Conformational analysis of difluoromethylornithine: factors influencing its gas-phase and bioactive conformations
Matheus P Freitas

TL;DR
This paper studies how fluorine affects the shape of DFMO, a drug used to treat African trypanosomiasis, and explains why certain molecular conformations are more stable.
Contribution
The study reveals that the fluorine gauche effect in DFMO is driven by hyperconjugative stabilization and is modulated by steric and electrostatic effects.
Findings
Type-I conformers dominate due to the fluorine gauche effect and hyperconjugative stabilization.
Steric effects influence the relative stability of low-energy conformers.
The bioconformation of DFMO in crystallographic data is a type-II structure due to hydrogen bonding.
Abstract
Difluoromethylornithine (DFMO, eflornithine) is a fluorinated analogue of ornithine that serves both as an inhibitor of ornithine decarboxylase and as a therapeutic agent against African trypanosomiasis. Beyond its pharmacological importance, DFMO provides a valuable model for examining how fluorine substitution governs molecular conformation. A comprehensive quantum-chemical study was performed to elucidate the origins of DFMO’s conformational stability. High-level DLPNO-CCSD(T)/CBS calculations revealed that type-I conformers – those maximizing gauche interactions between C–F and C–N bonds – dominate the equilibrium population, confirming the presence of the fluorine gauche effect. natural bond orbital (NBO) analysis showed that this preference arises primarily from hyperconjugative stabilization, particularly the σCH → σ*CN interaction, while steric effects modulate the relative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
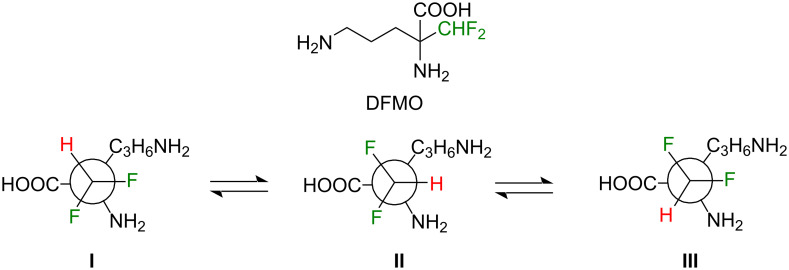 Figure 1
Figure 1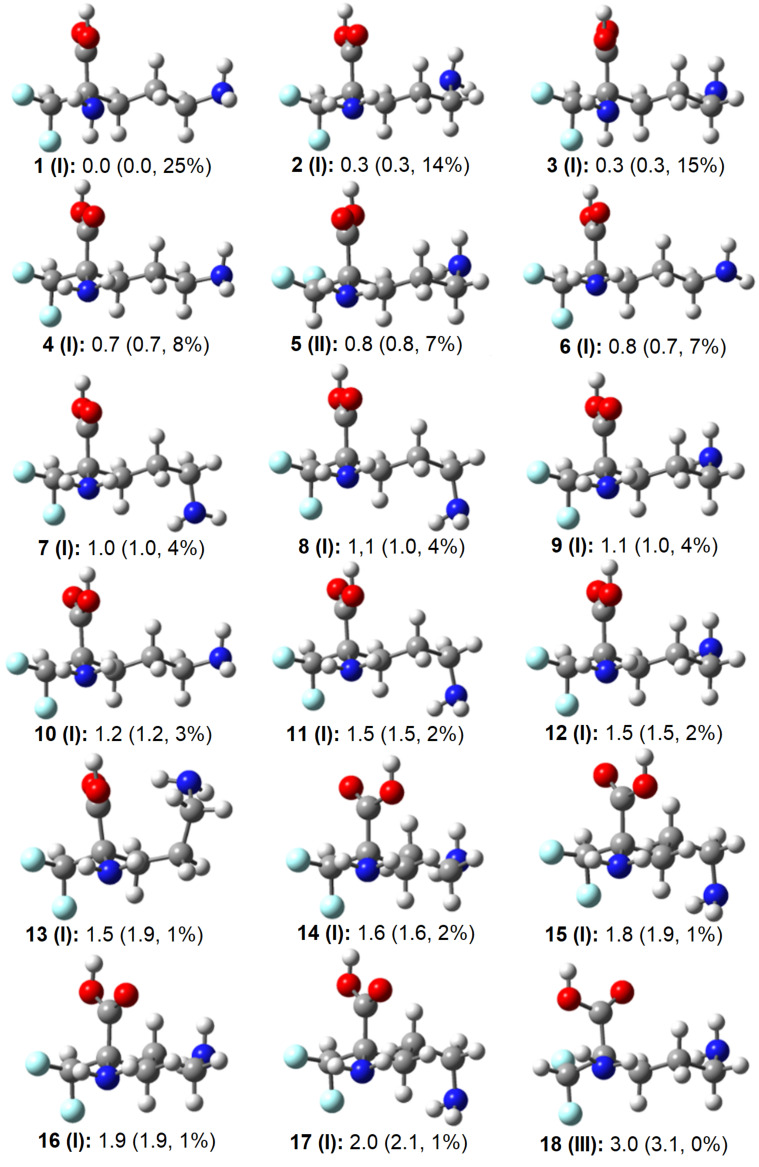 Figure 2
Figure 2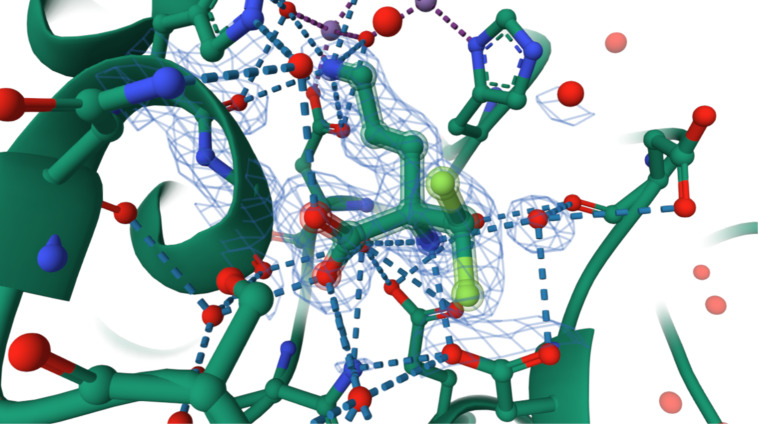 Figure 3
Figure 3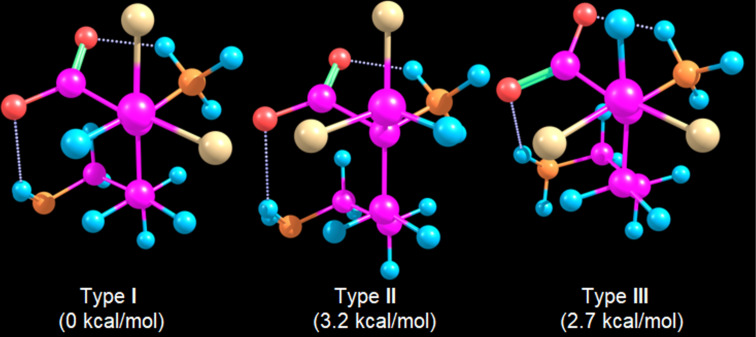 Figure 4
Figure 4| Conformer |
|
|
|
|
|
| ||||
|
| 0.0 | −502.6 | 502.6 | 295.4 |
