Effects of dietary dried distillers’ grains with solubles and NSP enzyme supplementation on growth performance, intestinal morphology, immunity, and economic efficiency in broilers
Amr M. Shams-Eldin, Ahmed Sayed-Ahmed, Ahmed R. Elbestawy, Moustafa Elhamouly, Fuad Saleh, Whad Fayed, Hamada A. Ahmed

TL;DR
Adding 5-10% DDGS with enzymes improves broiler growth, immunity, and gut health while being economically efficient.
Contribution
Demonstrates optimal DDGS inclusion levels with enzyme supplementation for enhanced broiler performance and economics.
Findings
Moderate DDGS (5–10%) with enzymes improved body weight gain and feed conversion ratio.
Enzyme-supplemented diets enhanced immune markers like IgA, IgM, and white blood cell counts.
Economically, 5% DDGS with enzymes provided the highest net profit and lowest cost.
Abstract
This study evaluated the effects of incorporating varying levels of dried distillers’ grains with solubles (DDGS) and non-starch polysaccharide (NSP) enzymes into broiler diets to assess their impact on growth performance, immune function, intestinal histomorphology, and economic efficiency. A total of 240 one-day-old broiler chicks (Avian 48; initial weight 51.57 ± 4.6 g) were randomly assigned to eight dietary treatments (30 birds each). The treatments consisted of four DDGS inclusion levels (0, 5, 10%, or 20%) provided either with or without NSP enzyme supplementation. Growth performance indicated that moderate DDGS inclusion (5–10%) with enzymes improved body weight gain and feed conversion ratio during the starter and grower phases, whereas 20% DDGS without enzymes led to poor performance. While most serum biochemical parameters remained unaffected (p > 0.05), triglycerides and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
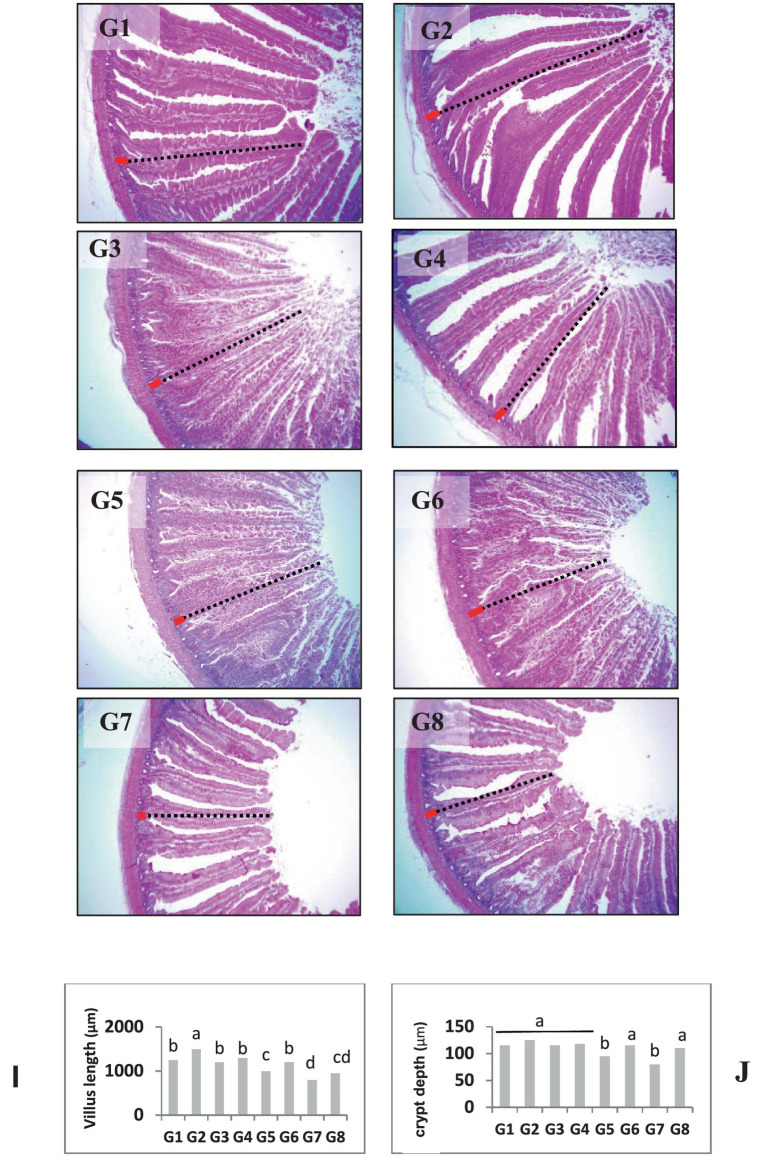 Figure 1
Figure 1| Ingredients, g kg−1 | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz |
|---|---|---|---|---|---|---|---|---|
| Yellow corn, ground | 552 | 576 | 525.7 | 543.2 | 498.5 | 512 | 432.2 | 446 |
| Soybean meal (46%) | 365 | 354 | 331 | 334 | 300 | 310 | 254 | 267 |
| Corn gluten | 16 | 20 | 24 | 19 | 30 | 20 | 33 | 20.5 |
| DDGS | − | − | 50 | 50 | 100 | 100 | 200 | 200 |
| Vegetable oil | 24.3 | 6.5 | 26 | 10.4 | 28 | 14 | 36 | 22 |
| L-Lysine HCL (99%) | 2.2 | 2.4 | 2.8 | 2.7 | 3.3 | 3.1 | 4.1 | 3.8 |
| DL-Meth (99%) | 2.4 | 2.5 | 2.4 | 2.4 | 2.2 | 2.3 | 2 | 2.1 |
| Dicalcium phosphate | 15 | 14.8 | 14.6 | 14.5 | 14.2 | 14.5 | 14.3 | 14.2 |
| Limestone | 13.05 | 13.65 | 13.45 | 13.65 | 13.75 | 13.95 | 14.35 | 14.25 |
| Dietary supplement blend | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 |
| Mycotoxin binder | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 |
| NSP enzymes | − | + | − | + | − | + | − | + |
| Phytase enzyme | + | + | + | + | + | + | + | + |
| Calculated nutrients | ||||||||
| ME (kcal·kg−1 diet) | 3002.37 | 3003.3 | 3002.27 | 3002.13 | 3001.81 | 3001.2 | 3002.6 | 3001.14 |
| Crude protein (%) | 22.86 | 22.85 | 22.85 | 22.84 | 22.83 | 22.80 | 22.84 | 22.80 |
| Ingredients, g kg−1 | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz |
|---|---|---|---|---|---|---|---|---|
| Yellow corn, ground | 618 | 633.5 | 592.7 | 602.5 | 556 | 565.9 | 508.9 | 502 |
| Soybean meal (46%) | 295 | 298 | 258 | 275 | 244 | 260 | 165.7 | 213 |
| Corn gluten | 21.4 | 17 | 32 | 18 | 27 | 15 | 50 | 18.4 |
| DDGS | − | − | 50 | 50 | 100 | 100 | 200 | 200 |
| Vegetable oil | 28 | 13 | 29 | 16.3 | 34.6 | 21.1 | 35.8 | 28 |
