. Single-copy orthologs across the 13 proteomes were identified using OrthoFinder v2.5.5 (Emms and Kelly, 2015; Emms and Kelly, 2019). The identified single copy ortholog proteins were concatenated and aligned using MAFFT v7.481 (Katoh and Standley, 2013). A phylogenetic tree was then constructed from the concatenated protein alignment with IQ-TREE v2.1.3 using 1,000 bootstrap replicates (-B 1000) and the LG amino acid replacement model (Minh et al., 2020).
Fungal isolates
AO_RIB40, AF_NPK13tox, and AF_AflaGuard were selected for rice fermentation and subsequent analysis because these strains represent distinct populations within the A. oryzae/A. flavus species complex (Figure 1). AO_RIB40 was obtained from ATCC, AF_AflaGuard was obtained from the USDA SRRC culture collection, and AF_NPK13tox was obtained from Nancy Keller’s lab at the University of Wisconsin Madison.
Rice fermentation experimental design and relationship of starter cultures. (A) Schematic of the experimental design, in which starter cultures fermented steamed rice after which the fermented products were subjected to volatile compound profiling via Dynamic Headspace Gas Chromatography Mass Spectrometry (DH-GC-MS), and flavor analysis via electronic tongue (e-tongue). (B) Maximum likelihood phylogenetic tree generated from a concatenated alignment of 8,092 single copy orthologs across 8 A. oryzae genomes (AO) and 4 A. flavus genomes (AF) and one A. parasiticus genome (AP), which was used to root the tree. Node labels represent bootstrap values. Genomes outlined in colored boxes note the strains used for rice fermentation in this study.
Culture conditions for alpha-amylase, e-tongue and DH-GC-MS analysis
We used Lundberg (Richvale, CA) Organic California Sushi Rice, which has a starch content ~75%, for all experiments involving rice fermentation. Freezer stocks of conidia for each strain were rapidly thawed and cultured on steamed rice 7 days at 30°C in petri dishes, at which time, the plates were flooded with 50% water + 49.9% glycerol + 0.1% tween to harvest conidia. Conidia were then quantified on a haemocytometer and normalized to 1x10^6^ conidia per mL. Rice was prepared for fermentation by autoclaving short grain white rice at 170% hydration for 15 minutes at 121°C. 12 g of sterilized short grain rice was inoculated with 1 mL of normalized conidia stocks (~1 million conidia) in 60 mm petri dishes sealed with parafilm and incubated for 48 hours at 30°C. Fermentations for amylase-activity and e-tongue were performed in triplicate, while fermentations performed for DH-GC-MS were performed in duplicate. However, we only analyzed a single unfermented rice (negative control) sample for e-tongue and DH-GC-MS analyses. Thus, control values serve as a baseline reference rather than a replicated measure of variability. As a negative control we also performed the same experiment but added 1 mL of the conidia harvesting solution (50% water + 49.9% glycerol + 0.1% tween) without conidia (rice only).
At the end of the 48 hours period, samples for e-tongue and DH-GC-MS were transferred to 50 mL conical tubes, 25 mL of water was added, and samples were vortexed for 60 s. A 48-hour incubation was selected because it reflects the typical duration used in standard koji production, during which A. oryzae reaches peak amylolytic activity and produce characteristic early-stage flavor compounds. Water was incorporated at the end of fermentation solely to enable e-tongue measurements, which require liquid-phase samples. Samples were flash-frozen in liquid nitrogen immediately after collection and shipped on dry ice via overnight express to Medallion Labs (Minneapolis, MN). Transit time was less than 12 hours, and samples were stored in a freezer immediately upon receipt. An unfractionated aliquot of the fermented rice/water slurry was used for E-tongue and DH-GC-MS analysis, minimizing the potential for volatile loss. Our overall experimental design is depicted in Figure 1.
