Croton buiquensis (Euphorbiaceae), a new species from northeastern Brazil, its phylogenetic placement, and niche modeling
Joesili C.P. Oliveira, Karen Y. Suarez-Contento, Ricarda Riina, Sarah M. Athiê-Souza

TL;DR
A new species of Croton, found in northeastern Brazil, is described and shown to be critically endangered.
Contribution
The discovery and description of Croton buiquensis, a new and critically endangered species in the Croton genus.
Findings
Croton buiquensis is a new species distinct from C. pedicellatus based on morphological and molecular data.
Ecological niche modeling suggests the species may occur in the Agreste region of Pernambuco.
This is the first record of multifid styles in Croton section Pedicellati.
Abstract
A new species of Croton, microendemic of the dry forests of northeastern Brazil in the state of Pernambuco, is here described. Morphological and ITS molecular data provide evidence for placing C. buiquensis sp. nov. as part of Croton section Pedicellati. Specimens of C. buiquensis had been previously identified in herbaria as C. pedicellatus, but the former can be distinguished from the latter by the hispid indumentum of the leaf blade, the ovate shape of the petals of staminate flowers, pistillate flowers with 4-fid to multifid styles, and a crossbow-shape caruncle. Evidence from ecological niche modeling points to other areas in the state of Pernambuco, specifically the “Agreste” region, as likely locations for the occurrence of the new species, which could be the target of future botanical surveys. Croton buiquensis is here provisionally considered as Critically Endangered based on…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
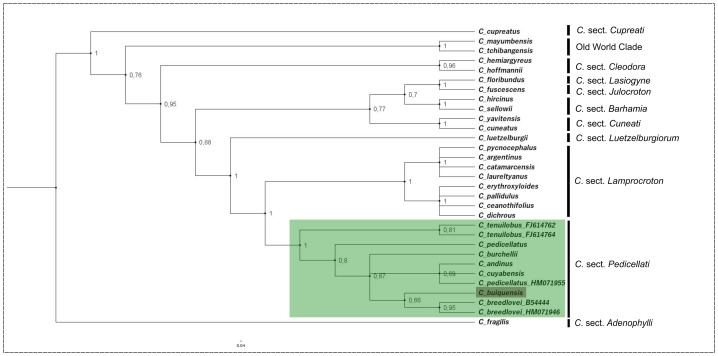 Figure 1