| L-Lysine HCL (99%) | 2.1 | 2 | 2.8 | 2.4 | 3 | 2.5 | 4.4 | 3.2 |
| DL-Meth (99%) | 2 | 2 | 1.9 | 1.9 | 1.8 | 1.7 | 1.5 | 1.6 |
| Dicalcium phosphate | 12 | 12.5 | 11.8 | 12.05 | 11.7 | 11.7 | 11.5 | 11.7 |
| Limestone | 11.45 | 11.85 | 11.75 | 11.7 | 11.85 | 11.95 | 12.15 | 11.95 |
| Dietary supplement blend | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 |
| Mycotoxin binder | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 |
| NSP enzymes | − | + | − | + | − | + | − | + |
| Phytase enzyme | + | + | + | + | + | + | + | + |
| Calculated nutrients | ||||||||
| ME (kcal·kg−1 diet) | 3102.75 | 3103.45 | 3102.04 | 3102.99 | 3102.84 | 3102.00 | 3102.1 | 3102.16 |
| Crude protein (%) | 20.63 | 20.64 | 20.65 | 20.64 | 20.63 | 20.69 | 20.61 | 20.68 |
| Group | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz | |
|---|---|---|---|---|---|---|---|---|---|
| Body weight gain (g) | |||||||||
| 0–14 days | 372.33 ± 2.27d | 413.733 ± 2.18a | 375.73 ± 1.54d | 406.82 ± 1.13b | 373.95 ± 1.32d | 403.65 ± 0.57b | 363.56 ± 1.74e | 391.03 ± 1.02c | 0.001*** |
| 15–28 days | 844.58 ± 10.2c | 930.26 ± 9.92a | 861.71 ± 5.3bc | 882.24 ± 16.2b | 850.18 ± 7c | 879.22 ± 2.78b | 772.05 ± 15.9e | 815.13 ± 5.57d | 0.001*** |
| Final BWG (0–28 days) | 1216.7 ± 12.5cd | 1342.8 ± 5.5a | 1237.6 ± 7.9c | 1289.1 ± 4.66b | 1225.13 ± 9cd | 1282.87 ± 9.9b | 1135.6 ± 20.4d | 1206.2 ± 7.9cd | 0.002** |
| Feed consumption (g/chick) | |||||||||
| 0–14 days | 522.83 ± 7.16d | 546 ± 7.50cd | 548.67 ± 2.97c | 557.62 ± 2.57c | 551.07 ± 5.69c | 560.57 ± 3.83c | 624.57 ± 3.81b | 643.67 ± 3.66a | 0.004** |
| 15–28 days | 1620.5 ± 34.95b | 1670.6 ± 12.41ab | 1634.7 ± 25.4ab | 1641.6 ± 24.1ab | 1654.1 ± 25.44ab | 1687.8 ± 31.9a | 1670.7 ± 0.98ab | 1676.4 ± 28.8a | 0.021* |
| Total FI (0–28 days) | 2144.33 ± 24c | 2216.6 ± 32.78bc | 2183.37 ± 40c | 2198.2 ± 42.6c | 2207.17 ± 9.7bc | 2248.37 ± 56ab | 2296.3 ± 24.8ab | 2322.1 ± 38.7a | 0.008** |
| Feed conversion ratio (FCR) | |||||||||
| 0–14 days | 1. 39 ± 0.012bc | 1.31 ± 0.014d | 1.45 ± 0.031b | 1.37 ± 0.007cd | 1.46 ± 0.01b | 1.38 ± 0.032bc | 1.71 ± 0.01a | 1.64 ± 0.007a | 0.001*** |
| 15–28 days | 1.92 ± 0.029c | 1.79 ± 0.014d | 1.89 ± 0.027cd | 1.85 ± 0.026cd | 1.94 ± 0.031c | 1.91 ± 0.035c | 2.16 ± 0.009a | 2.03 ± 0.02b | 0.003** |
| Cumulative FCR (0–28 days) | 1.76 ± 0.022bc | 1.65 ± 0.042d | 1.76 ± 0.015bc | 1.71 ± 0.029cd | 1.8 ± 0.068b | 1.75 ± 0.051bc | 2.02 ± 0.017a | 1.93 ± 0.02a | 0.012* |
| Parameter | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz | |
|---|---|---|---|---|---|---|---|---|---|
| Triglyceride mg/dL | 164.29 ± 0bc | 174.15 ± 1.4a | 165.24 ± 3.3bc | 170.07 ± 3.6ab | 164.59 ± 2bc | 170.1 ± 3.6ab | 159.1 ± 0.95c | 166.67 ± 2.38abc | 0.019* |
| Cholesterol mg/dL | 218.6 ± 14.9 | 226.95 ± 0.3 | 195.42 ± 4.18 | 201.89 ± 7.42 | 202.16 ± 1.7 | 203.77 ± 3.1 | 215.63 ± 15.8 | 220.76 ± 28.55 | 0.645 NS |
| total protein g/dL | 2.82 ± 0.12 | 2.91 ± 0.29 | 2.86 ± 0.09 | 3.06 ± 0.09 | 2.89 ± 0.06 | 3.02 ± 0.03 | 2.8 ± 0.01 | 2.98 ± 0.11 | 0.999 NS |
| Albumin g/dL | 1.5 ± 0.02 | 1.54 ± 0.03 | 1.45 ± 0.04 | 1.55 ± 0.03 | 1.45 ± 0.07 | 1.45 ± 0.02 | 1.45 ± 0.03 | 1.42 ± 0.04 | 0.994 NS |
| Globulin g/dl | 1.35 ± 0.04c | 1.37 ± 0.02 bc | 1.43 ± 0.02 abc | 1.5 ± 0.08 a | 1.39 ± 0.03 abc | 1.48 ± 0.03 ab | 1.32 ± 0.02 c | 1.4 ± 0.03 abc | 0.032* |
| A/G ratio | 1.12 ± 0.05 | 1.12 ± 0.03 | 1.01 ± 0.03 | 0.99 ± 0.08 | 1.04 ± 0.04 | 0.98 ± 0.01 | 1.1 ± 0.02 | 1.01 ± 0.03 | 0.24 NS |
