Tannic acid shaped microbiome composition in midguts and rearing microcosms of Aedes triseriatus (Say)
Shicheng Chen, Liang Cui, Bin Zuo, Jiangchao Zhao, Edward D. Walker

TL;DR
Tannic acid affects the gut microbiome of Aedes triseriatus mosquitoes and their rearing environment, potentially offering new biocontrol strategies.
Contribution
This study reveals how tannic acid alters mosquito gut microbiota and microbial community structures in rearing habitats.
Findings
TA treatment reduced microbial richness and diversity in mosquito guts.
TA increased Pseudomonadota abundance while decreasing Bacteroidota.
TA exposure enriched pathways related to carbohydrate and amino acid metabolism.
Abstract
Tannic acid (TA), a polyphenol derived from plants, often accumulates in water-holding containers where mosquitoes develop. Yet, its effects on mosquito gut microbiota remain poorly understood, representing an important knowledge gap. Because mosquito-associated microbiota are vital for host development, nutrition, and immunity, uncovering how TA shapes these microbial communities may yield new insights into mosquito biology and vector control strategies. In this study, we conducted a comparative analysis of bacterial communities in Aedes triseriatus midguts and rearing microcosms with or without TA supplement. Addition of TA at 0.35 mg/mL caused up to 50% larval Ae. triseriatus mortality, whereas combined supplementation with TA and kanamycin (100 μg/mL) increased mortality to 75% relative to controls. TA treatment significantly reduced microbial Chao 1 richness and Shannon diversity…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
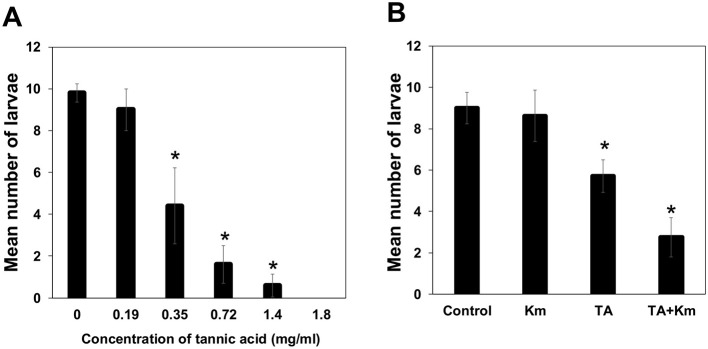 Figure 1
Figure 1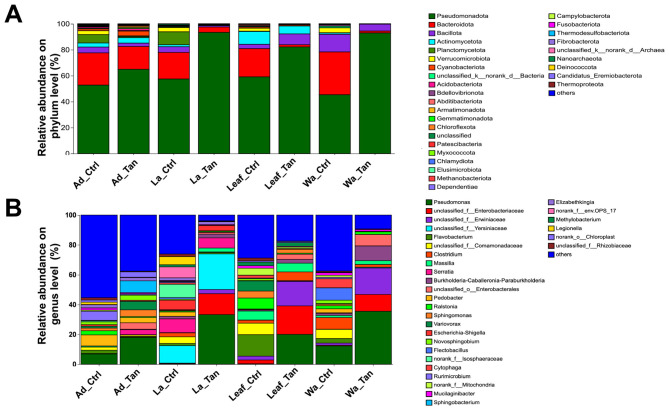 Figure 2