Figure 1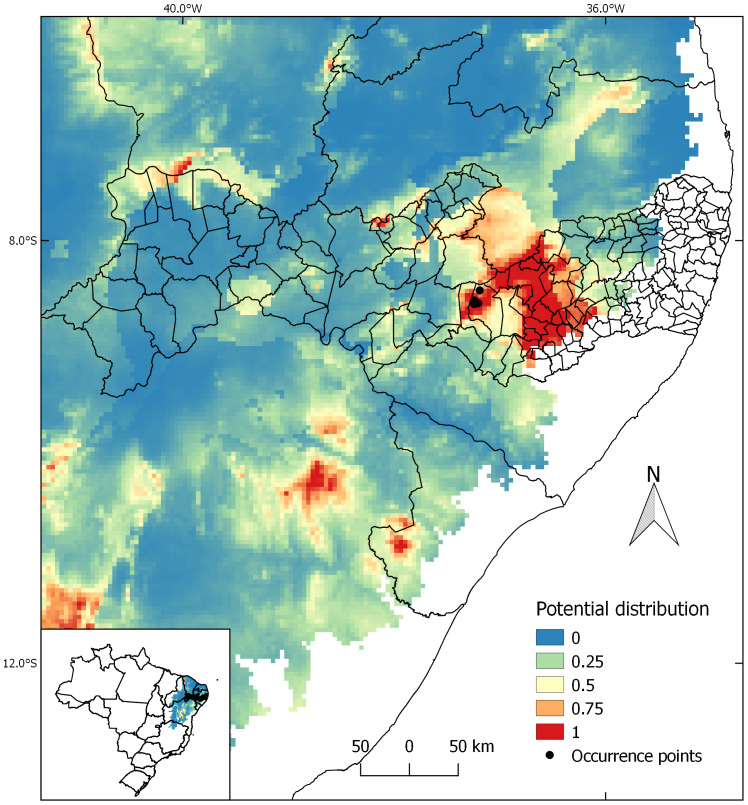 Figure 2
Figure 2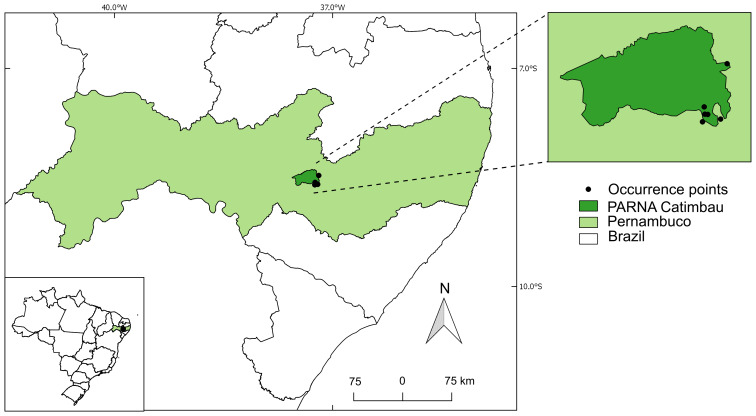 Figure 3
Figure 3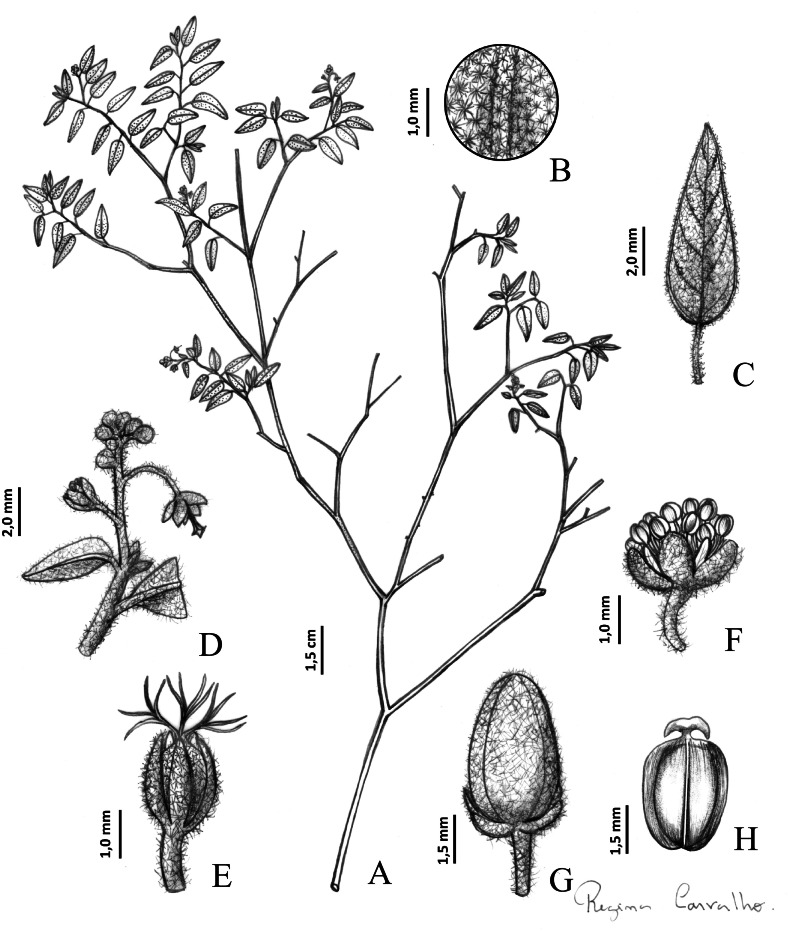 Figure 4
Figure 4 Figure 5
Figure 5- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
- —The Fundação de Amparo à Ciência e Tecnologia de Pernambuco
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Botanical Research and Applications · Plant and animal studies
Introduction
The current knowledge on the infrageneric classification of Croton L. in tropical America is synthesized in the work of Van Ee, Riina & Berry (2011). This classification placed all neotropical Croton species to section, guided by a molecular phylogenetic framework of representative taxa, and complemented by morphological and geographic evidence. Van Ee, Riina & Berry (2011) proposed 31 monophyletic sections corresponding to 31 well-supported clades. In this study, we propose and describe a new species that shows morphological affinities with C. sect. Pedicellati. This section comprises twenty species distributed disjunctly between Mexico and South America, with the majority of them occurring in central-western Brazil (11 spp.) (Van Ee & Berry, 2011; Van Ee, Riina & Berry, 2011). Croton sect. Pedicellati is currently described as monoecious, with alternate, petiolate, ovate-lanceolate, eglandular leaves, the presence of stellate to lepidote trichomes, short terminal inflorescences, pedicellate pistillate flowers with recurved pedicels, and bifid styles (Van Ee & Berry, 2011; Van Ee, Riina & Berry, 2011). All species in the section are found in scrubby tropical deciduous forests (Van Ee, Riina & Berry, 2011), which are patchy distributed across the neotropics, including those in dry inter-Andean valleys (DRYFLOR et al., 2016).