Alpha-amylase activity
We measured alpha amylase activity during rice fermentation across the three fermentation starters, because alpha amylase activity is essential for rice fermentation and is one of the defining features of A. oryzae domestication (Hunter et al., 2011; Gibbons et al., 2012). To measure alpha amylase activity, 12.25 g of fermented rice samples were combined with 10 mL of molecular water, vortexed for 1 minute, and a 2 mL aliquot was withdrawn with analysis. The 2 mL aliquot was fractionated by centrifugation at 1,000 g for 10 min. A 1 mL aliquot of the resulting supernatant was withdrawn and used for alpha amylase quantification. Alpha amylase activity of each strain was evaluated with Ceralpha α-Amylase Assay Kit (cat. No. K-CERA) in biological triplicate. This method utilizes non-reducing end-blocked p-nitrophenyl maltoheptaoside in the presence of an excess of thermostable α-glucosidase. The supernatant was diluted 1:24 with the assay buffer before proceeding with the assay protocol. Absorbance values (405 nm) were recorded with a SpectraMax i3 microplate reader (Molecular Devices, USA). The negative control for this assay was developed by combining all reagents in the same proportions as test samples, but without incubation such that the active enzyme was denatured and thus no longer capable of hydrolyzing starch. This was to confirm that any changes in absorbance (405 nm) were due to enzymatic activity rather than non-enzymatic activity or baseline fluctuations.
Aflatoxin quantification
Aflatoxin quantification was carried out using a modified protocol adapted from Alshannaq et al. (2018) and Acevedo et al. (2025) (Alshannaq et al., 2018; Acevedo et al., 2025). Isolates were grown in slant cultures prepared with 2 mL of potato dextrose (PD) medium in 10 mL glass test tubes. Approximately 10^5^ conidia were inoculated into each tube using a sterile loop. The tubes were placed at a 45° angle on a rack and incubated at 30 °C for 7 days to allow for mycelial growth and toxin production. After incubation, aflatoxins were extracted using an organic solvent method. One mL of chloroform was added directly to each culture tube, thoroughly mixed, and then centrifuged at 5,000 × g to facilitate phase separation. From the chloroform phase, 0.5 mL was carefully collected and left to evaporate at room temperature. The resulting residue was reconstituted in 0.5 mL of High-performance liquid chromatography (HPLC) mobile phase consisting of water, methanol, and acetonitrile (50:40:10, v/v/v). Samples were filtered through a 0.45 μm membrane filter prior to HPLC analysis.
HPLC was used to quantify aflatoxins AFB1, AFB2, AFG1, and AFG2. Analyses were performed on an Agilent 1100 series system equipped with a degasser, autosampler, quaternary pump, and a 1260 Infinity diode array detector (Agilent Technologies, CA, USA). Separation was achieved using a Zorbax Eclipse XDB-C18 column (4.6 mm × 150 mm, 3.5 μm particle size) with a flow rate of 0.8 mL/min. Aflatoxins were detected at a wavelength of 365 nm.
E-tongue
Traditional descriptive sensory testing with human subjects was not possible because AF_NPK13tox is an aflatoxin producer. Thus, we analyzed our samples via e-tongue, which measures flavor attributes by using an array of chemical sensors combined with pattern recognition algorithms to detect and differentiate flavor-related compounds in a liquid sample. E-tongue analysis assessed these compounds and provided an interpretation of how these might impact eight different flavor profiles (i.e., umami, aftertaste richness of umami, saltiness, sourness, bitterness, aftertaste of bitterness, astringency, and aftertaste of astringency). These attributes are based on a relative comparison to internally developed references. Data were reported as relative comparisons to the control, using a scale where 1 unit represents a level of differential sensitivity equivalent to an estimated 20% difference in taste. Three biological replicates were analyzed for each fermentation, and one sample was analyzed for the unfermented rice negative control to establish a baseline. For each flavor attribute, we conducted an ANOVA between the fermentation starter strains assuming a p-value < 0.05 cutoff. For comparisons in which p-values < 0.05, we conducted post hoc Tukey Kramer HSD tests to compare pairwise differences between samples, imposing a p-value < 0.05 cutoff. Statistical analysis was conducted in JMP v18.2.0.
