Outer membrane vesicles as a versatile nanoplatform for advanced vaccine development and immunotherapy
Jiawen Zhang, Yuhang Zhi, Yu Lu, Fang Ma

TL;DR
This paper reviews bacterial outer membrane vesicles as a promising nanoplatform for vaccine development and immunotherapy.
Contribution
The paper provides a comprehensive review of OMV biogenesis, composition, and therapeutic potential, aiming to inspire new research and clinical applications.
Findings
OMVs are nano-sized vesicles with diverse biomolecules that play roles in bacterial physiology and communication.
OMVs have promising therapeutic potential for vaccine development and immunotherapy.
The paper integrates current knowledge to guide future translational research on OMVs.
Abstract
Bacterial outer membrane vesicles (OMVs) are nano-sized, lipid-bilayer vesicles naturally released by Gram-negative bacteria. These vesicles are enriched with diverse biomolecules, including lipids, proteins, and nucleic acids, enabling them to mediate critical biological processes. Emerging evidence highlights their pivotal roles in bacterial physiology, intercellular communication, and environmental adaptation, alongside their promising therapeutic potential. This review focuses on recent advances in OMV biogenesis, composition, function and applications. By integrating current knowledge, we aim to inspire novel insights into the molecular mechanisms underlying OMV functions and facilitate their translational development in medicine. Ultimately, this work serves as a comprehensive reference to accelerate future research and clinical utilization of this versatile platform.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
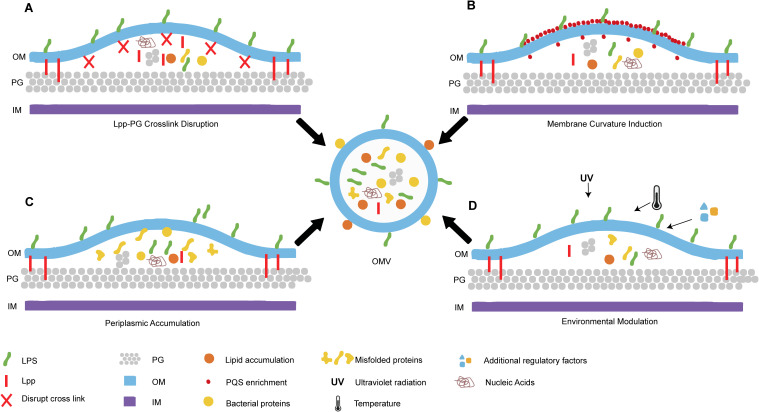 Figure 1
Figure 1| Related genes | Reported bacteria | Functions | References |
|---|---|---|---|
| 1. | 1.OprF is a hydrophilic channel-forming protein contributing to OM permeability; | ( | |
|
| Gram-negative bacteria like |
1. Gram-negative bacteria outer membrane β-barrels protein family; 2. An increased vesiculation due to deletion or truncation of | ( |
|
|
| 1. | ( |
|
|
| 1. YpjA is a hypothetical Omp; | ( |
|
|
| 1. BolA is thought to regulate morphology changes as well as increased resistance to antibiotics and detergents in stationary phase and under conditions of stress; | ( |
|
|
| 1. DsbA is a disulfide oxidoreductase which aids in periplasmic protein folding by inducing disulfide bonds; | ( |
| Tol-pal genes | 1. Tol-Pal system is a multiprotein complex of five envelope proteins, TolQ, TolR, TolA, TolB, and Pal; | ( | |
|
|
| 1. | ( |
|
| Gram-negative bacteria like | 1. VacJ/Yrb ABC (ATP-binding cassette) transport system is a proposed phospholipid transporter; | ( |
|
|
| 1. MreB is a rod shape-determining protein; | ( |
|
|
| 1. Thiol-disulfide interchange protein; | ( |
|
|
| 1. Phosphoheptose isomerase; | ( |
|
|
| 1. Transmembrane anti-sigma factor; | ( |
|
| 1. Hemolysin F (HlyF) lacks hemolytic activity acting as a cytoplasmic enzyme, and capable of delivering a bona fide hemolysin ClyA; | ( | |
|
|
| 1. CprA is an a short-chain dehydrogenase/reductase (SDR) contributing to resistance against colistin and antimicrobial peptides; | ( |
|
|
| 1. Salmonella outer membrane protein PagC is a major constituent of Salmonella membrane vesicles; | ( |
|
| Gram-negative bacteria | 1. OmpX belongs to a family of onserved proteins, including PagC from S. enterica serovar Typhimurium; | ( |
|
|
| 1. The | ( |
|
| 1. | ( | |
|
|
| 1. | ( |
|
|
| 1. NlpI is an outer-membrane lipoprotein associated with cell division; | ( |
| Anchor proteins | Function/potential application | Reference |
|---|---|---|
| clyA | ClyA is a pore-forming cytotoxin of enterobacteria, acting as a membrane anchor, ensuring the efficient display of antigens on the OMV surface. | ( |
| OmpA | 1. Outer membrane protein A (aminoacids 1-131), which can be fused by heterologous proteins on OMV surface; | ( |
| Lpp-OmpA | 1. Lpp-OmpA is a membrane anchor protein, which can be effectively exploited for the presentation of desired proteins on the OMV surface; | ( |
| Hbp | 1. | ( |
| OmpF | 1. Porins involved in diffusion of ions and molecules; | ( |
| MipA | 1. | ( |
| LamB | 1. LamB is a trimeric OMP that functions in the transport of maltose and maltose polymers and also serves as the receptor for bacteriophage λ; | ( |
| OmpC | 1. OmpC and OmpA were previously identified as OMV proteins, and they are often used as marker proteins for OMVs; | ( |
| OmpW | OmpW is constantly and abundantly expressed on the outer membranes of | ( |
| FhuA | 1. FhuA is a natural iron transporter located in the outer membrane of | ( |
| PelB peptide signal | 1. Pectate lyase subunit (PelB); | ( |
| OMV-based vaccine types | Features | Representative pathogens/disease models | The merits | The drawbacks | Current development stage | References |
|---|---|---|---|---|---|---|
| Native OMVs | Naturally released vesicles purified with detergents to reduce LPS content |
| High immunogenicity; Relatively simple production | Detergent use can lead to loss of key antigens; Heterogeneous composition; Potential for autoimmunity | Licenced (VA-MENGOC-BC, MenBvac); Clinical trials | ( |
| Genetically attenuated OMVs | Gene knockout/modification to produce detoxified lipid A; deletion of virulence factors | Improved safety profile; Retains self-adjuvanticity; Genetically defined process | May slightly reduce immunogenicity; Requires genetic engineering expertise | Clinical trials; Preclinical | ( | |
| Antigen-display OMVs | Genetic fusion of heterologous antigens to anchor proteins | SARS-CoV-2; Tumor therapy (displayed tumor antigens) | Targeted immune responses; Multivalent antigen presentation; No need for antigen purification | Correct folding and display can be challenging; Genetic fusion may affect OMV biogenesis | Clinical trials phase I (Avacc 10); Preclinical | |
| Cargo-loaded OMVs | Loading of therapeutic molecules (mRNA, drugs, proteins) via electroporation, sonication, or co-incubation | Directing enzyme packing delivery; Tumor therapy delivery | Protects cargo from degradation; Enables combination therapy; Versatile payload capacity | Low loading efficiency; Concerns about stability and toxicity; Potential damage to OMV structure; Uncontrolled release | Preclinical; | ( |
| Hybrid/Biomimetic OMVs | Fusion of OMVs with synthetic liposomes or coating onto synthetic nanoparticle cores | Tumor immunotherapy; Drug delivery for chronic bacterial infections | Tunable properties; Enhanced stability; Synergistic functionality | Complex manufacturing process may affect the membrane topology and biofunctions; Batch-to-batch variability | Preclinical | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Pneumonia and Respiratory Infections · Bacteriophages and microbial interactions
Introduction
1
The initial discovery of bacterial outer membrane vesicle (OMVs) dates to the 1960s, when electron microscopy first revealed their presence in Gram-negative bacteria (1). Initially dismissed as cellular debris due to limited understanding of their biological roles, OMVs gained scientific attention following their detection in the plasma of a patient with fatal meningococcal septicemia (2). As research advanced, these vesicular structures were identified in an expanding range of bacterial species, including Gram-positive organisms (3).