Figure 2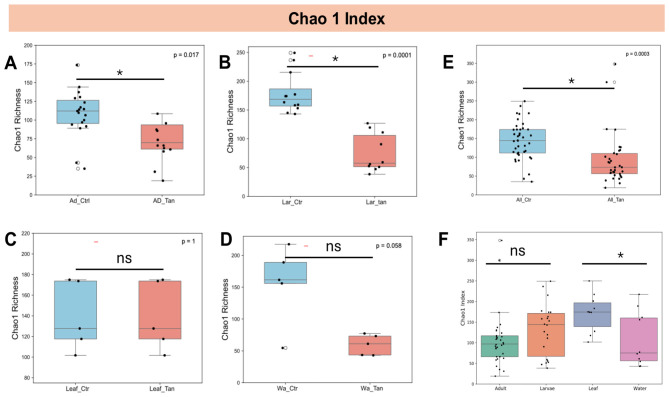 Figure 3
Figure 3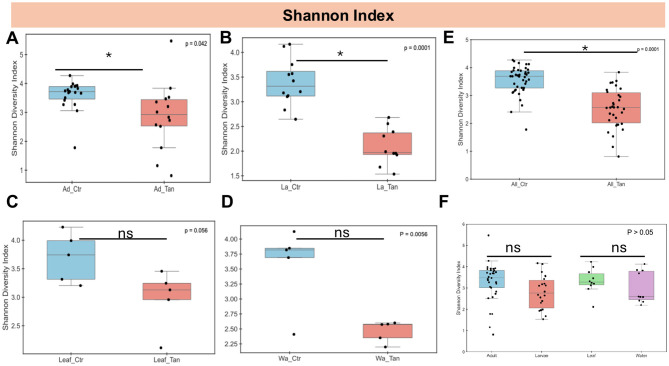 Figure 4
Figure 4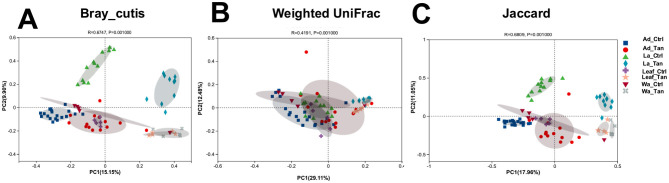 Figure 5
Figure 5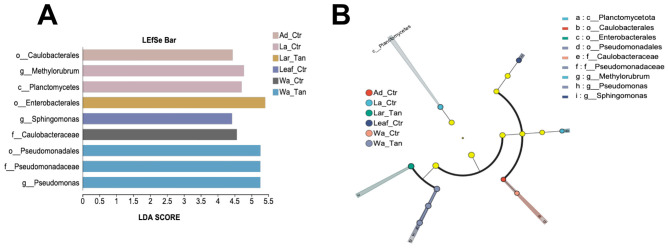 Figure 6
Figure 6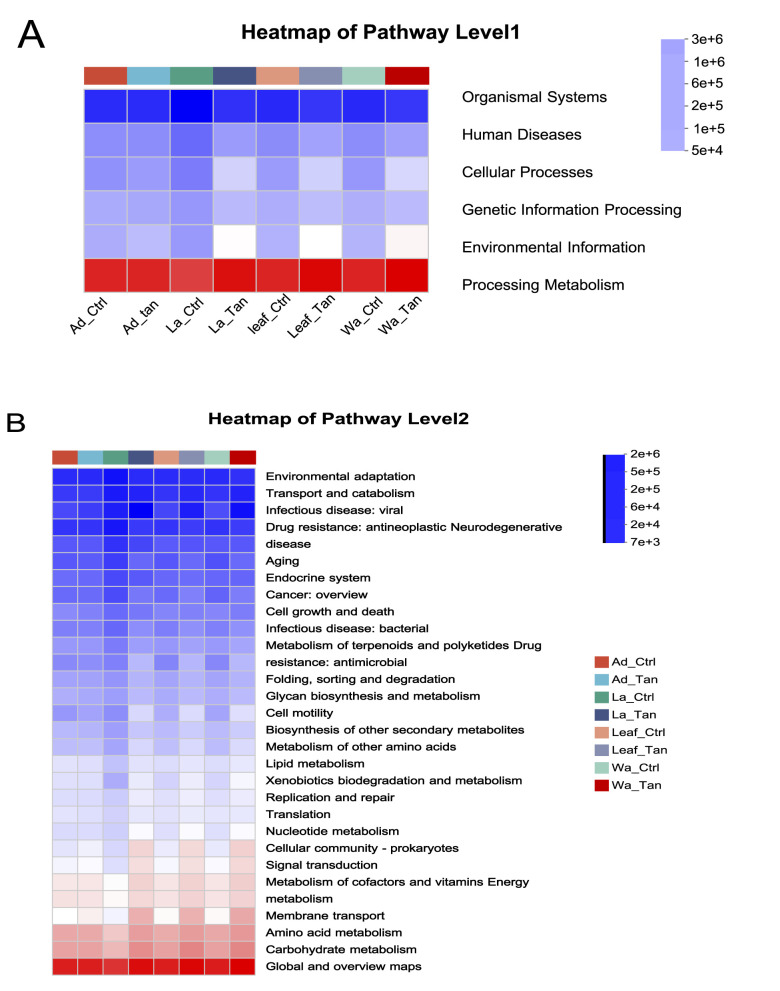 Figure 7
Figure 7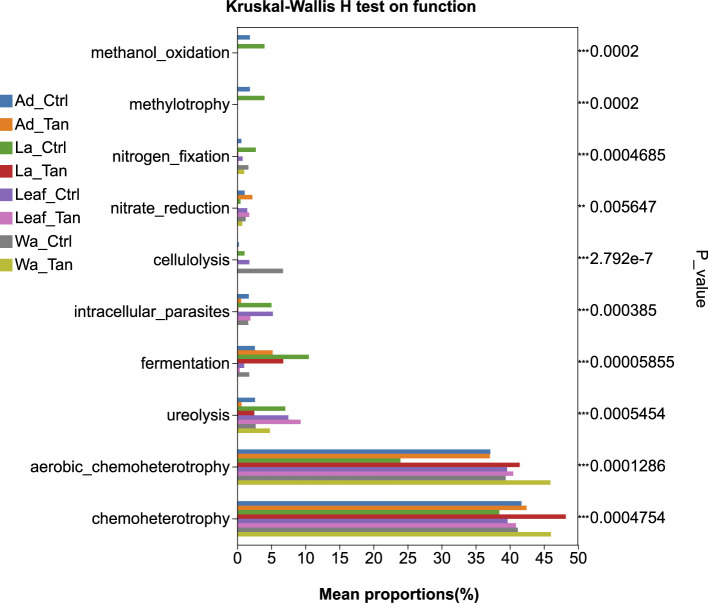 Figure 8
Figure 8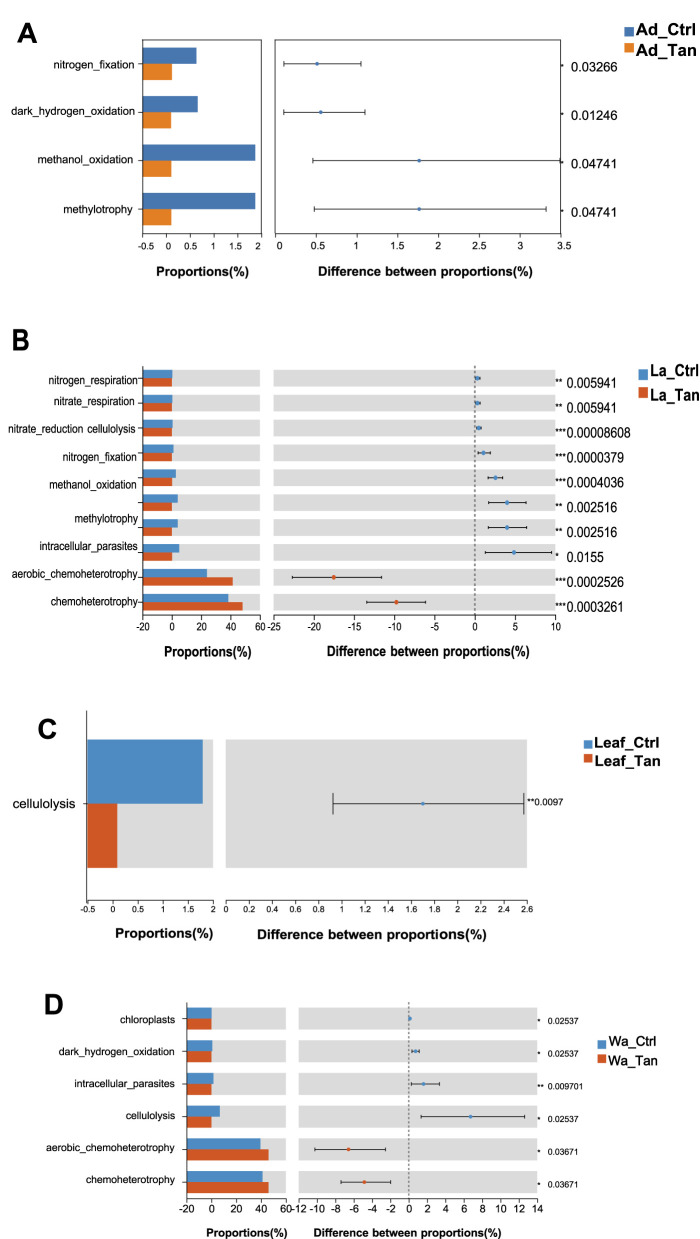 Figure 9
Figure 9|
|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|
| Water*leaf | Microcosm | 1 | 0.70 | 0.69 | 2.20 | 0.11 | 0.028 |
| Residuals | 18 | 5.66 | 0.31 | 0.89 | |||
| Total | 19 | 6.35 | |||||
| TA*control | Treatment | 1 | 0.64 | 3.05 | 8.05 | 0.01 | 0.001 |
| Residuals | 73 | 27.68 | 0.34 | 0.9 | |||
| Total | 74 | 30.73 | 1 | ||||
| Adults*lavae | Development | 1 | 3.22 | 3.32 | 9.08 | 0.15 | 0.001 |
| Residuals | 1 | 18.77 | 0.35 | 0.85 | |||
| Total | 54 | 21.99 | 1 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · Invertebrate Immune Response Mechanisms · Insect Utilization and Effects
Introduction
1
Aedes triseriatus (Say) (Diptera: Culicidae), the primary vector of La Crosse virus in the Midwestern United States, breeds in natural habitats such as water-filled tree holes, as well as artificial containers like discarded tires (Grimstad and Walker, 1991). These microhabitats are sustained by the accumulation of senescent leaves, plant detritus, and dead invertebrates, which drive microbial activity and support the base of the aquatic food web (Fish and Carpenter, 1982; Walker et al., 1991). Although the water in these habitats is regularly replenished by rainwater moving into tree holes as stemflow (Walker et al., 1991), the decomposition of leaf litter through chemical and biological processes