|
| 0.5 | −496.2 | 496.7 | 296.3 |
|
| 0.5 | −504.2 | 504.7 | 296.3 |
|
| 0.7 | −499.5 | 500.2 | 293.2 |
|
| 1.2 | −501.7 | 502.8 | 294.0 |
|
| 0.8 | −497.5 | 498.3 | 294.3 |
|
| 1.4 | −498.4 | 499.8 | 295.0 |
|
| 1.2 | −500.4 | 501.6 | 294.0 |
|
| 1.3 | −500.6 | 501.9 | 294.0 |
|
| 1.3 | −502.9 | 504.2 | 293.8 |
|
| 1.7 | −503.8 | 505.5 | 294.8 |
|
| 1.7 | −504.5 | 506.2 | 294.9 |
|
| 3.2 | −511.4 | 514.5 | 298.5 |
|
| 1.8 | −498.0 | 499.8 | 294.1 |
|
| 2.2 | −498.8 | 500.9 | 295.1 |
|
| 1.9 | −497.5 | 499.4 | 293.9 |
|
| 2.2 | −498.5 | 500.8 | 295.1 |
|
| 3.2 | −498.5 | 501.7 | 296.3 |
- —Conselho Nacional de Desenvolvimento Científico e Tecnológicohttps://doi.org/10.13039/501100003593
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superiorhttps://doi.org/10.13039/501100002322
- —Fundação de Amparo à Pesquisa do Estado de Minas Geraishttps://doi.org/10.13039/501100004901
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFluorine in Organic Chemistry · Polyamine Metabolism and Applications · Trypanosoma species research and implications
Introduction
Difluoromethylornithine (DFMO, in its racemic form, also known as eflornithine – Figure 1) is a fluorinated analogue of ornithine that has attracted considerable attention due to its dual relevance in medicine and structural chemistry. Clinically, DFMO has been employed in the treatment of African trypanosomiasis (sleeping sickness), particularly the second stage of the disease caused by Trypanosoma brucei gambiense [1–3]. It is also used as a topical agent to reduce excessive facial hair growth in women [4–5]. Beyond these applications, DFMO is recognized as a potent inhibitor of ornithine decarboxylase, the key enzyme that catalyzes the first step in polyamine biosynthesis [6]. This inhibition underlies both its therapeutic utility and its importance as a biochemical probe.
Structure of racemic difluoromethylornithine (DFMO) and conformers I–III of the (S)-enantiomer ((S)-DFMO) obtained from rotation around the difluoromethyl group.