OMVs are nano-sized with diameters ranging from 20-300 nm, spherical structures naturally released by Gram-negative bacteria through outer membrane (OM) budding (4). Their characteristic components, including lipopolysaccharide (LPS), outer membrane proteins (OMPs), and associated peptidoglycan (PG), vary significantly across bacterial species and growth conditions, directly correlating with functional diversity (5, 6). For bacteria, OMVs multifunctional tools central to survival and adaptation. They serve as communication vehicles, facilitating interbacterial signaling, horizontal gene transfer (HGT), and virulence factor dissemination (7–9). Simultaneously, they act as a defense system by neutralizing antimicrobial compounds and as an offensive weapon by delivering cytotoxic effectors to host cells (10, 11). Furthermore, OMVs are integral to biofilm formation, providing structural support and enhancing community resilience (12). In host interactions, OMVs directly modulate immune responses, capable of both activating and suppressing specific pathways (13). This complex interplay between microbial activity and host immunomodulation underscores their significance in infection. The convergence of these biological properties has propelled OMVs into biomedical applications. Their natural nanostructure and biocompatibility make them excellent drug delivery platforms, while their immunogenic and modifiable surfaces position them as novel vaccine candidates and sensitive diagnostic tools (14, 15).
Compared to other nanoparticle and extracellular vesicle platforms, OMVs possess distinct advantages for biomedical applications. Unlike synthetic liposomes, which require complex chemical synthesis and lack inherent immunostimulatory properties, OMVs are naturally produced with intrinsic immunogenic components, like LPS and OMPs, that can be harnessed for vaccine development (16). In contrast to mammalian-derived exosomes, which face challenges in large-scale production, batch-to-batch variability, and complex isolation procedures, OMVs can be produced in substantial quantities through bacterial fermentation with greater reproducibility and cost-effectiveness (17). Furthermore, OMVs offer superior genetic and surface engineering capabilities compared to exosomes, allowing precise modification of cargo and surface antigens through established bacterial genetic manipulation techniques (18). While synthetic nanoparticles provide controlled physicochemical properties, OMVs combine the benefits of biological origin, including natural biocompatibility, membrane fusion capacity, and pathogen-associated molecular patterns, with the scalability and modifiability typically associated with engineered systems (19). These unique characteristics position OMVs as a promising hybrid platform that bridges the gap between synthetic and naturally-derived delivery systems.
This review provides a systematic examination of the contemporary understanding of OMV biogenesis, molecular composition, and functional diversity, while offering a critical evaluation of their emerging applications in therapeutics and diagnostics. We identify and discuss pivotal challenges in OMV production standardization, safety optimization, and clinical translation. Importantly, we underscore persistent and significant knowledge gaps concerning the fundamental mechanisms of OMV biogenesis and their multifaceted roles in microbial ecology and infection biology. Addressing these gaps through focused investigation is crucial to fully unlocking the transformative biomedical potential of OMVs.
Mechanisms of OMV production and secretion by bacteria
2
OMV biogenesis occurs under diverse growth conditions and has been observed both in vivo and in various in vitro environments, including liquid cultures, solid media, biofilms, and host cell coculture systems. The fundamental process involves localized membrane blebbing, where outward curvature of the OM leads to constriction and eventual vesicle release. Although the precise regulatory mechanisms remain incompletely characterized, current evidence indicates that OMV production is a highly regulated biological process, not a stochastic event. Recent biochemical and genetic approaches have elucidated three predominant models currently debated in the literature: the lipoprotein depletion model, the envelope stress response model, and the periplasmic accumulation model. Each proposing distinct molecular triggers for initiating membrane curvature and vesicle pinching-off. A schematic summary of OMV biogenesis is presented in Figure 1.
Schematic summary of bacterial outer membrane vesicles (OMVs) biogenesis. (A) Disruption of lipoprotein-peptidoglycan (Lpp-PG) crosslinks promotes outer membrane (OM) bulging and vesiculation. (B) Asymmetric Pseudomonas quinolone signal (PQS) in the OM leaflet induces curvature. (C) Periplasmic accumulation of misfolded proteins or lipids generates mechanical pressure. (D) Environmental factors modulate OMV biogenesis. Center: OMV budding from OM with cargo. Key components are labeled: Lpp, Lipoproteins; Lpp-PG, lipoprotein-peptidoglycan; PG, peptidoglycan; OM, outer membrane; IM, inner membrane; PQS, Pseudomonas quinolone signal.
Disruption of OM peptidoglycan crosslinking
2.1
The Gram-negative cell envelope consists of an asymmetric OM, a cytoplasmic inner membrane (IM), and a periplasmic space containing a thin PG layer (20). Structural integrity of this envelope is maintained through covalent linkages between abundant outer membrane lipoproteins (Lpp) and the PG layer. Environmental stressors can induce proteolytic cleavage of Lpp, disrupting these critical crosslinks. This destabilization promotes localized OM bulging, leading to vesicle formation via membrane curvature. Schwechheimer and colleagues provided key evidence for this mechanism by demonstrating an inverse correlation between Lpp-PG crosslinking density and OMV production, establishing that reduced tethering forces directly facilitate OMV biogenesis (6).