Most species of C. sect*. Pedicellati* occurring in Brazil are endemic to this country (Van Ee & Berry, 2011; Caruzo et al., 2020). Among them, six are endemic to the Cerrado (Brazilian savanna): Croton angustifrons Müll.Arg., C. burchellii Müll.Arg., C. corchoropsis Baill., C. eriocladoides Müll.Arg., C. eriocladus Müll.Arg., and C. horminum Baill.; two are endemic to the Caatinga: C. catinganus Müll.Arg. and C. tridentatus Mart. ex Müll.Arg.; and two to the Atlantic Forest: C. dracunculoides Baill. and C. tenuicaulis B.W.van Ee & P.E.Berry. Some species have very restricted distributions: C. angustifrons, recorded only in the state of Minas Gerais; C. eriocladoides, restricted to Goiás; and C. tenuicaulis, known exclusively from the state of Rio de Janeiro (Van Ee & Berry, 2011; Caruzo et al., 2020). These biogeographic patterns highlight the high degree of endemism of the section in Brazil and its importance in conservation.
As part of systematic studies on C. sect*. Pedicellati*, we encountered several specimens that morphologically did not correspond to any known Croton species. Croton buiquensis sp. nov. is only known from the state of Pernambuco, in the municipality of Buíque, with the first record dating back to 1970 (Xavier-Filho & Alves, s.n.). Like C. buiquensis, numerous new species with restricted distributions have been described in Croton with only one or few collection records (e.g., Pereira, Riina & Caruzo, 2017; Farias, Medeiros & Riina, 2019; Sodré & Silva, 2022; Sodré et al., 2023; Tagane, Souvannakhoummane & Souldeth, 2022).
Here we describe C. buiquensis as a new species of C. sect. Pedicellati using evidence from molecular phylogenetics, morphology, ecology, and geographic distribution. The new species represents one of the three Caatinga-endemic species in its section, along with C. catinganus and C. tridentatus (Van Ee & Berry, 2011); and it is the first species of this group or clade endemic to the state of Pernambuco. We also estimate the species’ potential distribution using a niche modeling approach and provide a preliminary conservation status assessment based on all the data available.
Materials and Methods
Data collection and species description
The description of C. buiquensis was based on the analyses of collections from the herbaria (acronyms according to (Thiers, 2024), continuously updated): ALCB, BHCB, CEPEC, HUEFS, IPA, PEUFR, RB, SPF, UB and UFP and HST (not indexed). We followed the general terminology for shapes and types of indumentum of Radford, Dickison & Massey (1974). For trichomes terminology, we followed the classification proposed by Pinto-Silva et al. (2023).
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Code of Nomenclature for algae, fungi, and plants (ICN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. In addition, the new name contained in this work which has been issued with identifiers by IPNI will eventually be made available to the Global Names Index. The IPNI LSIDs can be resolved and the associated information viewed through any standard web browser by appending the LSID contained in this publication to the prefix “http://ipni.org/”. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central SCIE, and CLOCKSS.
DNA extraction, amplification, and sequencing
The DNA used in the phylogenetic analyses was extracted from dried leaf tissue of herbarium specimens using the CTAB method (Doyle, 1991). We extracted DNA from the following specimens of the putative new species: Laurenio A, 2013 (PEUFR); Laurenio A, 376 (PEUFR); Sales M. F., 1060 (PEUFR); Carneiro-Torres D., 954 (HUEFS). After quantification (Qubit™ dsDNA BR Standard (Invitrogen)), the DNA samples were diluted (1:20) to perform PCR reactions to obtain the nuclear ITS genetic region. PCR amplifications were carried out in 25 µL reactions (12 µL MyTaq Red Mix (Bioline), 10 µL H2O, 1 µL of each primer (forward and reverse), and 1 µL DNA). The amplification primers were those used in recent studies in Croton and other Euphorbiaceae lineages (Silva et al., 2020; Külkamp et al., 2023). PCR products were purified with ExoSap PCR Purification and sent for sequencing at MACROGEN (Macrogen, Madrid, Spain) using the same PCR amplification primers. The nucleotide sequences were assembled and edited in UGENE 33.0 (Okonechnikov et al., 2012) and automatically aligned using the online version of MAFFT (Katoh, Rozewicki & Yamada, 2019) with default parameters. Manual adjustments were made to the alignment matrix in UGENE 33.0, applying the similarity criterion (Simmons, 2004).