| Uric acid mg/dL | 5.77 ± 0.12 | 5.92 ± 0.28 | 5.92 ± 0.18 | 5.9 ± 0.31 | 5.95 ± 0.26 | 5.83 ± 0.4 | 5.77 ± 0.4 | 5.83 ± 0.15 | 0.14 NS |
| Creatinine mg/dL | 4.9 ± 0.39 | 5.32 ± 0.33 | 5.21 ± 0.65 | 5.36 ± 0.57 | 5.49 ± 0.12 | 5.18 ± 0.56 | 5.13 ± 0.36 | 5.2 ± 0.6 | 0.12 NS |
| ALT U/L | 12.22 ± 1.01 | 12.8 ± 0.58 | 12.8 ± 1.16 | 12.8 ± 0.58 | 12.8 ± 1.16 | 12.8 ± 0.58 | 12.22 ± 1.01 | 12.8 ± 0.58 | 0.998 NS |
| AST U/L | 267.14 ± 3.6 | 268.3 ± 3.5 | 267.72 ± 5.8 | 268.88 ± 2.02 | 263.65 ± 8.3 | 265.4 ± 4.4 | 267.72 ± 3.1 | 268.9 ± 8.79 | 0.997 NS |
| Parameter | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz | |
|---|---|---|---|---|---|---|---|---|---|
| Relative weight of lymphoid organs (%) | |||||||||
| Spleen % | 0.1 ± 0.01 | 0.1 ± 0.007 | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.616NS |
| Thymus % | 0.31 ± 0.01 | 0.315 ± 0.01 | 0.31 ± 0.01 | 0.31 ± 0.014 | 0.32 ± 0.01 | 0.32 ± 0.01 | 0.315 ± 0.015 | 0.33 ± 0.01 | 0.824NS |
| Bursa of Fabricius % | 0.11 ± 0.01b | 0.142 ± 0.01a | 0.12 ± 0.01ab | 0.14 ± 0.02ab | 0.12 ± 0.01ab | 0.13 ± 0.01ab | 0.11 ± 0.003ab | 0.14 ± 0.01ab | 0.016* |
| Immunoglobulins (mg/mL) | |||||||||
| IgA (mg/mL) | 0.46 ± 0.03c | 0.48 ± 0.05bc | 0.5 ± 0.03a | 0.51 ± 0.06 a | 0.49 ± 0.05 ab | 0.51 ± 0.08 a | 0.46 ± 0.03 c | 0.5 ± 0.04 a | 0.009** |
| IgM (mg/mL) | 0.63 ± 0.05c | 0.66 ± 0.06ab | 0.66 ± 0.02ab | 0.7 ± 0.02a | 0.64 ± 0.01bc | 0.69 ± 0.03a | 0.61 ± 0.02c | 0.69 ± 0.03a | 0.005** |
| Antioxidative status (U/mL) | |||||||||
| GPX (U/mL) | 259 ± 1.7 | 260 ± 1.8 | 264 ± 2.1 | 269 ± 1.33 | 268 ± 1.3 | 270 ± 2.33 | 260 ± 2.8 | 272 ± 1.32 | 0.197NS |
| Hematological parameters | |||||||||
| WBCs (×103 /mm3) | 20.37 ± 1.17b | 21.24 ± 1.14b | 27.05 ± 0.88a | 28.05 ± 0.92a | 27.58 ± 0.58a | 28.69 ± 1.29a | 21.52 ± 0.26b | 26.94 ± 0.54a | 0.02* |
| Lymphocytes % | 69.92 ± 0.24a | 68.50 ± 0.44a | 65.24 ± 0.56b | 62.19 ± 0.43c | 65.65 ± 1.02b | 62.68 ± 0.93c | 65.16 ± 0.78b | 65.16 ± 0.44b | 0.003** |
| Heterophiles % | 20.54 ± 0.36c | 21.61 ± 0.55c | 27.28 ± 0.53b | 30.29 ± 0.45a | 27.30 ± 0.74b | 29.89 ± 0.94a | 22.38 ± 0.57c | 29.27 ± 0.57a | 0.023* |
| H/L ratio | 0.29 ± 0c | 0.315 ± 0c | 0.418 ± 0.01b | 0.487 ± 0.01a | 0.415 ± 0.02b | 0.476 ± 0.02a | 0.343 ± 0c | 0.449 ± 0.13ab | 0.011* |
| Parameter | Control | Control + Enz | 5% DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz | ||
|---|---|---|---|---|---|---|---|---|---|---|
| ND titers | Day 7 | 7.33 ± 0.24a | 7 ± 0ab | 6.66 ± 0.24ab | 6.65 ± 0.47ab | 5.66 ± 0.24c | 5.66 ± 0.24c | 6.33 ± 0.24bc | 6.33 ± 0.24bc | 0.001*** |
| Day 14 | 2.97 ± 0.41ab | 2.97 ± 0.41ab | 4.62 ± 0.63a | 3.64 ± 0.47ab | 2.57 ± 0.63b | 2.57 ± 0.63b | 2.97 ± 0.41ab | 2.27 ± 0.48b | 0.001*** | |
| Day 21 | 5.25 ± 1.18 | 6.43 ± 0.57 | 5.33 ± 0.24 | 5.33 ± 0.24 | 5.33 ± 0.24 | 6.3 ± 0.62 | 5.94 ± 0.82 | 6.3 ± 0.62 | 0.69NS | |
| Day 28 | 4.95 ± 0.53c | 5.99 ± 0.41abc | 5.33 ± 0.24c | 5.99 ± 0.41abc | 6.66 ± 0.24ab | 5.66 ± 0.24bc | 6.99 ± 0.41a | 6 ± 0abc | 0.017* | |
| H5N8 titers | Day 7 | 9.11 ± 1.06 ab | 8.33 ± 0.24 b | 8.31 ± 0.62b | 10.99 ± 0.41a | 9.31 ± 0.62ab | 9.97 ± 0.82ab | 10.33 ± 0.24a | 9.99 ± 0.41ab | 0.002** |
| Day 14 | 5.66 ± 0.24 bc | 5.94 ± 0.82 bc | 6.64 ± 0.62abc | 7.66 ± 0.24a | 5.33 ± 0.24c | 6.64 ± 0.62abc | 7.33 ± 0.24ab | 5.79 ± 0.76bc | 0.024* | |
| Day 21 | 4 ± 0 ab | 3.28 ± 0.63 b | 4.66 ± 0.24ab | 5 ± 0a | 3.66 ± 0.24ab | 3.61 ± 0.63ab | 3.61 ± 0.63ab | 4.65 ± 0.47ab | 0.041* | |