alters its nutritional quality for mosquito larvae and other aquatic invertebrates (Walker et al., 1991). Microbial metabolism, in particular, modifies water chemistry and releases various compounds, including TA, which can accumulate to toxic levels and negatively impact mosquito larval development (Rey et al., 2000). Despite its ecological relevance, the physiological and behavioral effects of leaf-derived compounds such as TA on mosquito larvae remain poorly understood. Tannins in tree hole water inhibited growth of the western tree hole mosquito, Ae. sierrensis and caused larval mortality in a dose dependent manner (Mercer, 1993; Mercer and Anderson, 1994). Histological analyses have shown that midgut epithelium was damaged by TA or its derivatives, causing cellular degeneration, in particular affecting the clear cells of the anterior midgut before the dark cells of the posterior region (Rey et al., 1999). Additionally, TA has been reported to interfere with the digestion of starches, lipids, and proteins, thereby disrupting nutrient absorption and leading to growth inhibition, as observed in Hyphantria cunea larvae (Zhang et al., 2023a).
TA, a plant-derived polyphenol, exhibits broad-spectrum antimicrobial and anti-biofilm activities against various bacteria and fungi in vitro. It inhibited growth of both Gram-positive and Gram-negative bacteria by disrupting cell membranes, chelating essential metal ions, and interfering with enzymatic functions (Lorca et al., 2024; Leitão et al., 2025). TA also effectively prevented biofilm formation by reducing microbial adhesion to surfaces, disrupting cell-to-cell signaling, and destabilizing extracellular polymeric substances (Hosseini et al., 2025; Zheng et al., 2025). These