Phylogenetic analyses
The newly generated ITS sequence of C. buiquensis, along with a new sequence of C. breedlovei (another species in C. sect. Pedicellati from Mexico), were aligned together with a selection of publicly available ITS sequences of Croton. This selection included all sequenced species of C. sect. Pedicellati and its sister clade C. sect. Lamprocroton (Van Ee & Berry, 2011), and representatives of Croton sections: Barhamia, Cleodora, Cuneati, Cupreati, Julocroton, Lasiogyne, Luetzelburgiorum, and, as outgroup, Adenophylli, obtained from GenBank (https://www.ncbi.nlm.nih.gov/genbank/). In total, our taxon sampling included 31 ITS sequences representing 29 Croton species. Ten of these sequences belonged to C. sect. pedicellati, representing eight species, including the new taxon. A complete list of species, their systematic positioning, vouchers, and deposition herbaria is presented in File S1. Outgroups were selected based on previous molecular studies (e.g., Van Ee & Berry, 2011; Silva et al., 2020; Masa-Iranzo et al., 2021). The ITS matrix was trimmed at both ends to minimize the effects of missing data due to unequal sequence sizes. The aligned matriz used in the analysis is provided along with the two newly generated sequences (File S2). Bayesian Inference were performed in MrBayes ver. 3.2.7 (Ronquist et al., 2012), applying the GTR+I+G model following Masa-Iranzo et al. (2021). Posterior probability (PP) values were obtained from two Markov Chain Monte Carlo (MCMC) analyses, each consisting of four chains for 10 million generations, with sampling every 1,000th generation. Tracer v1.7.2 (Rambaut, 2018) was used to check the convergence through Effective Sample Size value (ESS > 200). A 50% majority rule consensus tree was built after discarding the first 25% of the samples as burn-in. Phylogenetic analyses were hosted on the CIPRES Science Gateway (Miller, Pfeiffer & Schwartz, 2010). The software FigTree ver. 1.4.4 (Rambaut, 2018) was used to visualize and edit the phylogenetic tree. In addition to the Bayesian inference, we conducted a Maximum Likelihood analysis in IQ-TREE version 1.6.12 (Lam-Tung et al., 2015), using the same model above. Differences in genetic distance (Jukes & Cantor, 1969) between C. buiquensis and the other species of C. sect. Pedicellati were estimated in PAUP* v 4.0a (Swofford, 2002).
Occurrence data and preliminary assessment of the species conservation status
Occurrence data for the species were compiled from information present in herbarium specimens, and distribution maps were generated using QGIS 3.8 software (https://qgis.org/). A preliminary assessment of the species conservation status was conducted based on IUCN Criteria B1 and B2 (IUCN & Petitions Committee, 2024), supplemented by the calculation of Extent of Occurrence (EOO) and Area of Occupancy (AOO), using the GeoCAT tool (Bachman et al., 2011).
Ecological niche modeling
The occurrence data used for distribution map and niche modeling analysis (File S3) along with the Caatinga shapes (File S4) are provided as supplementary files. A set of 19 bioclimatic variables was obtained from the WorldClim version 1.4 database (Fick & Hijmans, 2017), with a spatial resolution of 30 s (using the limits of the Caatinga region of Brazil). To avoid multicollinearity issues and improve model robustness, a 70% correlation threshold was applied in the selection of environmental variables (Booth et al., 2014). After this analysis, seven variables were selected as predictors (Table S1).
The species’ potential distribution was modeled using the modleR package (Sánchez-Tapia et al., 2020). Cross-validation techniques were employed to evaluate model accuracy and robustness. The Maxent algorithm was used to estimate the potential distribution; this algorithm applies the maximum entropy method to model species distribution using presence data and environmental variables (Phillips et al., 2017). The use of Maxent is particularly suitable for species with few occurrence records, with five to 10 unique occurrence points (not spatially duplicated) being the minimum required to run the analyses (Phillips, Anderson & Schapire, 2006). It performs well with small sample sizes by relying on presence-only data and incorporating regularization to prevent overfitting (Wisz et al., 2008; Phillips, Anderson & Schapire, 2006; Hernandez et al., 2006). The model’s accuracy was assessed using the area under the ROC curve (AUC). A calibration area for each species was defined by creating a minimum convex polygon around all occurrences based on 100% of occurrence points, surrounded by a 1.5° buffer. The projection area was defined by the limits of the Caatinga region of Brazil, as it corresponds to the biome where the species occurs.
The environmental suitability area was calculated from the distribution models using the “cellStats”() function from the raster package (Hijmans, 2024) in R software (v. 4.4.0, R Core Team, 2023). A suitability threshold of 0.6 was applied to generate binary maps, considering cells with values above the threshold as suitable. From the suitability area indicated in the ecological niche modeling, the extent of the territory (km^2^) protected was calculated using the mask_thr_projs_mscn_b function from the ENMwizard package (Heming, Dambros & Gutiérrez, 2019). Data on protected areas were obtained from UNEP-WCMC and IUCN databases (UNEP-WCMC & IUCN , 2023). All analyses were conducted in R software (v. 4.4.0, R Core Team, 2024).
Results
Taxonomy and morphological affinities
Our specimens survey showed that C. buiquensis had often been misidentified in herbarium collections as C. pedicellatus due to some morphological similarities like its habit and leaf blade form. The morphological analysis of the available specimens indicated that the new species can be distinguished from C. pedicellatus by the indumentum of the leaf blade, the shape of the staminate petal and the styles. Overall, C. buiquensis exhibits morphological traits that are diagnostic for Croton sect. Pedicellati, such as herbaceous to shrubby habit, entire leaves, acropetiolar nectary glands absence, indument with stellate to sublepidote trichomes, terminal inflorescence. However, it can be distinguished from the rest of the species in the section by its multifid styles (vs. bifid styles).