DH-GC-MS
All sample processing, compound identification and quantification were performed at Medallion Labs (Minneapolis, MN). Samples were analyzed as received by dynamic headspace on an Agilent 5977 MSD coupled to an Agilent 7890A gas chromatography unit with a Gerstel multipurpose sampler (Agilent, USA). Compounds were identified and matched by a proprietary spectrum library, with no other standards or calibrations. Quantitation of compounds present was determined by comparison of their response to that of an internal standard added to each sample. Prior to testing, Parts Per Million (PPM) values were calculated relative to the total sample weight as received (wet basis) (as samples consisted of frozen fermented rice/water slurry). Because each fermentation condition was represented by only two biological replicates (and the unfermented control by one), we did not perform inferential statistical tests on the DH-GC-MS volatile compound data. Instead, for each detected compound we summarized relative abundance across replicates using descriptive statistics (mean and range) to characterize strain-dependent patterns.
Results
Fermentation strains are from phylogenetically distinct groups
We analyzed the phylogenetic relationship of the three rice fermentation starters (AO_RIB40, AF_AflaGuard and AF_NPK13tox) in relation to 7 additional A. oryzae genomes, 2 additional A. flavus genomes that are representative of the major populations based on previously published work (Gibbons et al., 2012; Watarai et al., 2019; Drott et al., 2020; Chacon-Vargas et al., 2021; Kim et al., 2025), and a strain of A. parasiticus which was used as an outgroup. The genome of AF_NPK13tox was sequenced and assembled for the current study (BioProject ID PRJNA1277813), while all other genome assemblies were publicly available (see methods for accession numbers). We used OrthoFinder to identify single copy orthologs (Emms and Kelly, 2015; Emms and Kelly, 2019), and then built a maximum likelihood phylogenetic tree based on a protein alignment of 8,092 single copy orthologs (which included 4,445,407 amino-acid sites and 48,990 parsimony informative sites). The phylogenetic tree showed three phylogenetically distinct clades of A. oryzae, and two clades of A. flavus (Figure 1), consistent with previous studies. Importantly, the three fermentation strains resided in distinct clades.
Alpha-amylase activity is significantly higher in A. oryzae RIB40
We measured alpha-amylase activity at the end of our fermentation because variability in alpha-amylase production could impact volatile compound formation (e.g. higher sugar concentrations could lead to more ethanol production by yeasts which in turn could increase the pool of substrates for ester synthesis), and taste perception (e.g. higher alpha-amylase activity would increase sugar content and sweetness of the fermentation product) (Yoshizawa, 1999). Unsurprisingly, alpha-amylase activity was significantly different between isolates (Figure 2) (ANOVA, p-value = 5.2e-6) and significantly higher in AO_RIB40 compared to the A. flavus isolates (Tukey-Kramer: AO_RIB40 vs. AF_NPK13tox: p-value = 1.0e-4 and AO_RIB40 vs. AF_AflaGuard: p-value = 8.6e-6).
Alpha-amylase activity during rice fermentation. Alpha-amylase activity was quantified in the rice fermentation with the Ceralpha α-Amylase Assay Kit (cat. No. K-CERA) in biological triplicate. Box plots for each strain (x-axis) represent alpha-amylase activity (y-axis). The ANOVA p-value is reported, and letters above box plots signify statistically different groups based on post hoc Tukey Kramer HSD tests. AO_RIB40 has higher alpha-amylase activity compared to both A. flavus strains during rice fermentation.
Only A. flavus NPK13tox produces aflatoxins
We quantified the production of aflatoxins (AFB_1_, AFB_2_, AFG_1_, and AFG_2_) by AO_RIB40, AF_NPK13tox, and AF_AflaGuard under culture conditions known to promote aflatoxin biosynthesis (Alshannaq et al., 2018; Acevedo et al., 2025). Aflatoxins were not detected in cultures of AO_RIB40 or AF_AflaGuard. In contrast, AF_NPK13tox produced approximately 97 ppm AFB_1_ and 34 ppm AFG_1_. These results are consistent with prior knowledge: AF_NPK13tox is closely related to the aflatoxigenic strain A. flavus NRRL 3357 (Figure 1), whereas AO_RIB40 and AF_AflaGuard harbor well-documented loss-of-function mutations in the aflatoxin biosynthetic gene cluster (Gibbons et al., 2012; Alshannaq et al., 2018).