Membrane curvature induction via molecular enrichment
2.2
A biophysical model of OMV biogenesis, established by the Florez group, centers on the membrane-remodeling properties of Pseudomonas quinolone signal (PQS), 2-heptyl-3-hydroxy-4-quinolone, in Pseudomonas aeruginosa (P. aeruginosa) (21). As a quorum-sensing molecule, PQS preferentially accumulates in the outer leaflet of the OM, creating asymmetric lateral expansion relative to the inner leaflet. This molecular crowding generates an interleaflet tension differential that drives membrane curvature, the critical initial step in OMV budding (22). Current evidence indicates PQS promotes vesiculation through two synergistic mechanisms: chelating cations to disrupt stabilizing bridges between B-band LPS molecules, and enhancing electrostatic repulsion among the negatively charged LPS moieties (23). Although the PQS model represents one of the most mechanistically characterized pathways, its applicability remains phylogenetically constrained to P. aeruginosa. This limitation underscores the need to identify analogous curvature-inducing molecules in other Gram-negative species that may employ distinct molecular strategies for membrane remodeling and OMV production.
Periplasmic accumulation-induced vesiculation
2.3
The periplasmic compartment serves as another critical site for OMV biogenesis. Accumulation of misfolded proteins or peptidoglycan fragments in this confined space, due to stressors like temperature shifts or oxidative stress, generates substantial turgor pressure against the outer membrane. This mechanical stress initiates a cascade: compromising membrane integrity by disrupting OM-PG tethering, creating areas of local detachment, and ultimately driving vesicle formation through physical displacement. The resulting structural instability leads to membrane blebbing and OMV release. Roier et al. directly linked phospholipid accumulation in the outer membrane to vesiculation by showing that inactivation of the VacJ/Yrb ABC transport system in Haemophilus influenzae increases OMV production (24). This phospholipid-dependent mechanism appears evolutionarily conserved and is subject to precise genetic regulation, for example, through the ferric uptake regulator (Fur) system, illustrating how bacteria actively modulate OMV production in response to environmental cues.
Additional regulatory factors
2.4
Beyond the established models, emerging evidence indicates that various exogenous factors significantly modulate OMV biogenesis. Environmental stimuli such as temperature, ultraviolet radiation, and nutritional factors have been shown to regulate OMV production both quantitatively and qualitatively, affecting vesicle yield, size distribution, and molecular cargo composition (25, 26). This underscores that bacterial vesiculation is a dynamic response to extracellular conditions.
While the models described above provide valuable frameworks, none alone fully explains the complexity of OMV production across all contexts. The process is under intricate genetic control, with numerous genes implicated in its biosynthesis and regulation (summarized in Table 1). For example, utilizing a novel curvature-sensing fluorescent peptide (nFAAV5-NBD) in a genome-wide screen of Shewanella vesiculosa, Hiromu’s team identified several key genetic regulators of vesiculation, including putative dipeptidyl carboxypeptidase, glutamate synthase β-subunit, LapG protease, metallohydrolase, RNA polymerase sigma-54 factor, inactive transglutaminase, PepSY domain-containing protein, and Rhs-family protein (27). Importantly, elucidating the fundamental mechanisms of OMV biogenesis holds substantial practical value. A deeper mechanistic understanding is crucial for rationally optimizing production yields and cost-efficiency, which represents a major bottleneck in advancing therapeutic OMV applications.
Main composition of OMVs
3
Gram-negative bacteria naturally release OMVs, which encapsulate approximately 0.2%-0.5% of their OM and periplasmic components to form sophisticated, biomolecule-rich nanostructures (28). Structurally, OMVs maintain a characteristic bilayer architecture composed primarily of phospholipids, fatty acids, and cholesterol. This lipid-based framework provides both structural integrity and a vehicle for intercellular communication and virulence factor delivery. Unlike mammalian extracellular vesicles, OMVs contain unique bacterial components including LPS and PG (29). The vesicular cargo is composed of membrane-derived signaling lipids, genetic material, protein effectors including enzymes and toxins, as well as pathogen-associated molecular patterns. This assemblage of components collectively empowers OMVs to mediate a wide array of essential functions. Collectively, this multifaceted composition underpins the remarkable functional versatility of OMVs in microbial physiology and host-pathogen interactions, establishing them as pivotal mediators in bacterial survival and pathogenesis.
LPS
3.1
LPS is a complex molecule localized in the outer leaflet of the Gram-negative bacteria OM. Historically, LPS was first described as “endotoxin” by Pfeiffer, who characterized it as a highly toxic component of Vibrio cholera (30). Consequently, LPS constitutes the primary virulence factor in OMVs derived from Gram-negative bacteria. Notably, LPS exhibits significantly enhanced toxicity when associated with OMVs compared to its free form (31). This enhanced pathogenicity is exemplified by the unique capacity of OMVs to transport LPS across the blood-brain barrier, inducing inflammation in both the peripheral and central nervous system - a feat unattainable by purified LPS alone (32). The potent inflammatory potential of OMV-associated LPS is further demonstrated by observations that intranasal administration of P. aeruginosa OMVs elicits stronger pulmonary inflammation than equivalent doses of purified LPS, underscoring OMVs’ efficient delivery capability (33). At the molecular level, LPS recognition occurs via TLR4-mediated signaling: the LPS-TLR4 complex recruits TIRAP and MyD88 to activate IRAK and TRAF6, ultimately stimulating NF-κB and MAPK pathways (13). Notably, OMV-mediated delivery can bypass this surface recognition, facilitating LPS entry into the host cell cytosol where it triggers caspase-11-dependent immune responses (34). The 3-O-desacyl-4’ monophosphoryl lipid A (MPL, Corixa), derived from LPS of Salmonella Minnesota R595, is an approved adjuvant in human vaccines (35). These findings collectively emphasize the dual nature of OMV-associated LPS: while its endotoxin activity poses a challenge for therapeutic safety, its potent immunostimulatory properties, when strategically modulated through bioengineering, present significant opportunities for novel vaccine design.