properties were observed across diverse microbial taxa, including Streptococcus, Bacillus, and fungal genera such as Candida (Payne et al., 2013; Long et al., 2024; Moreira et al., 2024; Xu et al., 2025), underscoring its ecological significance and potential applications in microbial community modulation and biofilm management (Govindarajan et al., 2025). In vivo studies in insects demonstrated that TA disrupted gut microbial communities and interfered with digestive processes, contributing to reduced growth and survival (Tan et al., 2022). For example, in Hyphantria cunea larvae, TA ingestion caused significant damage to midgut epithelial cells and impaired the activities of key digestive enzymes, including amylase, lipase, and protease, resulting in poor nutrient absorption and growth retardation (Tan et al., 2022; Zhang et al., 2023a). Additionally, TA increased gut permeability in insects, potentially facilitating microbial translocation and immune activation (Choi et al., 2022). These findings highlighted the dual role of TA in directly affecting insect physiology and indirectly influencing development through microbiota modulation (Tan et al., 2022). Given that mosquito larvae depend heavily on their gut microbiota for digestion and development, elevated levels of TA possibly disrupt these microbial communities (either within the gut or the larval habitat), thus impeding larval development (Coon et al., 2014). Furthermore, midgut epithelial damage may compromise the gut barrier, allowing microbial translocation into the hemocoel and potentially leading to sepsis. This hypothesis is supported by studies showing enhanced larvicidal efficacy of Bacillus thuringiensis when co-administered with TA (Isayama et al., 2021).