From a structural standpoint, DFMO provides an intriguing case study for exploring conformational effects associated with the difluoromethyl (–CHF₂) motif. The presence of vicinal fluorine substituents is known to strongly influence conformational equilibria through stereoelectronic effects, notably the fluorine gauche effect [7]. This effect arises from hyperconjugative interactions (typically σ_CH_ → σ*CF) that stabilize conformations in which the C–F bonds adopt a gauche orientation relative to polar bonds, such as C–N or C–O [8–10]. In DFMO, the difluoromethyl group is directly connected to the ornithine backbone, and its conformation is expected to be shaped not only by intrinsic stereoelectronic preferences but also by intermolecular interactions within the ornithine decarboxylase active site.
Therefore, the conformational behavior of DFMO arises from a delicate balance between the fluorine gauche effect and the intermolecular interactions that stabilize the ligand–enzyme complex. Elucidating this interplay is essential for rationalizing DFMO’s inhibitory potency and may also offer broader insight into how fluorinated substituents control molecular conformation in biologically relevant environments. Although the biological milieu is known to induce only minor structural changes in DFMO [11], a systematic evaluation of its zwitterionic and non-zwitterionic forms in the gas phase, aqueous solution, and biological environments is necessary to disentangle the respective contributions of intra- and intermolecular interactions to its conformation and biological activity.
In this work, we present a quantum-chemical study aimed at elucidating the origin of the conformational stability of DFMO. Particular emphasis is placed on natural bond orbital (NBO) analysis [12], which enables a detailed characterization of hyperconjugative interactions and other stereoelectronic effects that contribute to the preferred conformations of the difluoromethyl motif.