Phylogenetic placement and relationships
Specimens of C. buiquensis were extremely difficult for PCR amplification and sequencing, even when doing the amplification of the ITS region with internal primers (i.e., sequencing it in two parts). Of the five specimens extracted for DNA only one (Carneiro-Torres 954; File S1) yielded a complete ITS sequence after several attempts using internal primers.
The results of the phylogenetic analyses conducted using Bayesian inference and maximum likelihood were practically identical in terms of topology and clade support. The Bayesian tree (Fig. 1) is used to describe the phylogenetic structure and relevant relationships. The representative species from the Croton sections sampled for our analysis formed monophyletic groups with high support value (PP = 0.95–1; Fig. 1). The species of C. sect*. Pedicellati* were also recovered in a clade (PP = 1), and sister to the C. sect. Lamprocroton clade (PP = 1).Croton buiquensis was recovered within the C. sect. Pedicellati clade, forming a moderately supported subclade together with two accessions of C. breedlovei (PP = 0.66; Fig. 1). The genetic distance of C. buiquensis from other species from C. sect. Pedicellati were low, especially when compared with the two accessions of C. breedlovei (represented by two different DNA isolates from the same voucher), its most closely related species given the available taxa with sequences (distance values 0.00982; Fig. S1).
Croton buiquensis sp. nov. in a phylogenetic context.Bayesian phylogram based on ITS sequences of the selected Croton species including C. buiquensis and representatives of C. section Pedicellati and other sections in the genus. Numbers at nodes indicate Bayesian posterior probability values (PP).
Niche modeling
Seven georeferenced points, not spatially duplicated, of C. buiquensis were confirmed and used to produce the models. The AUC value for niche modeling analysis was satisfactory (= 0.98). The generated models identified potential distribution areas of the species in a considerable portion of the “Agreste Pernambucano” mesoregion, mainly in the southern part of the state (Fig. 2), where the municipality of Buíque is located. The “Agreste” region represents a transitional zone between the “Zona da Mata”, characterized by a humid climate, and the “Sertão”, where the climate is semi-arid. It exhibits intermediate environmental conditions, with predominance of caatinga vegetation interspersed with patches of more humid vegetation at higher elevations, known as montane forests or “brejos de altitude.” The models also indicated other areas in the Northeastern region of Brazil, in the states of Bahia and Sergipe, with high potential for the species’ occurrence (Fig. 2). The occurrence points of C. buiquensis plotted on the map were located in the areas of highest probability of occurrence. The total area of suitability indicated by the models was 826,533.7 km^2^, of which only 43,995.86 km^2^ are protected by some conservation unit.
Niche modeling of Croton buiquensis sp. nov.Blue color represents regions with low climatic suitability and orange and red colors represent regions with high climatic suitability. The colored background represents the area of the Caatinga domain (see Methods).
Distribution and preliminary conservation status assessment
Croton buiquensis has been only recorded in the municipality of Buíque, in the state of Pernambuco, northeastern Brazil, within the Catimbau National Park (a conservation unit with full protection, according to Brazilian legislation) (Fig. 3). Considering the extent of occurrence (EOO) of 87.682 km^2^ and its area of occupancy (AOO) of 8 km^2^, the species is classified as Critically Endangered (CR). Information about the health of the populations, including the number of mature individuals, is currently lacking.
Geographic distribution map of Croton buiquensis sp. nov.
Taxonomic treatment
Croton buiquensis J.C.P. Oliveira sp. nov.—TYPE: Brazil, Pernambuco, Buíque, Parque Nacional da Serra do Catimbau, subida para Serra das Torres; 8°33′60″S, 37°14′28″W; 28 Jun 2007, fl., Carneiro-Torres D. 954; holotype: HUEFS (HUEFS000106140!), isotypes: HUESB-JQ (HUESB2980), SP (SP0396823!) (Fig. 4).
Main morphological features distinguishing Croton buiquensis sp. nov.Croton buiquensis sp. nov. (A) Flowering branch. (B) Indumentum on the abaxial side of the leaf blade. (C) Detail of a leaf. (D) Detail of an inflorescence. (E) Pistillate flower. (F) Staminate flower. (G) Capsule. (H) Seed. A from Laurênio A. 123 (PEUFR); (B, C) and (D) from Cano O. 791 (IPA); (E, F, G) and (H) from Fontana AP 10096 (PEUFR).
Etymology
The specific epithet of the new species refers to the municipality of Buíque in the state of Pernambuco where all specimens of C. buiquensis have been recorded to date. Buíque is a word of Tupi origin, the main indigenous language spoken in Brazil until the mid-18th century, and it means place of snakes.
Diagnosis
Besides its geographic range, C. buiquensis can be distinguished from other species in C. sect. Pedicellati by its 4-fid to multifid styles, the presence of sublepidote trichomes on branches and leaf blades, and the crossbow-shape caruncle.