| Day 28 | 2 ± 0 ab | 2.66 ± 0.24 ab | 2 ± 0ab | 3.29 ± 0.47a | 2.32 ± 0.24ab | 3.14 ± 0.59ab | 1.9 ± 0.71b | 3.1 ± 0.5ab | 0.038* | |
| H9N2 titers | Day 7 | 7.66 ± 0.24 ab | 8.33 ± 0.24 a | 7.66 ± 0.24ab | 7.64 ± 0.62ab | 6.65 ± 0.47b | 7.32 ± 0.47ab | 7.99 ± 0.41a | 8.33 ± 0.24a | 0.014* |
| Day 14 | 3.92 ± 0.71 bc | 3.61 ± 0.63 c | 5.65 ± 0.47ab | 4.29 ± 0.63abc | 3.9 ± 0.66bc | 4.31 ± 0.69abc | 5.99 ± 0.41a | 4 ± 0bc | 0.007** | |
| Day 21 | 6.58 ± 1.18 abc | 6.16 ± 1.44 abc | 5 ± 0abc | 4.07 ± 1.28c | 4.53 ± 1.04bc | 6.96 ± 0.71abc | 8.23 ± 1.25a | 7.62 ± 0.85 ab | 0.019* | |
| Day 28 | 9.75 ± 0.38ab | 10 ± 0ab | 9.63 ± 0.37b | 9.31 ± 0.62b | 9.66 ± 0.24b | 9.32 ± 0.47b | 10.99 ± 0.41a | 10.33 ± 0.24ab | 0.046* | |
| Parameter | Control | Control + Enz | 5%DDGS | 5% DDGS + Enz | 10% DDGS | 10% DDGS + Enz | 20% DDGS | 20% DDGS + Enz | |
|---|---|---|---|---|---|---|---|---|---|
| Total feed cost | 67.80 ± 7.80b | 68.15 ± 8.1b | 68.11 ± 8.1b | 65.96 ± 6.99b | 69.39 ± 9.1b | 67.98 ± 9.8b | 72.16 ± 7.11a | 71.23 ± 7.12a | 0.003** |
| Total costs | 142.8 ± 8.12b | 143.15 ± 10.1b | 143.11 ± 11.1b | 140.96 ± 10.14c | 144.39 ± 13.1b | 142.98 ± 12.1b | 147.16 ± 11.1a | 146.23 ± 14.1a | 0.009** |
| Value of final bird | 146.49 ± 4.90d | 167.73 ± 7.33a | 150.63 ± 6.33c | 157.89 ± 7.80b | 148.67 ± 4.85d | 156.59 ± 5.90b | 132.09 ± 3.29e | 146.54 ± 5.44d | 0.001*** |
| Total return | 156.49 ± 16.4d | 177.73 ± 17.37a | 160.63 ± 16.4c | 167.89 ± 18.19b | 158.67 ± 17.2d | 166.59 ± 15.2b | 142.09 ± 14.1e | 156.54 ± 14.2d | 0.001*** |
| Net profit | 13.69 ± 2.69e | 34.58 ± 5.8a | 17.52 ± 5.22d | 26.93 ± 3.14b | 14.28 ± 4.1e | 23.61 ± 3.1c | −5.07 ± 1.15g | 10.31 ± 2.58f | 0.001*** |
| Feed costs/total returns | 43.33 ± 3.34c | 38.34 ± 4.12 e | 42.40 ± 4.14 c | 39.29 ± 3.12 e | 43.73 ± 4.15 c | 40.81 ± 4.18 d | 50.78 ± 5.17 a | 45.50 ± 5.17b | 0.002** |
| Net profit/total costs | 9.59 ± 1.19e | 24.16 ± 4.15a | 12.24 ± 4.12d | 19.10 ± 3.14b | 9.89 ± 4.15e | 16.51 ± 5.14c | −3.45- ± 1.14g | 7.05 ± 2.15f | 0.001** |
| Benefit/cost ratio | 109.59 ± 5.9e | 124.16 ± 10.1a | 112.24 ± 12.1d | 119.10 ± 10.14b | 109.89 ± 10.2e | 116.51 ± 11.1c | 96.55 ± 5.15g | 107.05 ± 10.2f | 0.001*** |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology · Nanocomposite Films for Food Packaging · Aquaculture Nutrition and Growth
Introduction
1
The poultry industry constantly seeks cost-effective and sustainable feed ingredients to enhance profitability while ensuring optimal bird performance. Dried distillers’ grains with solubles (DDGS), a byproduct of ethanol production, has emerged as a valuable alternative protein and energy source in broiler diets due to its high nutritional content, wide availability, and potential to lower feed costs. Moreover, DDGS has been recognized as a functional feed additive that can enhance poultry productivity through microbiota-mediated mechanisms, providing further support for its use in sustainable poultry production (1). Global biofuel production is expected to increase by over 20% between 2022 and 2027, emphasizing the rising significance of DDGS as a co-product (2). However, the inclusion of DDGS in poultry diets is often restricted due to its high levels of non-starch polysaccharides (NSPs). Corn DDGS contains approximately 26.5% total NSPs, including 3.55% soluble and 23.5% insoluble fractions, along with 21.5% arabinoxylans and 0.32% β-glucans (3). These components can negatively impact growth performance, gut health, and nutrient absorption efficiency.