E-tongue analysis for A. oryzae RIB40 fermentation has more favorable profile
Fermented rice water slurries underwent e-tongue analysis to measure and compare flavor attributes between starter cultures. Compounds known to associate with perceived saltiness, sourness, umami, umami aftertaste, astringency, aftertaste-A (astringency aftertaste), bitterness and aftertaste-B (bitter aftertaste) were measured for the unfermented rice negative control, and the biological replicates of each fermentation starter culture. Four of the 8 comparisons were statistically significant (ANOVA, all p-values ≤ 0.002, Figure 3). Astringency is a tactile sensation perceived as dryness, roughness, and a tightening feeling in the mouth, often accompanied by a slight contraction of the tongue and oral tissues, and it is often accompanied with acidity or bitterness sensations (Pires et al., 2020). Interestingly, astringency and aftertaste-A was significantly lower in the AO_RIB40 fermentation than in the A. flavus fermentations (Figure 3). Additionally, bitterness was substantially lower in all samples relative to the rice control and significantly lower in the AO_RIB40 fermentation compared to the A. flavus fermentations which were themselves significantly different (Figure 3). Lastly, umami aftertaste, described as the richness, complexity, and full bodied, was significantly higher in the AO_RIB40 fermentation compared to the A. flavus fermentations which, were also significantly different from each other (Figure 3).
E-tongue analysis of rice fermentation using different starter cultures. Box plots of 8 taste attributes measured by e-tongue (y-axis) across rice fermented by AO_RIB40, AF_NPK13tox, and AF_AflaGuard (x-axis). The unfermented rice negative control is represented as a normalized score of 0 for all attributes; all sample values are plotted relative to this baseline, with positive values indicating an increase and negative values a decrease in the respective attribute compared to the control. Box plots represent measurements from three biological replicates per starter culture. The ANOVA p-value is reported for taste attributes that were statistically significant via ANOVA across the three starter cultures, and letters above box plots signify statistically different groups based on post hoc Tukey Kramer HSD tests. The Y-axis represents differential sensitivity in which 1 unit represents a 20% difference in taste.
Volatilome profiles vary significantly between AO and AF samples
We performed DH-GC-MS of the fermented rice water slurries to identify unique compounds and compare the relative abundances of compounds between starter cultures. We detected 34, 35, 48, and 61 unique compounds in the rice slurry without a starter culture, the AF_NPK13tox fermentation, the AF_AflaGuard fermentation, and the AO_RIB40 fermentation, respectively (Figure 4, Table 1). Between the starter culture samples, 30 compounds were detected in all samples, while 1, 4 and 19 compounds were uniquely detected in the AF_NPK13tox, AF_AflaGuard, and the AO_RIB40 fermentations, respectively (Figure 4).
Volatile compound profiling of rice fermentation using different starter cultures. Volatile compound profiles were identified relatively quantified for rice fermentations across biological replicates of AO_RIB40, AF_NPK13tox, and AF_AflaGuard and unfermented rice (neg. control). (A) Number of unique volatile compounds detected the unfermented and fermented rice samples. (B) Venn diagram of volatile compounds between AO_RIB40, AF_NPK13tox, and AF_AflaGuard rice fermentations. (C) Principal Components Analysis of volatile compound profiles between samples. PC1 explains 62.5% of variance, while PC2 explains 14.9% of variance. (D) Heatmap depicting relative differences in volatile compound abundance across AO_RIB40, AF_NPK13tox, and AF_AflaGuard rice fermentations. Values are scaled within each compound (row) to highlight patterns of higher and lower abundance among starter cultures, where black represents absence and red represents the maximum PPM value observed for that compound.
To understand how samples volatile profiles were related, we performed a PCA on each sample and replicate using the compound abundance (in parts per million) matrix as input. PC1 explained 62.5% of variance, and separated rice, AF_NPK13tox, and AF_AflaGuard, by AO_RIB40, while PC2 explained 14.9% of variance and separated the AF strains and the rice sample (Figure 4).