Description
Herbs to subshrubs, monoecious, approximately 50 cm tall. Branches ferruginous to grayish, indumentum hispid to glabrous, sometimes tomentose, with stellate to sublepidote trichomes; stipules 0.8–1.0 mm long, ovate, hispid indumentum, stellate trichomes. Petiole 0.4–0.7 cm long, hispid, stellate to porrect-stellate trichomes (porrect ray measuring 60–100 µm), eglandular. Leaves alternate, chartaceous, leaf blades ovate to ovate-lanceolate, 1.1–1.6 × 0.4–0.7 cm, venation eucamptodromous, apex acute, base rounded, adaxial surface hispid, with densely grouped stellate trichomes, abaxial surface hispid, stellate trichomes, sometimes ferruginous, margin entire. Inflorescence a thyrse, 0.4–1.7 cm long, bracts ovate, approximately 1.0 mm long, hispid to pubescent, stellate trichomes, pistillate flowers located at the base of the axis and staminate flowers on the distal portion, sometimes flowers are very close to one another due to the short length of the inflorescence axis. Staminate flowers ca. 1.4 × 1.7 mm, pedicel 2.0–2.1 mm long, hispid to pubescent, stellate trichomes; sepals 5, ca. 1.0 × 1.1 mm, ovate to elliptical, hispid to pubescent, stellate trichomes; petals 5, ca. 0.3 × 1.0–1.1 mm, ovate to ovate-lanceolate, glabrous, margin entire, sometimes with long simple trichomes, appearing ciliate; stamens 10, 1.1–1.2 mm long. Pistillate flowers ca. 2.0 × 2.8 mm, pedicels 0.3–8.0 mm long; sepals 5, ovate, 2.0–2. 1 × 0.7 mm, free from one another, apex acute, margin entire, hispid to pubescent indumentum, stellate trichomes; styles 4-fid to multifid, glabrous to pubescent, stellate, porrect trichomes; petals absent; ovary globose, approximately 1.1 mm in diameter, hispid to pubescent, with stellate trichomes. Capsules ellipsoid, ca. 0.3 × 0.5 cm, hispid indumentum, stellate, porrect trichomes; fruiting pedicel 4.0–7.0 mm long; columella 2.0–3.0 mm long. Seeds rounded, slightly rough, approximately 3.0–4.0 × 3.0 mm, caruncle crossbow-shape.
Paratypes
BRAZIL. Pernambuco: Buíque, Parque Nacional do Catimbau, Chapada São José, 8°37′23″S, 37°09′21″W, 14.IX.2011, fl., Costa A. C. G. et al. 46 (IPA); Chapada de São José; 8°37′23″S 37°09′21″W, 03.IV.2000, Sales M. 1060 (JPB, PEUFR, UFP); Estrada Buíque Catimbau, 37°09′15″W, Laurênio A. 123 (K, NY, PEUFR); Parna Catimbau; 8°35′37″S 37°12′18″W, 01.VII.2015, Felix L.P. 15663 (CSTR, EAN); Parque Nacional do Catimbau, Serrinha, 8°37′23″S 37°09′21″W, 13.I.2011, Costa A. C. G. et al. 12 (IPA); Sobrado, Serra das Andorinhas, 8°28′17″S 37°11′26″W, 30.VIII.2018, Fontana A. P. 10096 (HUEFS); Serra do Catimbau, 8°37′23″S 37°09′21″W, 15.X.1970, Xavier-Filho L. & Alves J. L. H. s.n. (UFP); Vale do Catimbau, 8°37′23″S 37°09′21″W, Costa Filho L. O. s.n. (UFP); Vale do Catimbau, Paraiso Tropical, alto do lajeiro, 8°34′58″S 37°14′23″W, 18.VII.2007, Cano O. et al. 791 (IPA, HUEFS); Vale do Catimbau, 8°37′23″S, 37°09′21″W, fl., Gomes A. P. S. et al. 509 (IPA); Vale do Catimbau, Serra das Torres, Savana Estépica/caatinga, 8S 35′58″, 37°14′41″W, 866 m.a.s.l., 29.XI.2016, fl., Oliveira M. 6509 (HUEFS, PISF, RB).
Habitat and ecology
Croton buiquensis is only known from the municipality of Buíque, in the state of Pernambuco, northeastern Brazil, within the Catimbau National Park. It grows in hilly areas of steppe savanna with dense shrub vegetation and/or shrub-arboreal vegetation, as well as on rocky outcrops, at elevations ranging from 780 to 900 m. The collected specimens were found flowering between April and October.
Notes
The crossbow-shape caruncle of C. buiquensis appears to be unique within Croton. It is characterized by an arched and elongated structure, resembling a crossbow, with lateral projections and a short, central stalk attached to the apical part of the seed (Figs. 4H and 5E). The crossbow-shape differs from the known caruncle shapes present in most Croton species (e.g., reniform, trapezoidal, oblong, elliptic, winged) (e.g., Riina et al., 2010; Secco et al., 2012; Anton et al., 2024; Pereira et al., 2025).