Previous research indicated that moderate levels of DDGS can support broiler growth when properly balanced within the diet. Young broilers, in particular, are sensitive to feed quality because their digestive system are not fully developed until approximately 2 weeks of age (4). Water-soluble NSPs, when included in young chick diets, have been shown to reduce nutrient digestion and absorption by increasing digesta viscosity in the gut (5). Consequently, the use of exogenous enzymes, such as NSP enzymes, has been suggested as a strategic approach to enhance nutrient utilization by breaking down fiber components, thereby improving digestibility and mitigating the potential negative effects associated with DDGS (6–10).
Broiler growth performance is significantly influenced by dietary composition across different feeding phases, as nutrient requirements vary from the starter to finisher phases. Assessing the impact of DDGS inclusion during these phases is essential for understanding its effects on body weight gain, feed efficiency, and overall productivity. Furthermore, immune responses are crucial for maintaining bird health and performance, with indicators such as immunoglobulin levels and antibody titers providing valuable insight into immune function. Intestinal histomorphology, particularly villus height and crypt depth, also serves as a key measure of gut health and nutrient absorption efficiency.
The present study investigated the effects of different levels of DDGS, with or without enzymes supplementation, on growth performance, immune responses, and intestinal histomorphology in broilers across feeding phases. The findings will contribute to optimizing DDGS utilization in poultry nutrition and support the maintenance of health and performance standards.
Materials and methods
2
All animal procedures and sample collection were conducted in accordance with national and institutional guidelines for the care and use of broilers and were approved by the Animal Ethics Committee of the Faculty of Veterinary Medicine, Menoufia University (Approval No. MN/VET/NUT/25/02/03/01).
Experimental diets
2.1
The experiment employed an experimental design involving eight distinct dietary treatments, conducted across two feeding phases: starter (0–14 days), and grower (15–28 days). Diets were formulated in accordance with the nutrient specifications for Avian 48 broilers. All experimental groups had similar raw material and nutrient contents, except for the DDGS and enzyme inclusion. The diets were provided in mash form. DDGS used in this experiment was imported from the United States, and its main nutrient composition as follows: Dry matter (DM), 88.42%; Crude protein (CP), 25%; Ether extract (EE), 8%; Crude fiber (CF), 7.9%; Ash, 4.79%; Nitrogen-free extract (NFE), 42.73%; and apparent metabolizable energy (AME), 2,525 Kcal/kg. Smart NSP enzymes® was the commercial feed additive used, obtained from Devenish Nutrition Ltd., 96 Duncrue Street, Belfast, BT3 9AR, Northern Ireland. This product contained a combination of 5 active enzymes (7,503 U/g xylanase, 2,500 U/g glucoamylase, 1,443 U/g β-glucanase, 375 U/g pectinase, 144 U/g cellulase) produced by advanced fermentation fungal strains. This enzyme provides an energy matrix of 1,050,000 Kcal/kg ME. The NSP enzymes were supplemented at a consistent rate of 100 mg·kg^−1^ in the relevant diets. Additionally, Smart phytase enzyme® (10,000 FTU/g phytase enzyme), derived from Escherichia coli (E. coli), was also obtained from the commercial company Devenish Nutrition Ltd., Belfast, Northern Ireland. The composition of the treatment diets during the starter and grower phases is presented in Tables 1, 2, respectively.
Broiler management
2.2
A total of 240 one-day-old broiler chicks (Avian 48; mean initial body weight 51.57 ± 4.6 g) were randomly allocated to 8 experimental groups in a completely randomized design, with 30 birds per group (three replicates of 10 birds each). The study was conducted over a 28-day feeding trial. Chicks were housed in floor pens with a litter-floor system, utilizing wood shavings as bedding material. All broilers were provided ad libitum access to experimental diets and water. Environmental conditions, including temperature, humidity, ventilation, and lighting, were monitored and adjusted according to the management guidelines for Avian 48 broilers. Broiler performance parameters, including body weight gain (BWG) and feed intake, were recorded weekly to evaluate growth performance and feed efficiency. Additionally, the feed conversion ratio (FCR) was calculated as the ratio of cumulative feed intake to cumulative body weight gain throughout the experimental period.
Chemical analysis
2.3
The components in the samples were determined using standard procedures of the Association of Official Analytical Chemists (11) for DM (934.01), CP (976.05), CF (962.09), and EE (920.39). ME values were calculated based on the analyzed chemical composition of each ingredient using the conversion factors described by Janssen (12) and expressed as a weighted sum according to their inclusion levels in the diet.
Vaccination program
2.4
Chicks were vaccinated at day 7 of age with EgyFlu®-ND7-3 in 1 (HARVAC, Harbin, China) via subcutaneous injection, providing inactivated protection against avian influenza virus (AIV; H5N8 and H9N2 subtypes) and RG-Newcastle disease virus (NDV; HB38 strain). Additionally, ocular administration of MEVAC LaSota+ H120® Elite (MEVAC, Egypt) was performed, delivering live lentogenic NDV (LaSota strain, 10^6^–10^7^ TCID₅₀) and attenuated infectious bronchitis virus (IBV; H120 strain, Massachusetts serotype, 10^3.5^–10^4.5^ EID₅₀). On day 14, booster vaccinations were administered via eye drops using RINNOVAC ELI-7®, a recombinant live NDV vaccine (VG/VG + F7 strains), and MEVAC IBD 818®, an intermediate-plus live vaccine against infectious bursal disease (IBD).