PG
3.2
PG is the main component of the prokaryotic cell wall. It is a multi-layered, network-like macromolecular formed by the polymerization of N-acetylglucosamine (GlcNAc), N-acetylmuramic (MurNAc), and four to five amino acid short peptides. Notably, the human receptors NOD1 exhibits remarkable specificity for the GlcNAc-MurNAc tripeptide motif containing diaminopimelate (GM-TriDAP), a structural signature predominantly associated with Gram-negative bacterial PG (36). Upon binding this ligand, NOD1 initiates a signaling cascade via its caspase activation and recruitment domain (CARD). This involves recruiting receptor-interacting protein 2 (RIP2) to activate both NF-κB and MAPK pathways and ultimately driving the production of proinflammatory cytokines such as IL-8 (37). Intriguingly, while extracellular administration of synthetic or natural NOD1 agonists fails to activate NOD1 signaling in non-phagocytic epithelial cells, Kaparakis et al. demonstrated that Gram-negative bacterial PG delivered via OMVs can efficiently enter these cells through lipid raft-mediated endocytosis, thereby triggering robust NOD1-dependent immune responses (38). This OMV-mediated delivery mechanism provides critical insights into how Gram-negative bacteria may promote inflammatory pathology during infection by subverting normal immune recognition pathways.
Enzymes and toxins
3.3
Proteins constitute a functionally diverse and critical component of OMVs, encompassing virulence factors, antibiotic resistance determinants, and host interaction mediators. These vesicle-associated proteins facilitate intercellular communication and regulation, thereby enhancing bacterial survival and dissemination capabilities.
A key facet of OMV functionality is their role as specialized vehicles for delivering bacterial toxins and enzymes into eukaryotic cells. Enterotoxigenic Escherichia coli exemplifies this mechanism by packaging heat-labile enterotoxin (LT) within composite vesicular structures derived from both OM and periplasmic components (39). Similarly, P. aeruginosa employs OMVs as an alternative secretion pathway for multiple virulence factors, including phospholipase C, protease, hemolysin, and alkaline phosphatase - enzymes that collectively contribute to the pathogenesis of Pseudomonas infections (40). The periodontal pathogen Aggregatibacter actinomycetemcomitans employs a related strategy by displaying leukotoxin on OMV surfaces, facilitating its involvement in both localized aggressive periodontitis and systemic disease (41). Bacteroides thetaiotaomicron targets host immune cells via OMVs harboring bacterial sulfatase activity, demonstrating how OMVs-associated enzymes can promote inflammatory immune stimulation in genetically susceptible hosts (42). It is worth noting that clyA, a pore-forming cytotoxin expressed by Enterobacteriaceae, is also secreted via OMVs, a finding with important implications for biotechnology applications (43). The systematic characterization of OMV-associated proteins remains a crucial research direction, offering insights into fundamental microbial pathogenesis and revealing potential avenues for therapeutic applications.
Nucleic acids
3.4
OMVs serve as natural carriers for various nucleic acid species, including both DNA and RNA, which play crucial roles in bacterial gene regulation and intercellular genetic exchange. The incorporation of DNA into OMVs may occur through multiple pathways, ranging from spontaneous encapsulation during cell lysis events to selective packaging mechanisms (44). Compelling evidence for non-random DNA packaging comes from Bitto et al., who demonstrated preferential enrichment of specific chromosomal regions, encoding virulence factors, stress response elements, antibiotic resistance determinants, and metabolic enzymes, within P. aeruginosa OMVs. Their findings revealed a distinctive spatial organization, with most DNA associated with the vesicle surface and smaller quantities contained internally (45). The role of OMVs in mediating HGT is well-documented across multiple bacterial species. Dorward et al. established that Neisseria gonorrhoeae utilizes OMVs as vectors for intercellular plasmid transfer under physiological conditions (46). This phenomenon has been corroborated by numerous studies demonstrating OMV-facilitated dissemination of genetic material encoding diverse functions, including metabolic pathways, virulence traits, and antibiotic resistance markers (47).
In addition to DNA, various RNA species are packaged into OMVs, and their contributions to host-microbe interactions have been confirmed in several studies (48). The majority of OMV-associated RNAs are small RNAs (sRNAs), including microRNAs (miRNAs), and miRNA-sized sRNAs (msRNAs) (48, 49). A striking example is provided by Porphyromonas gingivalis, whose OMVs deliver sRNA-23392 to promote oral squamous cell carcinoma progression by inhibiting desmocollin-2 expression (50). Despite these advances, the molecular mechanisms governing the selective sorting of RNA into OMVs remain poorly understood, representing a critical knowledge gap in the field.
Functions of OMVs
4
OMVs serve as multifunctional biological nanoparticles pivotal to bacterial physiology and host-microbe interactions. Bearing diverse cargo, these sophisticated nanostructures mediate both interbacterial communication and cross-kingdom signaling. As the sole known mechanism for delivering hydrophobic compounds to host cells, OMVs have been formally classified as type 0 secretion systems (51). Their functional spectrum encompasses nutrient acquisition, environmental adaptation, and antibiotic resistance dissemination through HGT. In pathogenic contexts, OMVs act as precision delivery vehicles for virulence factors, facilitating host cell invasion while simultaneously modulating immune responses. They also contribute significantly to biofilm architecture and stability, enhancing bacterial community resilience. This remarkable functional versatility derives from their composition dynamically adapts to environmental cues, enabling homeostasis and ecological responsiveness. The capacity of OMVs to participate in such diverse biological processes underscores their fundamental role in microbial ecology and pathogenesis.
Bacterial defense mechanisms
4.1
OMV biogenesis represents an evolutionarily conserved defense strategy employed by bacteria to cope with environmental stressors. Under nutrient limitation, antibiotic pressure, or other extreme challenges, bacteria actively modulate OMV production to enhance survival through multiple protective mechanisms (52). These include neutralizing membrane-targeted threats, scavenging harmful extracellular compounds, and dynamically remodeling vesicular protein composition (53). Vibrio fischeri upregulates its major OMP OmpU under acidic pH encountered during host colonization. This pH-dependent protein restructuring significantly alters OMV cargo composition to facilitate host-symbiont interactions (54). Pseudomonas putida rapidly releases OMVs upon stress exposure, increasing cell surface hydrophobicity to promote biofilm formation and persistence (55). OMVs from enterotoxigenic Escherichia coli (E. coli) can quickly and irreversibly bind T4 bacteriophage, allowing parental bacteria to effectively evade OM-target attack (56).
OMVs function as a critical defense mechanism against antibiotics by shielding bacteria from antibiotic-induced damage and by acting as export vehicles for antibiotic-degrading enzymes. For instance, the Antarctic bacterium Pseudomonas syringae produces OMVs that mimic parent cell, effectively neutralizing membrane-active antibiotics (57). Similarly, E. coli secretes the metallo-β-lactamase NDM-1 via OMVs. These vesicles act as extracellular reservoirs of the enzyme to degrade β-lactam antibiotics in the environment and protect neighboring bacterial populations (58). Additionally, OMVs production increases when bacteria are briefly exposed to low doses of antibiotics, as OMVs can reduce antibiotic concentration by interacting with antibiotics. Specifically, sublethal doses of β-lactams antibiotics can induce hypervesiculation in Salmonella enterica sv. Typhi. The newly generated OMVs exhibit high affinity for polymyxin B, sequestering it and thereby enhancing bacterial survival against this agent (59). These examples underscore how OMV production serves as a versatile, rapid-response defense system that enhances bacterial survival across varied ecological niches and stress conditions.