Results and Discussion
According to Wolfe [7], the gauche effect is defined as “a tendency to adopt that structure which has the maximum number of gauche interactions between adjacent electron pairs and/or polar bonds.” In this framework, DFMO is expected to display the gauche effect when type-I structures dominate, because the polar C–N bond forms two gauche interactions with the polar C–F bonds. High-level DLPNO-CCSD(T)/CBS calculations support this expectation: the four lowest-energy structures are type I and together account for about 62% of the equilibrium population. By contrast, type-II and type-III structures contribute only 7% and 0%, respectively (Figure 2). These results demonstrate that DFMO exhibits the gauche effect, unlike 1,1,2-trifluoroethane and 1,1,2,2-tetrafluoroethane, whose preferred geometries minimize gauche C–F interactions [13–18]. It is also noteworthy that the zwitterionic form of DFMO dissociates upon gas-phase optimization, whereas the non-zwitterionic form in implicit water, modeled using the solvation model density (SMD), remains stable in a type-I conformation, with conformer 4 being the most stable (14%). Consequently, the gauche effect persists in solution (Figure S1, Supporting Information File 1).
Representative conformations of (S)-DFMO with their relative electronic energies (in kcal mol−1), standard Gibbs free energies (in parenthesis), and Boltzmann populations calculated at the DLPNO-CCSD(T)/CBS level of theory. Color labels: H = white, C = grey, N = blue, O = red, F = electric blue.
To clarify the origin of this behavior, natural bond orbital (NBO) analyses were carried out to separate steric and electron-delocalization effects. The principal low-energy structures (1–4) differ mainly by rotations about the C–COOH, C–NH_2_, and C–C(NH_2_) bonds. As a result, their relative energies are governed primarily by the local environments of these groups rather than by the orientation of the difluoromethyl unit. Steric analysis based on exchange energies between natural localized molecular orbitals shows that structure 4 experiences the weakest steric repulsion, whereas structures 1–3 are among the most sterically crowded (Table 1). Several interactions contribute to this trend; notably, the nN/σ_CC_ interaction involving the NH₂ group geminal to the difluoromethyl moiety is significantly less repulsive in structure 4. In addition, the zigzag conformation of the N-alkyl chain reduces σ_CN_/σ_CC_ repulsions.
Structure 5 (type II) displays a comparatively low steric penalty (294.0 kcal mol^−1^), much smaller than those of structures 1–3. This reduction arises in part because its C–N bond is not flanked by two C–F bonds. At the other extreme, structure 13 is highly destabilized sterically due to folding of the N-alkyl chain, which brings N–H and C–N bonds into close contact with C–H, C–C, and C=O bonds. This geometry generates several n/σ and σ/σ interactions of 0.5–1.4 kcal mol^−1^ each (see Supporting Information File 1). Overall, steric effects alone cannot explain the stability ordering, as indicated by the near-zero correlation between relative energy (Erel) and steric energy (Esteric). Electron delocalization therefore plays a decisive role.
This contribution can be quantified by decomposing Erel into Lewis (EL) and non-Lewis (ENL) terms. The former includes destabilizing 4-electron/2-orbital interactions, while the latter accounts for stabilizing 2-electron/2-orbital interactions. Although EL contains steric effects, its correlation with Esteric is weak (R^2^ = 0.30). In practice, Erel and ENL are obtained directly from the NBO analysis, whereas EL is determined by difference (Table 1). Structure 13 is the most delocalized but also experiences strong destabilizing interactions. The five least stable structures (14–18), including the type-III geometry, gain less than 500 kcal mol^−1^ from delocalization, whereas the most stable structure (1) is stabilized by 502.6 kcal mol^−1^. Notably, structure 5 (type II) is also strongly stabilized by delocalization (ENL = −501.7 kcal mol^−1^), while structure 2 shows weaker stabilization (−496.2 kcal mol^−1^). These results indicate that the gauche effect in DFMO arises from a balance between strong non-Lewis stabilization and only moderate Lewis-type destabilization.