Blood sampling and biochemical analysis
2.5
Blood samples were collected weekly from three birds per replicate (n = 9 birds per treatment) at 7, 14, 21, and 28 days of age to assess hemagglutination inhibition (HI) titers against NDV and AIV. At the end of the trial, nine birds per group (three per replicate) were randomly selected for blood collection. Two types of samples were taken from the wing vein: one into plain tubes for serum separation (centrifuged at 3000 rpm for 10 min and stored at −18 °C), and the other into ethylene diamine tetra-acetic acid (EDTA) tubes for hematological analysis. Serum samples were analyzed using commercial diagnostic kits for total protein and its fractions (albumin and globulin), cholesterol, triglycerides, uric acid, creatinine, glutathione peroxidase (GPX), aspartate aminotransferase (AST), alanine aminotransferase (ALT), and immunoglobulin levels (IgA, IgM). Absorbance readings were obtained using a UV–visible spectrophotometer (UVS-85, Acculab, USA).
Immune organ evaluation
2.6
At the end of the experimental period, nine birds from each group (three per replicate) were randomly selected for immune organ evaluation. The Bursa of Fabricius, spleen, and thymus were carefully dissected and weighed to determine their absolute weights. Relative organ weights were calculated as (organ weight/live body weight) × 100 to account for individual variation.
Intestinal histomorphological analysis
2.7
At the end of the trial, jejunal samples were collected from two birds per replicate within each treatment. A 3-cm mid-jejunal segment (between the bile duct entry and Meckel’s diverticulum) was excised, rinsed with physiological saline to remove luminal contents, and fixed in 10% neutral-buffered formalin (100 mL 40% formaldehyde, 4 g monobasic phosphate, 6.5 g dibasic sodium phosphate, and 900 mL distilled water) for 18–24 h. Fixed tissues were dehydrated, embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin and eosin (H&E) as described by Kim et al. (13). Slides were examined under a light microscope, and morphometric measurements (villus height and crypt depth) were performed using QuPath (version 0.4.3). Villus height (VH, μm) was measured from the tip of the villus to the villus–crypt junction, while crypt depth (CD, μm) was measured from the base of the villus to the bottom of the crypt. For each bird, ten well-oriented villi were evaluated in triplicate, and mean values were used for statistical analysis (see Figure 1).
Histopathological examination of chicken jejunum samples from different groups. G1: control group, G2: basal diet plus NSP enzymes group, G3: 5% DDGS without NSP enzymes, G4: 5% DDGS plus NSP enzymes, G5: 10% DDGS without NSP enzymes, G6: 10% DDGS plus NSP enzymes, G7: 20% DDGS without enzymes, G8: 20% DDGS plus NSP enzymes, showing villus length (black dotted line), and crypt depth (red line). Scale bar = 500 μm. Image analysis of the studied groups for measuring villus length (I) and crypt depth (J). Bars with different letters are significantly different (p < 0.05).
Economic evaluation
2.8
An economic evaluation was conducted as a supportive and descriptive analysis to contextualize the biological performance results of broilers fed diets containing different levels of DDGS, with or without enzyme supplementation. Feed cost and return-related parameters were calculated based on actual feed intake, final live body weight, and prevailing market prices using standard economic calculations (14). Economic efficiency was expressed as the benefit–cost ratio (BCR) to provide practical insight into the potential economic implications of the dietary treatments.
Statistical analysis
2.9
Data were expressed as mean ± standard error (SE). All results were analyzed using two-way analysis of variance (ANOVA), and differences among means were evaluated using the least significant difference (LSD) post hoc test. Statistical analyses were performed using SPSS software (version 16.0; SPSS Inc., Chicago, IL, USA, 2001). A significance level of (p < 0.05) was considered statistically significant.
Results
3
Growth performance
3.1
Growth performance results in Table 3 showed that dietary DDGS levels and enzyme supplementation significantly (p < 0.05) affected BWG, FI, and FCR. During the starter period (0–14 days), the highest BWG was recorded in broilers fed the enzyme-supplemented basal diet, whereas the poorest growth occurred in birds receiving 20% DDGS without enzymes. Moderate DDGS levels (5–10%) combined with enzymes also improved BWG. Similar patterns were observed during the grower phase (15–28 days) and over the overall experimental period (0–28 days).
Feed intake was also significantly (p < 0.05) affected. In the early phase (0–14 days), diets with higher DDGS levels supplemented with enzymes were associated with higher feed intake, conversely, the lowest FI was observed in birds fed the control diet without DDGS. A similar trend was observed in the grower phase (15–28 days). Total FI (0–28 days) was higher in birds receiving enzyme-supplemented, high-DDGS diets. Feed utilization efficiency showed a consistent pattern. In the starter stage, birds receiving enzyme-supplemented diets recorded the most efficient FCR, while the poorest values were observed in diets containing high DDGS without enzymes. FCR progressively increased with DDGS inclusion across the entire period (0–28 days), though enzyme supplementation consistently provided a partial improvement.
Intestinal histomorphology
3.2
Histopathological examination and morphometric analysis of the intestinal samples revealed distinct structural variations among the dietary treatments. Birds receiving enzyme-supplemented diets, particularly those fed the basal diet or moderate DDGS inclusion levels (5–10%), exhibited well-developed intestinal villi characterized by increased villus height and comparatively reduced crypt depth. Conversely, birds fed the highest level of DDGS (20%) without enzyme inclusion showed marked reductions in villus height and increased crypt depth. Morphometric analysis confirmed these observations; as villus height (VH) was significantly lower (p < 0.05) in the high-DDGS group without enzymes. No significant differences (p > 0.05) were observed among the control, enzyme-supplemented, or moderate DDGS groups. Crypt depth (CD) remained statistically similar across most treatments, except in the high-DDGS group, which showed shallow crypts.
Physiological and immune status
3.3
Serum biochemical parameters
3.3.1
Serum biochemical parameters presented in Table 4 showed significant differences (p < 0.05) among treatments in serum triglycerides and globulin levels. Conversely, no significant differences (p > 0.05) were found for cholesterol, total protein, albumin, A/G ratio, uric acid, creatinine, ALT, and AST levels. Specifically, groups receiving enzyme supplementation showed statistically higher levels in triglycerides and globulin compared to specific control groups. Furthermore, cholesterol levels were marginally reduced in DDGS-fed groups.