Communication within bacterial populations
4.2
OMVs are vital for intercellular signaling, carrying various signaling molecules to coordinate group behavior, a process referred to as quorum sensing (QS). The differential content of their structural components and the selective packaging of biomolecules indicate that OMVs may function as “communicasomes,” mediating communication not only within the bacterial communities but also across different domains of life.
OMV enables the transfer of information between bacteria, which affects biological processes such as population behavior, growth, and metabolism. A prime example is P. aeruginosa, in which the hydrophobic signal PQS cannot diffuse freely across bacterial membranes and therefore relies on OMV packaging for transport. Studies indicate that more than 86% of the PQS produced by P. aeruginosa is packaged into OMVs (60). Removal of these OMVs from the bacterial community disrupts intercellular communication and suppresses PQS-controlled group behaviors. Similarly, the long-chain N-acyl-homoserine lactones, such as N-hexadecanoyl-L-homoserine lactone used by Paracoccus denitrificans, are disseminated via OMVs. These vesicles fuse selectively with recipient cells to facilitate signal delivery (61). The marine pathogen Vibrio harveyi packages the hydrophobic QS molecule CAI-1 into OMVs, stabilizing it in aqueous environments and extending its signaling range (62).
Beyond signaling, OMVs mediate bacterial antagonism by delivering antimicrobial cargo to eliminate competitors. For instance, P. aeruginosa secretes peptidoglycan hydrolases via OMVs to degrade rival bacteria (63). Myxococcus xanthus produces OMVs laden with proteases, phosphatases, and hydrolases that lyse E. coli cells upon membrane fusion (64). These findings align with the concept of OMVs as predatory tools in microbial warfare (65). Moreover, OMVs have also been identified as mediators of HGT in interspecies communication. Plasmids packaged within OMVs serve as the basis for HGT, thereby facilitating the spread of beneficial mutations. For example, OMVs from Acinetobacter baylyi (A. baylyi) transfer DNA to E. coli and other A. baylyi cells (66). Although interspecific exchange of OMVs is common in microbial communities, the mechanisms underlying how bacteria recognize and take up OMVs from different species remain elusive.
Communication between host and bacteria
4.3
OMVs are critical mediators of host-bacteria interactions, primarily through several key mechanisms, including host cell invasion, targeted delivery of virulence factors, and immunomodulation. As discussed above, OMVs carry numerous virulence factors, such as toxins, degrative enzymes, and adhesins. Crucially, compared to their free forms, OMV-associated virulence factors exhibit enhanced efficacy due to increased local concentration and protection from degradation. Active Cholera toxin can be delivered to intestinal epithelial cells by OMVs avoiding degradation by intestinal proteases (67). Certain periodontal pathogen-derived OMVs carry sRNAs that suppress cytokine expression in Jurkat T cells, highlighting their capacity for immune modulation (68).
Components like LPS and flagellin act as pathogen-associated molecular patterns, directly modulating host immune pathways. Jang et al., found that the intraperitoneal administration of OMVs triggered significant increases in TNF-α and IL-6 levels in both serum and bronchoalveolar lavage fluid (69). Additionally, OMVs are small enough to even cross the blood-brain barrier (BBB) and induce an immune response. Studies have shown that extracellular RNAs in periodontopathogenic OMVs can successfully cross the BBB and prompt TNF-a production in murine brain (70). Mature OMVs are capable of adhering to the host cell membrane following their release and detachment, and subsequently enter the cell via endocytosis or membrane fusion to release their cargo at specific locations (71). This process can be influenced by the factors such as the size of OMVs themselves, LPS types, and glycoproteins (72, 73). The protective membrane structure of OMVs preserve the stability of these molecules, enabling them to modulate interactions and responses between bacteria and cells via long-distance transmission. Leveraging this property, OMVs can be harnessed for the treatment of intracellular infections, and antimicrobial-loaded OMVs enter host cells infected by pathogenic bacteria through endocytic or membrane fusion pathways.
OMV can act as potent mediators to enhance vaccine and drug delivery by efficiently accessing the lymphatic system and stimulating immune responses. This efficacy stems from their ability to fuse easily with target cell membranes, their long circulation time in the bloodstream, as well as their natural capacity for loading biomolecules (74). Yersinia enterocolitica OMVs elicit robust humoral and mucosal immune responses in murine models (75). Engineered OMVs have demonstrated remarkable antitumor effects by inducing IFN-γ-dependent tumor regression, enhancing antitumor cytokines production, and displaying tumor antigens to stimulate antigen-specific immunity (76, 77). These strategies effectively inhibit tumor growth and metastasis while establishing long-term immunological memory. The versatility of OMVs across infectious disease and oncology underscores their potential as next-generation biotherapeutic platforms.
Biological modification and application of OMVs
5
Recent years have witnessed significant progress in the biological modification of OMVs, revolutionizing their potential in biomedical and biotechnological applications. Current strategies for engineering OMVs primarily focus on three key objectives: attenuating virulence factors to enhance safety, optimizing production yields for scalable manufacturing, and functionally enhancing vesicles to maximize therapeutic efficacy. Genetic engineering has emerged as a powerful tool for precisely tailoring OMV properties through the targeted manipulation of bacterial genes that regulate vesicle biogenesis and cargo sorting. By engineering the host bacterial strain, researchers can systematically reprogram OMV characteristics while maintaining their inherent biological functions. Concurrently, the integration of synthetic nanomaterials with natural OMV components has given rise to a new generation of biohybrid nanostructures.
Licensed and clinical-stage OMV vaccines
5.1
OMV-based vaccines (VA-MENGOC-BC, MenBvac, MeNZB, and 4CMenB) have been employed worldwide in response to epidemic meningococcal disease outbreaks caused by Neisseria meningitidis. Meanwhile, several OMV-based candidates have advanced into clinical development, targeting a range of pathogens including bacteria and viruses.