To compare compound abundance patterns between fermentations, we summarized the two biological replicates for each starter culture using descriptive statistics, rather than inferential tests, because replication was limited (n = 2 per fermentation; n = 1 control). Several clear trends emerged. Across the panel of detected compounds, AO_RIB40 consistently produced substantially higher levels of many alcohols, aldehydes, ethers, esters, and ketones compared with the A. flavus starter cultures (Table 1). For example, some compounds of note include 2,3-butanediol and R-2,3-butanediol, which was uniquely detected in AO_RIB40 and contributes to the floral and fruity aroma of soy sauce, 3-methyl-Butanal, which is an aldehyde detected in moromi fermentation (Li et al., 2023), and 2-methyl-butanal, which has a cocoa or coffee-like aroma and has been detected in fermented sausages (Olivares et al., 2009) (Table 1, Figure 4). Conversely, a small number of compounds were detected in both A. flavus fermentations but not in AO_RIB40. Interestingly, one such compound, ethyl decanoate, has been linked to both sweet and vinegar aromas (Xiao et al., 2016), and was not detected in the AO_RIB40 fermentations.
Discussion
Food fermentation has a profound impact on the taste and flavor profile of foods. However, little work has compared the impact of A. oryzae and A. flavus on these compounds during food fermentation. To address this gap, we conducted rice fermentations using three distinct starter cultures: the food-grade strain AO_RIB40 and two closely related wild strains, AF_NPK13tox and AF_AflaGuard. We performed volatile compound profiling using DH-GC-MS and evaluated sensory attributes using an e-tongue (Figure 1). In addition, we quantified aflatoxin production to assess food safety and measured alpha-amylase activity to evaluate fermentation efficiency across the starter cultures.
The higher alpha-amylase output of AO_RIB40 (Figure 2) is a direct consequence of domestication, where artificial selection favored more efficient starch metabolism (Hunter et al., 2011; Gibbons et al., 2012; Watarai et al., 2019). Consistent with a previous proteomic analysis of rice fermentation, AO_RIB40 exhibited significantly higher alpha-amylase activity than the A. flavus strains (Gibbons et al., 2012). This parallels genomic observations that all A. oryzae isolates carry multiple copies of the α-amylase gene, whereas wild A. flavus typically has only one copy (Gibbons et al., 2012; Watarai et al., 2019). The A. flavus strains in our study (with one alpha-amylase gene copy) showed significantly weaker amylase activity. The enhanced starch breakdown by AO_RIB40 likely accelerates saccharification and yeast fermentation, increasing ethanol yield and fermentation speed. Interestingly, variation in alpha-amylase activity is present even within A. oryzae strains. For instance, Chacón‐Vargas et al., 2020 found that AO_RIB40 produced significantly more amylase (and grew faster on starch) than A. oryzae 14160 during solid rice fermentation (Chacon-Vargas et al., 2021), and suggested the importance of strain selection depending on fermentation substrate.
Additionally, we quantified aflatoxin production across our three isolates and confirmed that only AF_NPK13tox produced aflatoxin. AO_RIB40 has at least 4 loss-of-function mutations in the aflatoxin encoding gene cluster (Tominaga et al., 2006) and AF_AflaGuard has a large deletion encompassing the entire aflatoxin encoding gene cluster (Alshannaq et al., 2018; Chang et al., 2021). However, despite this lack of aflatoxin production in AF_AflaGuard, this strain lacks the alpha-amylase activity and favorable sensory profile of AO_RIB40 (Figures 3, 4).
The E-tongue results reveal clear differences in compounds associated with flavor attributes. Based on these results, it is anticipated that rice fermentations of A. oryzae RIB40 would be perceived has producing significantly less bitterness, astringency, and aftertaste A compared to those made with A. flavus strains (Figure 3). This observation likely reflects the different metabolite profiles between starter cultures and may be explained by a shift from primary metabolism in A. oryzae (e.g., high alpha-amylase activity) to secondary metabolism in wild A. flavus isolates. Secondary metabolites could lead to bitter compound production, and, generally, less proteolytic activity by shifting energy investment away from primary metabolism (Gibbons, 2019). Due to the safety concerns with consumption, these E-tongue results provide the first insights into the potential flavor differences produced by these two strains.