Immune response, antioxidant status, and hematological parameters
3.3.2
As presented in Table 5, the inclusion of DDGS, whether with or without enzyme supplementation, did not significantly (p > 0.05) influence the relative weights of the thymus and spleen. However, the relative weight of the Bursa of Fabricius was significantly influenced (p < 0.05) by the treatments. Specifically, Bursa weight was significantly (p < 0.05) higher in the enzyme-supplemented control group compared to the control group without enzymes, whereas other DDGS-fed groups with or without enzyme inclusion showed intermediate values.
Regarding humoral immunity, serum IgA and IgM levels were significantly elevated (p < 0.05) in birds receiving enzyme-supplemented DDGS diets, particularly at 5–10% inclusion levels. Glutathione peroxidase (GPX) activity showed no significant differences (p > 0.05) among the groups. However, numerically higher values were observed in enzyme-supplemented DDGS treatments, especially at 10 and 20% inclusion levels. Hematological results revealed significantly higher total white blood cell counts (p < 0.05) in broilers fed diets containing 5 and 10% DDGS, particularly when combined with enzyme supplementation, compared to the control groups. In parallel, lymphocyte percentages were significantly reduced, while heterophil percentages and H/L ratios were markedly elevated in these groups.
Serum antibody titers
3.3.3
As shown in Table 6, the ND titers varied significantly (p < 0.05) among groups on Day 7, Day 14, and Day 28, while no significant difference (p > 0.05) was observed on Day 21. Initially (Day 7), the highest titers were recorded in the control diets, but by Day 28, birds fed 10% DDGS showed significantly improved responses compared to the control group. Regarding H5N8, significant differences (p < 0.05) were detected across all measurement days. Birds fed 5% DDGS with enzymes had the significantly highest titers at Day 7, while the lowest titer at Day 28 was recorded in birds receiving 20% DDGS without enzyme supplementation. Overall, H5N8 titers declined over time. In contrast, H9N2 titers also showed significant differences (p < 0.05) across all days and generally increased over time. The highest levels were observed significantly at Day 28 in birds fed 20% DDGS.
Economical evaluation
3.4
The economic-related parameters of broilers fed the experimental diets are summarized in Table 7. Statistically significant differences (p < 0.01) were detected among the experimental groups for the calculated economic indicators.
Discussion
4
The results indicated that enzyme supplementation significantly improved BWG and FCR, particularly in the basal diet, highlighting the positive role of enzyme inclusion, especially non-starch polysaccharide-degrading enzymes, in enhancing early nutrient digestibility and utilization (6–10). The observation that moderate levels of DDGS (5–10%) combined with enzymes also showed improved BWG is consistent with earlier findings (15), whereas birds fed the highest DDGS level (20%) without enzymatic support displayed the poorest growth, is consistent with previous studies that report reduced nutrient availability associated with high DDGS inclusion in the absence of enzymes (16).
The poorest performance observed in the high DDGS group without enzymes further emphasizes the challenges posed by high fiber and associated anti-nutritional compounds. Although enzyme inclusion in high-DDGS diets led to numerical improvements, the gains did not match those achieved with enzyme-supplemented diets free from high DDGS. These findings highlight the partial efficacy of enzymes in mitigating the negative effects of high DDGS inclusion, in line with the observations of previous studies (17). By the end of the period (0–28 days), the cumulative effect demonstrated that enzyme-supplemented groups at low to moderate DDGS levels maintained higher gains, while birds on unsupplemented high-DDGS diets showed the lowest BWG (18).
The significant effect on feed intake in the early phase (p < 0.05), where diets with higher DDGS levels plus enzymes stimulated greater feed consumption, is possibly due to compensatory feeding behavior and the higher fiber content of DDGS (4). Birds fed enzyme-supplemented diets consumed more feed than the control group (19, 20). These findings are consistent with reports indicating that early-phase FI increases with moderate DDGS inclusion but may decline with extended feeding and higher levels (21). The higher total FI (0–28 days) in enzyme-supplemented, high-DDGS diets further emphasizes the role of enzymes in enhancing palatability and digestion efficiency in fiber-rich diets.
The consistent pattern of FCR across phases, where DDGS inclusion progressively increased FCR, reflects the negative impact of dietary fiber and associated anti-nutritional factors on nutrient utilization. Enzyme supplementation consistently improved FCR across all inclusion levels, but the improvement was only partial and did not completely restore feed efficiency to the enzyme-supplemented control. These findings indicate that DDGS inclusion reduces nutrient digestibility and feed efficiency, while enzyme supplementation can alleviate, but not eliminate, the adverse effects. These findings align with the pattern documented previously (22).
The observation that birds receiving enzyme-supplemented diets exhibited well-developed intestinal villi with increased height and reduced crypt depth indicates enhanced absorptive capacity and improved mucosal integrity, supporting the role of enzymes in promoting gut health. Similar improvements in intestinal morphology with enzyme supplementation have been observed in previous studies (23), conversely, the marked reductions in villus height and increased crypt depth observed in birds fed the highest level of DDGS without enzyme inclusion suggest compromised intestinal function and nutrient absorption. These alterations are in line with reports indicating that DDGS levels above 15% may adversely affect intestinal morphology (24).
The morphometric analysis, confirming that the high-DDGS group without enzymes had significantly lower villus height and shallow crypts (p < 0.05), reflects the negative impact of high fiber and anti-nutritional factors. The shallow crypts suggest reduced epithelial turnover in response to impaired villus growth, which is a key indicator of poor gut health and reduced capability for nutrient absorption.
The finding that most serum biochemical parameters (cholesterol, total protein, albumin, A/G ratio, uric acid, creatinine, ALT, and AST) remained unaffected (p > 0.05) partially agrees with earlier observations indicating no significant alterations in plasma biochemical indices with dietary DDGS inclusion up to 20% in poultry (25). This suggests that incorporating DDGS, with or without NSP enzymes, does not adversely affect overall broiler metabolism, and did not compromise liver or kidney function. However, the observation of statistically higher levels in triglycerides and globulin (p < 0.05) in enzyme-supplemented groups compared to specific control groups indicates enhanced nutrient utilization or metabolic activity related to these components. Similar trends were described in studies involving DDGS inclusion up to 16% (26). The marginal reduction in cholesterol levels observed in DDGS-fed groups is possibly due to the cholesterol-lowering effect of dietary fiber inherent in DDGS, as previously noted (27).