Beyond licensed meningococcal vaccines, modified meningococcal OMVs have demonstrated versatility as platform technology in clinical development. A clinical trial in Brazil (MenB-Bio vaccine) evaluated a tailored OMV-based vaccine containing detergent-treated OMVs and detoxified lipooligosaccharide (dLOS) derived from two prevalent local MenB strains. The Phase II/III study demonstrates the clinical translation of modified, strain-specific OMVs designed to address regional meningococcal epidemiology (78). Genetically engineered meningococcal OMVs with constitutive expression of FetA (MenPF-1) completed Phase I clinical trials, showing enhanced bactericidal antibody responses beyond PorA-mediated immunity (79). The Walter Reed Army Institute of Research (WRAIR) has conducted pioneering clinical trials using genetically engineered native OMVs (nOMVs) derived from meningococci without exposure to detergent or denaturing agents. Early Phase I trials evaluated intranasal administration of nOMVs derived from capsule-negative strain 9162, demonstrating safety and induction of cross-reactive bactericidal antibodies despite the presence of LPS (80, 81). Subsequently, WRAIR developed intramuscular nOMV vaccines using strains with genetic modifications to enhance safety and immunogenicity. A phase 1 trial assessed a candidate prepared from a lpxL2(−) synX(−) mutant of strain 44/76 with stabilized opcA expression. This vaccine showed an acceptable safety profile and induced bactericidal antibodies, supporting its potential to provide protection against group B meningococcus (82). Furthermore, meningococcal OMVs are being explored as delivery platforms for heterologous antigens from non-bacterial pathogens. For instance, the SARS-CoV-2 Spike protein displayed on Neisseria meningitidis OMVs (Avacc 10), developed by IntraVacc B.V., has received authorization to enter Phase I clinical trials (NCT05604690).
OMV vaccines against Neisseria gonorrhoeae have garnered significant clinical interest in recent years. The 4CMenB and MeNZB vaccines have shown a protective effect against gonorrhoea, reducing acquisition by 30-40% (83). Additionally, a gonococcal generalized modules for membrane antigens (GMMA) vaccine has completed preliminary preclinical studies and is reported in the literature to be advancing toward clinical evaluation (84). GMMA are genetically engineered OMVs designed to achieve hypervesiculation and tailored membrane composition. This platform features a simple manufacturing process, leading to affordable vaccines (85). The first clinically tested GMMA vaccine, Shigella sonnei 1790GAHB, demonstrated safety and immunogenicity in phase I/II trials, including a human challenge model, though it failed to confer protection, likely due to insufficient O-polysaccharide dosing (86, 87). This finding prompted the development of a next-generation version with tenfold higher antigen content. The optimized GMMA has now been formulated into a tetravalent vaccine combining Shigella flexneri serotypes 1b, 2a, and 3a, which is currently undergoing phase I/II clinical evaluation (NCT05073003). A Phase I trial (SALVO study, ISRCTN51750695) evaluated the safety and immunogenicity of the bivalent invasive non-typhoidal Salmonellae (iNTS)-GMMA vaccine in healthy adults. The vaccine was well-tolerated with no serious adverse events and successfully induced persistent functional antibody responses. Despite the extensive reporting of OMV-based bacterial vaccine candidates in the scientific literature, most remain in the preclinical stage of development.
Strategies for lipid A-targeted attenuation of OMVs
5.2
The lipid A moiety of LPS constitutes the predominant and most extensively studied virulence factor in OMVs, making it the primary target for detoxification. Lipid A consists of a conserved (β1→6)-linked glucosamine disaccharide backbone, typically phosphorylated at the 1 and 4’ positions and acylated at the 2 and 3 positions of each sugar unit (88). Controlled modulation of lipid A architecture can balance immunogenicity with safety. Its immunostimulatory capacity critically depends on the acylation pattern, and hexa-acylated forms exhibit maximal cytokine-inducing activity via TLR4 activation, whereas tetra-acylated variants often act as TLR4 antagonists (89). Other structural features, including phosphate group number and acyl chain length, further modulate lipid A’s inflammatory potential (88). In E. coli, the Raetz pathway orchestrates lipid A biosynthesis through nine conserved enzymatic steps mediated by LpxA-LpxD, LpxH, LpxK, LpxM, and WaaA (90).
It is generally consensus that deleting late acyltransferases attenuates pathogens by leading to underacylated, less toxic lipid A species (91). LpxM and LpxL belongs to the lauryl/myristyl acyltransferases family involved in lipid A biosynthesis, and inactivation of either enzyme generates penta- or tetra-acylated lipid A species (92). In the Shigella flexneri 2a human challenge strain, deletion of the late acyltransferases msbB1 and msbB2 genes results in underacylated lipid A production (91). The 4’-phosphatase LpxF from Francisella tularensis dephosphorylates lipid A at the 4’ position in an E. coli LpxM mutant, and it does not act on wild-type hexa-acylated lipid A (93). LpxE from Francisella novicida selectively removes the 1-phosphate group of lipid A (94). Additionally, the palmitoyl transferase PagP in E. coli and Salmonella adds a secondary palmitoyl chain (C16) at the 2-position of lipid A, and such palmitoylated LPS exhibits attenuated signaling through the TLR4/MD-2 complex (95). In Salmonella enterica, Pag L hydrolyzes the ester bond at the 3-position of lipid A, thereby modulating lipid A recognition by the TLR4/MD-2 complex (96).
While the aforementioned lipid A engineering strategies provide powerful tools for endotoxin attenuation, translating these modifications into clinically viable OMV therapeutics requires navigating complex regulatory frameworks for endotoxin management. Regulatory agencies impose strict endotoxin limits for parenteral biologics, with FDA guidelines typically requiring ≤5 EU/kg/hour for systemic administration. However, these thresholds, established for conventional pharmaceuticals, may not appropriately reflect the risk-benefit profile of OMV-based immunotherapeutics where controlled TLR4 activation is often desirable for adjuvant effects (97). Application-specific endotoxin specifications are emerging: OMV vaccines may tolerate higher endotoxin levels to leverage immunostimulation, while OMV drug carriers require maximal detoxification. Critical regulatory considerations include establishing validated endotoxin quantification methods suitable for vesicular formulations, as standard limulus amebocyte lysate/tachypleus amebocyte lysate (LAL/TAL) assays may underestimate or overestimate activity in complex lipid matrices. Demonstrating batch-to-batch consistency in lipid A structure and TLR4 activity through orthogonal analytical methods such as mass spectrometry and cell-based potency assays is essential for regulatory approval. Correlating in vitro endotoxin measurements with in vivo pyrogenicity and safety through rabbit pyrogen tests or monocyte activation tests remains a regulatory requirement, though these methods have limitations for complex biological products. Defining acceptable endotoxin ranges that balance safety with therapeutic efficacy for each OMV application requires extensive preclinical dose-ranging studies with comprehensive toxicity assessment. The establishment of OMV-specific guidance, currently absent from key regulatory agencies, would facilitate standardized approaches to endotoxin risk assessment and management.