Herbarium images of Croton buiquensis sp. nov.Croton buiquensis sp. nov. (A) Image of the type at HUEFS. (B) Inflorescence, showing pistillate flowers and staminate cymules. (C) Pistillate flower at anthesis showing the pedicel 4-multifid styles and oval to elliptical sepals. (D) Opened capsule stage shown from the side. (E) Seed shown from the front side, and the crossbow shaped caruncle; abaxial leaf surface on the left showing the ferrugineous trichomes. (A) from Carneiro-Torres 954 (HUEFS); (B) and (E) from Cano O. 791; (C) and (D) Fontana AP 10096 (PEUFR).
Discussion
Overall, the topology of our phylogenetic analysis at the genus level (backbone) and within section Pedicelati is congruent with previous ITS phylogenetic analyses of Croton (Van Ee & Berry, 2011; Van Ee, Riina & Berry, 2011; Arévalo et al., 2017). The membership of C. buiquensis to C. sect. Pedicellati is supported by morphological traits shared with other species in this clade, including herbaceous to shrubby habit, entire leaves, absence of acropetiolar nectary glands, stellate trichomes on both leaf sides and ovate pistillate sepals (Van Ee & Berry, 2011). However, as indicated in the diagnosis above, there are three key morphological characters differentiating C. buiquensis from the other species in the section (Table 1). The new species represents the first record of 4-fid and multifid styles in C. sect. Pedicellati, thereby justifying an adjustment to the description of the section to reflect the current variation in style branching: bifid, 4-fid or multifid styles. Tetrafid (multifid) styles are also present in C. sect*. Lamprocroton*, the sister section to C. sect*. Pedicellati* and other sections of Croton (Lima de & Pirani, 2008; Van Ee & Berry, 2011).
Table 1: Main morphological differences between Croton buiquensis sp. nov. and some of the most similar species of the C. sect. Pedicellati.
In general Croton species are relatively easy to sequence for the ITS region as demonstrated for the numerous phylogenetic publications using this genetic marker (e.g., Berry et al., 2005; Van Ee, Riina & Berry, 2011; Rossine et al., 2025). However, C. buiquensis appears to be an exception. It took us many tries in the molecular lab using material from five different herbarium specimens until we finally obtained one ITS sequence for the phylogenetic analysis. It is possible that the chemistry of this species includes one or more compounds that interfere with the PCR amplification process. Croton and Euphorbiaceae in general are well known for their highly diverse chemistry (Salatino, Salatino & Negri, 2007; Lopes et al., 2012; Coelho et al., 2016; Xu, Liu & Liang, 2018; Guerra-Júnior et al., 2022), including numerous secondary metabolites, so it is not surprising that PCR inhibitors could be present in some species.
Specimens of C. buiquensis had been misidentified in herbaria as C. pedicellatus this confusion is probably due to the high similarity in some morphological features between the two species. However, the new species can be distinguished from C. pedicellatus by the indumentum of the leaf blade (hispid vs. hirsute-pubescent to pubescent in C. pedicellatus), the shape of the staminate petal (ovate vs. oblong), and the style (4-fid to multifid vs. bifid). The distinction of the type of leaf blade indumentum is due to differences in the length of the central (porrect) ray of the trichomes.
The phylogenetic analysis (Fig. 1) as well as the Jukes-Cantor analysis (Fig. S1), shows that C. buiquensis is genetically close to C. breedlovei, a species endemic to the Chiapas region in Mexico (Van Ee & Berry, 2011). Despite the phylogenetic proximity, the two species can be distinguished by their geographical distribution as well as by several morphological characters (see Table 1). Disjunct distribution patterns have also been reported in other Croton lineages, which show fragmented geographic ranges across Neotropics (e.g., Riina et al., 2018; Riina et al., 2021). This fragmented geographic pattern is also shown below the species level with species showing disjunct populations at continental or regional scales (e.g., Caruzo et al., 2016; Rossine et al., 2025).
Considering that C. buiquensis occurs within a permanent preservation area (Catimbau National Park), one might assume that the species is not as exposed to threats. However, areas with rupestrian field vegetation are quite sensitive to climate change, and recent future projections for these environments show considerable reduction in size, even in protected areas (Fernandes et al., 2018). Despite being a permanent conservation unit, Catimbau National Park is not immune to environmental degradation processes due to anthropogenic actions such as ecotourism and housing (Vranckx & Castilho, 2010; Silva & Maia, 2011; Azevedo & Bernard, 2015). Protected areas are also impacted by the establishment of invasive exotic species (Silva & Fabricante, 2019), which subject endemic native species to a high degree of threat. For these reasons, C. buiquensis is here considered as critically endangered pending formal evaluation by the IUCN.