Further insight was obtained by examining specific hyperconjugative interactions through second-order perturbation analysis. In 1,2-difluoroethane, the classical gauche effect is attributed to σ_CH_ → σCF hyperconjugation [8–1019]. In DFMO, the σ_CH_ orbital of the difluoromethyl group acts as the primary donor, while the vicinal σCN orbital is the main acceptor. Additional antiperiplanar interactions within the (F₂H)C–C fragment, which together with Lewis-type effects control the orientation of the difluoromethyl group, are summarized in Table S1 (Supporting Information File 1). As expected, σ_CH_ → σCN is the dominant interaction (>3 kcal mol^−1^) and is present in all type-I structures. When these interactions are summed for each structure, 12 of the 13 lowest-energy structures show stabilization greater than 11 kcal mol^−1^. Structure 5 is the only exception, yet it remains low in energy due to its reduced steric repulsion. By contrast, the least stable structure (18) shows the weakest hyperconjugative stabilization because, as a type-III geometry, it exhibits a σ_CH_ → σCC interaction (2.4 kcal mol^−1^) instead of the more favorable σ_CH_ → σ*CN interaction.
Overall, the gauche effect in DFMO appears to be driven by hyperconjugation and modulated by steric effects. For instance, conformers 1 and 2 are similarly stabilized by hyperconjugation, but the former experiences weaker steric repulsion. Conversely, conformer 18 is both the least stabilized by hyperconjugation and one of the most sterically hindered.
The gas-phase preferences can be contrasted with the bioconformation of (S)-DFMO. Although PDB entry 9FOS is reported as ornithine decarboxylase from Leishmania infantum complexed with PLP and DFMO, the bound ligand corresponds instead to human arginase I (PDB code 3GN0). While binding to arginase I is not related to DFMO’s pharmacological activity, both enzymes recognize ornithine-related substrates, allowing DFMO to occupy the arginase active site in crystallographic studies. In this environment, the fluorine atoms do not engage in favorable contacts with residues or water molecules. Instead, the geometry is dictated by interactions involving the NH₂ and COOH groups, yielding a bioconformation similar to arrangement II. Specifically, the NH₂ group forms a hydrogen bond with ASP183, and the carboxylate interacts with ASN130 (Figure 3), with additional hydrogen bonds to water molecules. Consequently, intermolecular interactions dominate and override the intramolecular forces responsible for the gauche effect in the isolated molecule. This is consistent with the relative strength of hydrogen bonds (5–40 kcal mol^−1^) compared with typical hyperconjugative interactions [20].
(S)-DFMO bound within the active site of human arginase I obtained from the Protein Data Bank (3GN0) (https://doi.org/10.2210/pdb3gn0/pdb, [21]). Dotted lines represent hydrogen-bonding interactions with amino acid residues and water molecules, while the ligand is delineated by a contour line to distinguish it from the binding cavity.
Finally, because neutral amino acids dominate in the gas phase whereas zwitterions prevail in solution [21–24], the zwitterionic form of DFMO was examined in implicit water. A conformational search using the global optimizer algorithm at the GFN2-xTB level identified 66 structures within 3 kcal mol^−1^. All but one were type I (Figure 4). The first type-II and type-III geometries lie 3.2 and 2.7 kcal mol^−1^ above the lowest-energy structure, corresponding to negligible Boltzmann populations. Thus, zwitterion formation reinforces the gauche effect, likely through an electrostatic attraction between negatively charged fluorine atoms and the positively charged NH₃^+^ group, as observed in related systems [25]. Even so, this intramolecular stabilization remains insufficient to overcome the stronger intermolecular interactions present in biological environments.
Lowest-energy type-I, type-II, and type-III conformers of the zwitterionic form of (S)-DFMO. Color labels: H = blue, C = pink, N = orange, O = red, F = yellow.
Conclusion
DFMO exhibits the fluorine gauche effect, as type-I conformers – those maximizing gauche interactions between the C–F and C–N bonds – dominate the conformational equilibrium by a large margin. This preference arises primarily from hyperconjugative stabilization, particularly the σ_CH_ → σ*CN interaction, which is absent in type-II and type-III conformers. This behavior contrasts with that of 1,1,2-trifluoroethane and 1,1,2,2-tetrafluoroethane, which instead favor conformations that minimize gauche interactions between the polar C–F bonds. The relative energies among the type-I conformers, however, are further modulated by steric effects. Notably, the type-II conformer (5) ranks among the most stable structures owing to its minimal steric repulsion. In a biological environment, this balance shifts, since strong intermolecular interactions – especially hydrogen bonding with amino acid residues – favor a type-II geometry as the predominant bioconformation. Nevertheless, when different conformers engage in comparable intermolecular interactions, intramolecular effects such as the gauche effect are expected to play a decisive role in determining the bioactive conformation and, consequently, the molecular bioactivity.