The finding that Bursa of Fabricius weight was significantly influenced (p < 0.05) by the treatments, contrasting with the thymus and spleen, partially supports previous findings (27, 28), which reported no negative impact of DDGS (up to 15–20%) on immune development. The significantly higher bursa weight observed in the enzyme-supplemented control group, with other DDGS-fed groups showing intermediate values, suggests enhanced immune tissue development without compromising physiological function. The observation that IgA and IgM levels were significantly elevated (p < 0.05) in enzyme-supplemented DDGS diets, reflecting enhanced immune responsiveness, aligns with findings attributing improved antibody levels to the prebiotic effects of yeast components present in DDGS (29, 30). While Glutathione peroxidase (GPX) activity showed no significant differences (p > 0.05), the numerically higher values observed in enzyme-supplemented DDGS treatments support previous findings that DDGS may enhance antioxidant enzyme activity through its polyunsaturated fatty acid content and sulfur-containing amino acids, as indicated (31, 32). The revealed hematological shifts, including significantly higher WBC counts and H/L ratios in DDGS-enzyme groups, may reflect physiological immune stimulation or mild systemic stress, potentially driven by increased metabolic demand or the presence of DDGS fermentation by-products. This interpretation aligns with the findings that elevated WBCs and H/L ratios are reliable indicators of immune activation and stress responses in poultry, as previously documented (33).
The significant variation in ND titers across different days, with 10% DDGS showing significantly improved responses compared to the control group by Day 28, suggests potential long-term immunomodulatory effects of the diet. Regarding H5N8, the overall decline in titers over time could be attributed to environmental exposure to circulating H5N8 viral particles, natural waning of immunity, and reduced vaccine efficacy over time, in agreement with earlier observations (34). In contrast, the significant increase in H9N2 titers over time, with the highest levels observed significantly at Day 28 in birds fed 20% DDGS, indicates an enhanced late-phase immune response to this antigen in the higher DDGS inclusion groups.
Economic efficiency was evaluated as a secondary outcome derived from growth performance and feed utilization parameters. The observed variation in economic indicators, where the highest total cost was observed in broilers fed the 20% DDGS diet without enzymes, whereas the lowest cost was recorded in birds receiving the 5% DDGS diet supplemented with enzymes, a trend that is consistent with the findings of reports indicating a reduction in feed cost per kilogram of gain when moderate levels of DDGS were combined with enzyme supplementation (35). Although total costs were relatively comparable among most treatments, broilers fed the basal diet with enzyme supplementation and those receiving 5% DDGS plus enzymes achieved the highest final bird price and total return (p < 0.01), reflecting superior growth performance and feed utilization. Consequently, these groups also recorded the highest net profits, followed by the 10% DDGS with enzyme supplementation group. In contrast, the negative economic returns observed in broilers fed 20% DDGS without enzymes highlight the economic penalty associated with reduced nutrient availability and impaired performance, likely due to increased levels of anti-nutritional factors at high DDGS inclusion. These findings are consistent with previous reports (15), confirming that enzyme supplementation is a critical strategy for enhancing economic efficiency and maximizing profitability when incorporating DDGS into broiler diets.
Conclusion
5
Dietary inclusion of DDGS at moderate levels (5–10%) throughout the starter, and grower phases, when combined with NSP enzyme supplementation, improved broiler performance, immune response, intestinal morphology, and overall economic efficiency without negative effects on metabolism. Although the control diet with enzyme supplementation showed the best overall performance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bao G Wei H He D Wang D Ding B Zhang J . Dietary supplementation with corn distillers’ dried grains with solubles improves egg quality in Haidong chickens by promoting beneficial gut microbiota and modulating serum biochemical profiles. Front Microbiol. (2025) 16:1709407. doi: 10.3389/fmicb.2025.1709407, 41395495 PMC 12698570 · doi ↗ · pubmed ↗
- 2Biofuels International. Global biofuel demand to grow 20% in next five years. Biofuels Int. (2023). Available online at: https://biofuels-news.com/news/global-biofuel-demand-to-grow-20-in-next-five-years/ (accessed October 11, 2024)
- 3Świa,tkiewicz S Koreleski J. Effect of dietary level of maize- and rye distiller dried grains with solubles on nutrient utilization and digesta viscosity in laying hens. J Anim Feed Sci. (2007) 16:668–77. doi: 10.22358/jafs/66824/2007 · doi ↗
- 4Batal AB Parsons CM. Effects of age on nutrient digestibility in chicks fed different diets. Poult Sci. (2002) 81:400–7. doi: 10.1093/ps/81.3.400, 11902418 · doi ↗ · pubmed ↗
- 5Ward AT Marquardt RR. The effect of saturation, chain length of pure triglycerides, and age of bird on the utilization of rye diets. Poult Sci. (1983) 62:1054–62. doi: 10.3382/ps.0621054, 6878135 · doi ↗ · pubmed ↗
- 6Yuan L Wang M Zhang X Wang Z. Effects of protease and non-starch polysaccharide enzyme on performance, digestive function, activity and gene expression of endogenous enzyme of broilers. P Lo S One. (2017) 12:1–13. doi: 10.1371/journal.pone.0173941, 28323908 PMC 5360255 · doi ↗ · pubmed ↗
- 7Kumar R Tiwari RK Kumari A Shahi B Singh KM Saha SK . Effect of supplementation of non-starch polysaccharide cocktail enzyme on performance in broiler. J Agri Search. (2019) 6:95–100.
- 8Yaghobfar A Kalantar M. Effect of non–starch polysaccharide (NSP) of wheat and barley supplemented with exogenous enzyme blend on growth performance, gut microbial, pancreatic enzyme activities, expression of glucose transporter (SGLT 1) and MUCIN producer (MUC 2) genes of broiler. Rev Bras Cienc Avic / Brazilian J Poult Sci. (2017) 19:629–38. doi: 10.1590/1806-9061-2016-0441 · doi ↗