Cutting-edge technologies for functional enhancement of OMVs
5.3
Due to their nanoscale dimensions, OMVs possess ideal biophysical properties for immune system engagement. They are readily internalized by antigen-presenting cells, efficiently trafficked through lymphatic vessels, and demonstrate superior capabilities for antigen delivery, thereby potently stimulating both humoral and cellular immune responses. Recent advances in bioengineering have further expanded this therapeutic potential. The convergence of OMV biology with cutting-edge nanotechnology and genetic engineering is now opening new frontiers in biomedical research, creating unprecedented opportunities for the development of next-generation vaccines and immunotherapies.
Cell membrane-coated nanoplatforms for advanced biomedical application
5.3.1
Cell membrane coating technology has emerged as a powerful strategy in biomedical research, particularly for vaccine development and targeted therapeutics. Coating synthetic nanoparticles with OMVs preserves the complex biological characteristics of bacteria and mimics the natural process of antigen presentation to the immune system. OMVs derived from Bordetella bronchiseptica were conjugated with PEGylated nano-Rehmannia glutinosa polysaccharide to synthesize a nanovaccine, which exhibits enhanced protective immunity against bacterial infection by modulating T-cell receptor signaling and Th1/Th2/Th17 differentiation (98). OMVs from E. coli Nissle 1917 served as nanoreactors to fabricate biomimetic copper sulfide nanoparticles, exhibiting high photothermal conversation efficacy, excellent photostability, and significant tumor-targeting capability (99). The OMV-based camouflage of gold nanocages facilitated the targeted uptake of dexmedetomidine-loaded nanoparticles into M1-like senescent macrophages, leading to a marked increase in drug delivery efficiency (100). In summary, coating nanocarriers with OMVs enhances the immunostimulatory effects of free OMVs by improving their stability and targeting capability. These biomimetic systems combine the advantages of artificial nanoparticles with the biological functionality of OMVs. The OMV-coating strategy offers several key benefits. It can enhance stability of therapeutic payloads, improve targeting specificity through natural membrane proteins, and amplify immunostimulatory effects via pathogen-associated molecular patterns. Such hybrid platforms bridge the gap between synthetic and biological systems. However, the extent of these improvements depends on the selected nanoparticle core, underscoring the need for optimal nanocarrier design.
OMVs as versatile drug delivery vehicles
5.3.2
The concept of utilizing OMVs for drug delivery was first proposed in the late 1970s and early 1980s. Since then, OMVs have emerged as promising nanocarriers capable of encapsulating drugs or bioactive molecules, enabling targeted delivery to specific tissues or cells while enhancing drug bioavailability and therapeutic efficacy. Therapeutic agents can be loaded into OMVs via either energy-dependent (active loading) or energy-independent (passive loading) techniques (101). A key advancement in OMV-based delivery involves genetically modifying to display functional proteins on their surface. Outer membrane protein A (OmpA) has been used to stably display phosphotriesterase (PTE) from Brevundimonas diminuta while preserving its enzymatic activity (102). Cytolysin A (clyA) has been engineered to display affiEGFR-GALA peptides on OMVs derived from an endotoxin-reduced E. coli W3110 strain. These engineered OMVs serve as potential carriers for cell-specific drug delivery, particularly against cancer cells, as they are capable of reaching the cytoplasm while escaping the endosome due to the incorporation of a fusogenic GALA peptide. Selecting an appropriate anchor protein is crucial for ensuring effective surface display of recombinant proteins (103). A summary of commonly used anchors is provided in Table 2.
Therapeutic mRNA vaccines have advanced rapidly in recent years. Concurrently, OMVs have been proved to be a promising mRNA delivery platform through genetic engineering. For example, OMVs genetically decorated with RNA-binding and lysosomal escape proteins (OMV-LL) can deliver mRNA antigens, markedly inhibiting tumor progression in models of metastatic and subcutaneous colon cancer (104). The successful demonstration of OMV-based mRNA delivery heralds a significantly advance in next-generation vaccine and immunotherapies. Meanwhile, bioconjugation systems, such as the SpyCatcher-SpyTag enables covalent protein coupling to OMV surfaces, offering a modular and efficient approach for functionalization. Additionally, CRISPR-based systems have been leveraged to enhance the efficiency of targeting and packaging therapeutic molecules into OMVs (105). These technologies further expand the possibilities for customizable OMV engineering, allowing for multifunctional drug delivery and immune modulation.
Summary and outlook
6
Challenges in OMV-based application
6.1
In clinical applications, the approval of several OMV-based meningococcal vaccines for human use highlights their potential, with promising prospects in vaccine development, targeted drug delivery, and tumor therapy. However, the clinical implementation of OMVs faces multiple substantial barriers.
OMV production is inherently constrained by bacterial physiological factors including microenvironmental conditions, viability status, and growth phase, leading to low yields that require costly optimization through culture condition refinement or genetic engineering approaches. Purification methods involve isolation via techniques such as ultracentrifugation, filtration, or chromatography, and these processes may introduce impurities including free proteins that affect OMV purity and safety.
Beyond production yield and purity, achieving batch-to-batch consistency represents a fundamental manufacturing challenge for OMV therapeutics. The biological nature of OMV production introduces inherent variability in critical quality attributes including vesicle size distribution, protein and lipid composition, antigen loading efficiency, and immunogenic potency. Controlling this requires stringent management of fermentation parameters such as pH, temperature, dissolved oxygen, growth phase, and media composition. This control must operate within narrowly defined ranges following validated bioprocess protocols. Furthermore, implementation of Process Analytical Technology (PAT) for real-time monitoring and adaptive control of OMV production has emerged as a promising approach to reduce batch variability. Subsequently, comprehensive release testing must encompass physical characterization through dynamic light scattering for size and polydispersity index, nanoparticle tracking analysis for concentration, biochemical profiling including proteomics and lipidomics, endotoxin quantification, and functional potency assays. Despite these measures, the establishment of certified reference standards for comparative batch analysis remains a critical unmet need in the field.
Current manufacturing compliance with cGMP requirements presents additional challenges, as existing frameworks are designed for mammalian cell culture or chemical synthesis rather than bacterial fermentation-derived vesicles. The lack of industry-wide standardization protocols and OMV-specific regulatory guidance from major agencies including FDA, EMA, and NMPA currently impedes inter-laboratory reproducibility and necessitates case-by-case regulatory evaluation. Establishing distinct regulatory pathways for OMV therapeutics rather than forcing classification into existing vaccine or drug categories is essential for streamlining approval processes while ensuring appropriate safety oversight.