Methods
A stochastic conformational search of (S)-DFMO was performed at the semiempirical AM1 level [26] using the Spartan program [27]. The Monte Carlo algorithm employed explores the conformational space by randomly sampling molecular geometries and accepting or rejecting new structures according to their relative energies. Among the 100 generated conformers, 20 exhibited nonzero Boltzmann populations, of which 16 were unique. These 16 conformers (arrangement I), along with two additional structures – one with the COOH group (II) and another with the C_2_H_4_NH_2_ group (III) positioned between the two C–F bonds – were subsequently optimized at the B3LYP-GD3BJ/6-311++G(d,p) level [28–31] of density functional theory. Frequency calculations confirmed that all optimized geometries correspond to true minima. These calculations were performed using the Gaussian 16 program [32]. The optimized structures were then subjected to single-point energy calculations at the DLPNO-CCSD(T)/CBS level [33–34] with the ORCA software package [35] to obtain accurate conformational energies, where DLPNO-CCSD(T) denotes the domain-based local pair natural orbital coupled-cluster method including single, double, and perturbative triple excitations, and the complete basis set (CBS) limit was obtained by extrapolating Hartree–Fock and correlation energies from calculations performed with the cc-pVTZ and cc-pVQZ basis sets. Natural bond orbital (NBO) analyses [12] were also performed at the same DFT level, employing the NBO7DEL, LEWIS, and STERIC keywords to suppress electron-delocalization effects and thereby quantify the contributions of Lewis- and non-Lewis-type interactions to the total electronic energy, as well as to evaluate steric exchange energies from pairwise interactions. Additionally, the zwitterionic structure of DFMO reported by Ilies et al. [21] was investigated through a conformational search using the global optimizer algorithm (GOAT) at the GFN2-xTB(ALPB) semiempirical level [36–37] with an implicit water solvent model, considering that the β-alanine zwitterion predominates in aqueous solution [22], while neutral forms arising from proton transfer are typically observed in the gas phase [23–24].
Supporting Information
File 1NBO outcomes, standard orientations and Gibbs free energies for the studied compounds.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Taelman H Marcelis L Sonnet J Kazyumba G Van den Enden E Wery M Schechter P J Bull Soc Pathol Exot Ses Fil 1988815785883143491 · pubmed ↗
- 2Van Nieuwenhove S Schechter P J Declercq J BonéG Burke J Sjoerdsma A Trans R Soc Trop Med Hyg 19857969269810.1016/0035-9203(85)90195-63938090 · doi ↗ · pubmed ↗
- 3Doua F Boa F Y Schechter P J Miézan T W Diai D Sanon S R De Raadt P Haegele K D Sjoerdsma A Konian K Am J Trop Med Hyg 19873752553310.4269/ajtmh.1987.37.5253120607 · doi ↗ · pubmed ↗
- 4Martin K A Chang R J Ehrmann D A Ibanez L Lobo R A Rosenfield R L Shapiro J Montori V M Swiglo B A J Clin Endocrinol Metab 2008931105112010.1210/jc.2007-243718252793 · doi ↗ · pubmed ↗
- 5Hoffmann R Eur J Dermatol 200818657010.1684/ejd.2008.031318086592 · doi ↗ · pubmed ↗
- 6Qu N Ignatenko N A Yamauchi P Stringer D E Levenson C Shannon P Perrin S Gerner E W Biochem J 200337546547010.1042/bj 2003038212859253 PMC 1223689 · doi ↗ · pubmed ↗
- 7Wolfe S Acc Chem Res 1972510211110.1021/ar 50051 a 003 · doi ↗
- 8Goodman L Gu H Pophristic V J Phys Chem A 20051091223122910.1021/jp 046290 d 16833433 · doi ↗ · pubmed ↗