Our study demonstrates that rice fermented with A. oryzae produces a significantly richer volatile profile in compounds associated with favorable sensory attributes compared to fermentations performed with A. flavus strains (Hu et al., 2020). Specifically, A. oryzae fermentations yielded higher levels of unsaturated alcohols, ketones, and aldehydes that contribute complex sensory attributes (Kum et al., 2015). Among the elevated compounds were 2-methyl-3-buten-2-ol and 3-octen-2-one. 2-methyl-3-buten-2-ol may impart distinctive fruity or green nuances that heighten the freshness of the aroma, while 3-octen-2-one is generally associated with mushroom or earthy notes, which may suggest a combination of earthy and umami attributes in the fermented rice (Lee et al., 2019).
Furthermore, both 2-methyl-butanal and 3-methyl-butanal (aldehydes associated with malty aromatic notes) were produced at significantly higher levels in the AO_RIB40 fermentations (Figure 4). These compounds were also detected in rice koji fermented by A. oryzae (Hwang and Kim, 2023). These branched-chain aldehydes are key contributors to the characteristic rich malty aroma of soy sauce (Lee et al., 2006; Ly et al., 2023).
Several heterocyclic compounds (4-methyl-1,3-dioxane and 2,4,5-trimethyl-1,3-dioxolane), were also more abundant in the A. oryzae fermentation. These compounds, often formed through Maillard-type reactions during fermentation, can contribute sweet, caramel-like, or even complex fruity notes that enhance the overall flavor profile (Hu et al., 2020). Their presence may also indicate A. oryzae’s more efficient enzymatic transformations and sugar degradation processes (Kum et al., 2015). We also detected several alcohols at significantly greater abundance in AO_RIB40 compared to the A. flavus fermentations (i.e. 2-methyl-3-Buten-2-ol, hexanol, 2-(2-Naphthyl)-2-propanol, 2-hexanol, pentanol, 2,3-butanediol, R-2,3-butanediol, and propanol). These observations are consistent with rice wine fermentation, where alcohols are more abundant early in the fermentation process and are gradually converted into esters and other sensory compounds during as fermentation continues (Liu et al., 2020; Yu et al., 2023).
Additionally, phenyl-substituted aldehydes such as 4-methyl-2-phenyl-2-pentenal, 2-phenyl-5-methyl-2-hexenal, 3-phenyl-2-pentene, and 2-phenyl-2-butenal, as well as the aromatic alcohol 2-(2-naphthyl)-2-propanol, were present in higher concentrations. These compounds often contribute floral, balsamic, and nutty nuances that have been linked to elevated sensory appeal in rice-based fermentations, consistent with the role of aromatic ring structures in enhancing sweet and floral odor profiles (Hu et al., 2020). The appearance of these compounds suggests that A. oryzae fermentations favor the release or synthesis of aromatic precursors derived from amino acid metabolism and lipid oxidation (Yang et al., 2013). Further, higher abundances of simpler alcohols (2-hexanol, pentanol, and propanol), as well as branched aldehydes (3-methyl-butanal, 2-methyl-butanal, and isobutanal), were detected at higher abundance in the AO_RIB40 fermentations. These compounds are known to contribute green, fresh, malty, and even slightly fatty aromas, all of which are desirable in the context of rice fermentations for achieving a well-rounded flavor profile (Hu et al., 2020).
Our results are similar to Bodinaku et al. (2019), in which Penicillium strains were experimentally evolved in conditions mimicking the cheese making environment and volatile compounds were compared against the ancestral strains (Bodinaku et al., 2019). The wild ancestors characteristically produced musty, earthy odorants such as geosmin, whereas lineages serially cultured in the cheese environment largely lost the capacity to synthesize this compound. Instead, cheese lineages shifted their metabolome and produced volatile compounds contributing descriptors like “cheesy” and “fatty,” which are desirable attributes in cheese making. This study, and our previous work (Gibbons et al., 2012), identified a down-regulation of genes involved in the production of secondary metabolites in strains adapted to the food environment compared to their wild counterparts, which likely indicates a shift away from the production of costly defense compounds not needed in the nutrient-rich, low-competition fermentation environments, where the energetic cost of producing a broad array of secondary metabolites is disadvantageous [ (Gibbons, 2019; Chen et al., 2022)].