Comprehensive safety assessment is paramount for clinical translation of OMV-based therapeutics. The immunostimulatory properties that make OMVs attractive for vaccines and immunotherapy simultaneously pose immunotoxicity risks requiring careful evaluation. Acute concerns include cytokine release syndrome from excessive TLR activation, particularly with high-dose or repeated administration, necessitating establishment of maximum tolerated doses and monitoring of pro-inflammatory cytokines including IL-6, TNF-α, and IL-1β in preclinical studies. Systemic inflammatory response syndrome risk is elevated in immunocompromised patients or those with pre-existing inflammatory conditions, necessitating patient stratification strategies and potentially exclusion criteria in early-phase clinical trials. Additionally, potential autoimmune reactions may arise if OMV-displayed antigens exhibit molecular mimicry with host epitopes, a concern that requires comprehensive epitope mapping and immunological profiling during preclinical development. Hypersensitivity reactions upon repeated exposure warrant investigation through appropriate animal models that recapitulate chronic dosing regimens. Mitigation strategies include lipid A detoxification as discussed in Section 4.1, dose optimization through careful pharmacokinetic and pharmacodynamic studies, premedication protocols to dampen excessive inflammatory responses, and comprehensive immunotoxicity panels including cytokine profiling, complement activation assays, and immunophenotyping in preclinical safety assessments.
The long-term safety profile of repeated OMV administration, particularly relevant for chronic immunotherapy regimens, remains inadequately characterized. Critical knowledge gaps include potential for immune tolerance or exhaustion with chronic exposure that could diminish therapeutic efficacy over time. Risk of persistent inflammation or organ damage in tissues with high OMV accumulation requires histopathological examination in extended toxicology studies. The impact of OMV administration on microbiome composition and host-microbe homeostasis represents an emerging safety consideration, as disruption of commensal bacterial communities could have unintended metabolic and immunological consequences (106). Reproductive and developmental toxicity for long-term treatments must be evaluated through standard reproductive toxicology study designs. Genotoxicity concerns related to bacterial DNA fragments potentially carried by OMVs require assessment. Establishing these safety parameters requires rigorous long-term toxicology studies in relevant animal models with extended observation periods of at least six months, repeated-dose regimens that accurately mimic proposed clinical protocols, and comprehensive histopathological examination of all major organ systems. For approved OMV products, pharmacovigilance programs with long-term patient follow-up are essential to detect rare or delayed adverse events not apparent in limited clinical trials.
The clinical translation of OMVs necessitates coordinated advances in scalable PAT-controlled bioprocessing, harmonized analytical and release criteria, comprehensive and predictive safety science, and the development of a tailored regulatory science framework. It is still worth noting that the targeting ability of OMVs and their circulation time in the bloodstream will influence their efficacy in clinical applications. If OMVs are easily cleared from the bloodstream or exhibit poor targeting ability, their antimicrobial or antitumor efficacy will be compromised.
Outlook
6.2
Notably, many clinical adjuvants, such as MPL and CpG, originate from bacterial components, and bacterial-based cancer therapies further highlight bacteria as valuable source of immunostimulatory agents. In contrast to live bacteria, OMVs are non-replicating and thus inherently safer, yet they retain natural immunogenicity and can be systematically engineered. Rather than being mere bacterial byproducts, OMVs represent versatile immunotherapeutic platforms beyond single-purpose vaccines. By integrating rapidly evolving synthetic biology tools with growing insights in microbiology and immunology, it is becoming feasible to precisely tailor OMV composition, targeting, and functionality. The OMV-based platforms allow for systematic and independent modification of both internal and surface-displayed antigens due to their modular nature. This flexibility enables rapid responses to emerging infectious threats, such as COVID-19, and other unforeseen public health challenges.
The characteristics and clinical applications of different OMV-based vaccine types are summarized in Table 3. The successful clinical application of OMVs represents not an endpoint but the inception of a broader paradigm shift in biomedical nanotechnology. The path forward requires deepening fundamental insights into OMV biogenesis and host interaction dynamics, coupled with the establishment of robust, scalable manufacturing and tailored safety-assessment frameworks. By uniting these foundational advances with translational engineering, OMV-based platforms are positioned to fulfill their potential as a versatile and transformative tool in precision medicine.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chatterjee SN Das J . Electron microscopic observations on the excretion of cell-wall material by Vibrio cholerae. J Gen Microbiol. (1967) 49:1–11. doi: 10.1099/00221287-49-1-1, PMID: 4168882 · doi ↗ · pubmed ↗
- 2Namork E Brandtzaeg P . Fatal meningococcal septicaemia with “blebbing” meningococcus. Lancet. (2002) 360:1741. doi: 10.1016/S 0140-6736(02)11721-1, PMID: 12480427 · doi ↗ · pubmed ↗
- 3Briaud P Carroll RK . Extracellular vesicle biogenesis and functions in gram-positive bacteria. Infect Immun. (2020) 88:e 00433–20. doi: 10.1128/IAI.00433-20, PMID: 32989035 PMC 7671900 · doi ↗ · pubmed ↗
- 4Liu H Zhang Q Wang S Weng W Jing Y Su J . Bacterial extracellular vesicles as bioactive nanocarriers for drug delivery: Advances and perspectives. Bioact Mater. (2022) 14:169–81. doi: 10.1016/j.bioactmat.2021.12.006, PMID: 35310361 PMC 8892084 · doi ↗ · pubmed ↗
- 5Zhu Z Antenucci F Villumsen KR Bojesen AM . Bacterial outer membrane vesicles as a versatile tool in vaccine research and the fight against antimicrobial resistance. m Bio. (2021) 12:e 0170721. doi: 10.1128/m Bio.01707-21, PMID: 34372691 PMC 8406158 · doi ↗ · pubmed ↗
- 6Schwechheimer C Kulp A Kuehn MJ . Modulation of bacterial outer membrane vesicle production by envelope structure and content. BMC Microbiol. (2014) 21:324. doi: 10.1186/s 12866-014-0324-1, PMID: 25528573 PMC 4302634 · doi ↗ · pubmed ↗
- 7Orench-Rivera N Kuehn MJ . Environmentally controlled bacterial vesicle-mediated export. Cell Microbiol. (2016) 18:1525–36. doi: 10.1111/cmi.12676, PMID: 27673272 PMC 5308445 · doi ↗ · pubmed ↗
- 8Noundou VL Levy A Modla S Yu Y Qu J Hanson TE . Chlorobaculum tepidum outer membrane vesicles may transport biogenic elemental sulfur. Appl Environ Microbiol. (2025) 91:e 0101925. doi: 10.1128/aem.01019-25, PMID: 40530856 PMC 12285230 · doi ↗ · pubmed ↗