The results of the ecological niche modeling point to other areas in the “Agreste Pernambucano” region as likely locations for the species’ occurrence. These areas have similar climatic conditions associated with sub-humid and semi-arid climates, over pediplains or mountain ranges (Albuquerque et al., 2019). However, no records of this species have been found in these areas up until now. Thus, the species is for now considered restricted to the municipality of Buíque, being the first species of C. sect. Pedicellati endemic to the state of Pernambuco, but we believe that further collection efforts in these areas are needed. The models also identified other areas in the states of Bahia and Sergipe as having high potential for the species’ occurrence. However, models can overestimate species distribution, as the environmental conditions predicted for an ecological niche may be present in several locations within a relatively large geographic area (Urbina & Loyola, 2008; Jetz, Sekercioglu & Watson, 2008; Suarez-Contento et al., 2024). The existence of potential areas outside the current distribution of the species does not guarantee that it can effectively colonize these regions. Factors such as limitation in dispersal capacity, species competition or biological barriers can prevent geographic range expansion (Mendes et al., 2020; Velazco et al., 2020).
Conclusions
The description of C. buiquensis as a new endemic species from Northeast Brazil underscores the importance of continuous efforts in documenting the flora in highly diverse and often underexplored regions of the Neotropics. This study not only positions the species within C. sect*. Pedicellati* but also provides essential information about its ecology and potential distribution based on niche models. The new species also expands the morphological concept of C. sect. Pedicellati, which should now include bifid and multifid styles as well as sublepidote trichomes. In addition, this is the first time that a crossbow-shape caruncle is described in Croton. The restricted geographic range of C. buiquensis, combined with anthropogenic pressure on ecosystems in the Brazilian semi-arid region, including legally established conservation areas, reinforces its threatened status and the need for additional conservation strategies.
Supplemental Information
10.7717/peerj.20718/supp-1Supplemental Information 1Supplementary Files
10.7717/peerj.20718/supp-2Supplemental Information 2Sequences of ITS (fasta format) used in the studyPreviously published and newly generated sequences.
10.7717/peerj.20718/supp-3Supplemental Information 3Occurrence points (coordinates) of Croton buiquensis sp. nov
10.7717/peerj.20718/supp-4Supplemental Information 4Shapefiles of the Brazilian Caatinga region used in the niche modeling analysis of Croton buiquensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Albuquerque LS Nóbrega RS Albuquerque LS Moreira WAB Menezes RSC 2019 The relationship between relief and climate as proposition of characterization of landscape physiology in Pernambuco, Brazil BRAZIL Geo UERJ 34e 40942(In Portuguese)10.12957/geouerj.2019.40942 · doi ↗
- 2Anton AM Belgrano MJ Berry PE Cardiel JM Carrión RJF Chiarini FE Silva OLM González A Kulkamp J Riina R 2024 Zuloaga FO Zanotti CA Moroni PD Dicotiledoneae: Euphorbiaceae Vol. 10Flora vascular de la República Argentina Buenos Aires IBODA, Consejo Nacional De Investigaciones Científicas y Técnicas (CONICET)
- 3Arévalo R Van Ee BW Riina R Berry PE Wiedenhoeft AC 2017 Force of habit: shrubs, trees and contingent evolution of wood anatomical diversity using Croton (Euphorbiaceae) as a model system Annals of Botany 119456357910.1093/aob/mcw 24328065919 PMC 5458714 · doi ↗ · pubmed ↗
- 4Azevedo IS Bernard E 2015 Avaliação do nível de relevância e estado de conservação da caverna Meu Rei no PARNA Catimbau, Pernambuco Revista Brasileira de Espeleologia 15123
- 5Bachman SJ Moat AW Hil JT Scott B 2011 Supporting Red List threat assessments with Geo CAT: geospatial conservation assessment tool Zoo Keys 15011712610.3897/zookeys.150.2109 PMC 323443422207809 · doi ↗ · pubmed ↗
- 6Berry PE Hipp AL Wurdack KJ Van Ee B Riina R 2005 Molecular phylogenetics of the giant genus Croton and tribe Crotoneae (Euphorbiaceae sensu stricto) using ITS and trn L-trn F DNA sequence data American Journal of Botany 921520153410.3732/ajb.92.9.152021646170 · doi ↗ · pubmed ↗
- 7Booth TH Nix HA Busby JR Hutchinson MF 2014 BIOCLIM: the first species distribution modelling package, its early applications and relevance to most current MAXENT studies Diversity and Distributions 2011910.1111/ddi.12144 · doi ↗
- 8Caruzo MBR Santos RF Feio AC Meira RMSA Riina R 2016 Redefinition of Croton macrobothrys (Euphorbiaceae), a tree species from the Brazilian Atlantic Forest, with the description of a new subspecies Phytotaxa 27315110.11646/phytotaxa.273.1.5 · doi ↗