From an industrial perspective, the strain-specific sensory and volatile signatures observed here illustrate how starter culture selection can be used strategically to tune flavor development in rice-based fermented products (Knez et al., 2023; Li et al., 2023). Because these sensory outcomes arise from distinct metabolic activities, our findings also suggest possible nutritional implications (Kaya et al., 2025; Villasante et al., 2025), such as differences in enzymatic breakdown and formation of bioactive compounds. Together, these insights provide a framework for integrating sensory-chemistry profiling into strain selection and quality-control strategies in commercial fermentation.
Conclusions
This study highlights the biochemical and sensory distinctions between the domesticated strain AO_RIB40 and its wild relatives, AF_NPK13tox and AF_AflaGuard, during rice fermentation. The enhanced α-amylase activity and absence of aflatoxin production in A. oryzae underscore its safety and effectiveness as a fermentation starter. Notably, rice fermented with A. oryzae exhibited a more favorable sensory profile, characterized by reduced bitterness and astringency and an increased umami-rich aftertaste. Volatile compound analysis revealed that A. oryzae produced higher levels of alcohols, aldehydes, ketones, and heterocyclic compounds associated with fruity, malty, and umami aromas. Collectively, these findings suggest that long-term domestication has shaped the volatilome of A. oryzae to enhance desirable sensory qualities in fermented foods.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Acevedo K. L. Eaton E. Leite J. Zhao S. Chacon-Vargas K. Mccarthy C. M. . (2025). Population genomics of aspergillus sojae is shaped by the food environment. Genome Biol. Evol. 17. doi: 10.1093/gbe/evaf 067, PMID: 40195023 PMC 12014904 · doi ↗ · pubmed ↗
- 2Alshannaq A. F. Gibbons J. G. Lee M. K. Han K. H. Hong S. B. Yu J. H. . (2018). Controlling aflatoxin contamination and propagation of Aspergillus flavus by a soy-fermenting Aspergillus oryzae strain. Sci. Rep. 8, 16871. doi: 10.1038/s 41598-018-35246-1, PMID: 30442975 PMC 6237848 · doi ↗ · pubmed ↗
- 3Amaike S. Keller N. P. (2011). Aspergillus flavus. Annu. Rev. Phytopathol. 49, 107–133. doi: 10.1146/annurev-phyto-072910-095221, PMID: 21513456 · doi ↗ · pubmed ↗
- 4Bankevich A. Nurk S. Antipov D. Gurevich A. A. Dvorkin M. Kulikov A. S. . (2012). SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021, PMID: 22506599 PMC 3342519 · doi ↗ · pubmed ↗
- 5Bodinaku I. Shaffer J. Connors A. B. Steenwyk J. L. Biango-Daniels M. N . (2019). Rapid phenotypic and metabolomic domestication of wild penicillium molds on cheese. M Bio 10. doi: 10.1128/m Bio.02445-19, PMID: 31615965 PMC 6794487 · doi ↗ · pubmed ↗
- 6Chacon-Vargas K. Mc Carthy C. O. Choi D. Wang L. Yu Gibbons J. H. J. G . (2021). Comparison of two aspergillus oryzae genomes from different clades reveals independent evolution of alpha-amylase duplication, variation in secondary metabolism genes, and differences in primary metabolism. Front. Microbiol. 12, 691296. doi: 10.3389/fmicb.2021.691296, PMID: 34326825 PMC 8313989 · doi ↗ · pubmed ↗
- 7Chang P. K. Chang T. D. Katoh K. (2021). Deciphering the origin of NRRL 21882, the active biocontrol agent of Afla-Guard(R). Lett. Appl. Microbiol. 72, 509–516. doi: 10.1111/lam.13433, PMID: 33251654 · doi ↗ · pubmed ↗
- 8Chen W. Lv X. Tran V. T. Maruyama J. I. Han K. H. Yu J. H. . (2022). Editorial: from traditional to modern: progress of molds and yeasts in fermented-food production. Front. Microbiol. 13, 876872. doi: 10.3389/fmicb.2022.876872, PMID: 35401444 PMC 8992652 · doi ↗ · pubmed ↗