Several bacteria possess the ability to detoxify TA through enzymatic degradation pathways, enabling them to survive in tannin-rich environments such as leaf litter or herbivore insect guts (Zimmer, 1999; Barbehenn and Peter Constabel, 2011). Key enzymes include tannase, which hydrolyzes TA into gallic acid and glucose. Gallate decarboxylase then further metabolizes gallic acid (Lekshmi et al., 2021). Microbes such as Lactobacillus plantarum, Bacillus subtilis, Enterococcus faecalis, and Streptococcus gallolyticus have been shown to express these enzymes and tolerate TA exposure (Goel et al., 2011; Jiménez et al., 2013, 2014; Govindarajan et al., 2024). This microbial detoxification not only benefits the bacteria themselves but may also alleviate tannin toxicity for host organisms, such as insects, by reducing oxidative stress and supporting gut homeostasis (Barbehenn and Peter Constabel, 2011; Lekshmi et al., 2021; Tan et al., 2022; Govindarajan et al., 2025). In this study, we examined how varying concentrations of TA influenced larval mosquito survival and found that the removal of microbial communities further reduced larval survivorship, suggesting a protective role of the microbiota. We conducted high-throughput amplicon sequencing of the 16S rRNA gene to characterize shifts in microbial communities in both larval habitats and mosquito midguts following TA exposure. Our results support the potential application of TA as a standalone or synergistic agent in mosquito control strategies. Furthermore, a deeper understanding of detoxification mechanisms in Aedes mosquitoes and their associated microbiota may reveal novel targets for disrupting vector populations and reducing disease transmission.
Materials and methods
2
Mosquitoes and experimental design
2.1
Aedes triseriatus mosquito larvae were from a laboratory colony originating from larvae collected from bark-lined, pan tree holes in American beech (Fagus grandifolia Ehr.) on the Michigan State University campus and maintained under standard insectary conditions (27 °C, 12:12 h light:dark cycle, and 85% relative humidity) (Walker et al., 1997). Upon adult emergence, mosquitoes were provided with a 10% sucrose solution ad libitum via a cotton wick. Adult Ae. triseriatus were blood-fed using bovine blood (Hemostat Lab, Dixon, CA, USA) delivered through an artificial membrane feeder. Two days post-blood meal, oviposition substrates (filter paper moistened with water and placed in Petri dishes) were introduced into rearing cages. Collected eggs were transferred to plastic containers filled with distilled water for hatching. First instar larvae were fed Tetramin tropical fish food flakes (Tetra, Blacksburg, VA, USA) ad libitum, followed by daily ad libitum feedings of Purina Cat Chow (Nestlé) for subsequent instars.
To evaluate the effects of TA (Sigma-Aldrich, St Louis, MO) on mosquito development and microbiota, experimental microcosms were prepared with 1 g of senescent, American beech leaf litter sampled in autumn, dried, and placed into 80 mL of Milli-Q water, and 1 ml of microbial inoculum, consistent with our previous experiments (Walker et al., 1997; Norman and Walker, 2018). Immersed leaves after 6 days of conditioning allow for microbial colonization (Norman and Walker, 2018, 2019). To evaluate the dose-dependent effect of TA (Supplementary Figure 1), larvae were hatched from eggs and seeded into microcosms; 10 s instar larvae were exposed to a range of supplemented TA concentrations (0.00, 0.19, 0.35, 0.72, 1.4, and 1.8 mg/mL). The number of surviving larvae was counted after 1 week.
The effects of TA and kanamycin on Ae. triseriatus larval survival were assessed by counting surviving larvae 1 week after treatment (Kuiters and Sarink, 1986). Kanamycin, a water-soluble broad-spectrum antibiotic, was used to reduce the bacterial load in larval mosquitoes (data not shown). For these combinatorial treatments (Supplementary Figure 2), four groups were established: (1) kanamycin-only group (100 μg/mL), (2) TA-only group (sublethal dose, 0.35 mg/mL), (3) TA + kanamycin group (0.35 mg/mL TA + 100 μg/mL kanamycin), and (4) untreated control group. Microcosms received either 2 mL TA solution plus 2 mL Milli-Q water (treatment) or 4 mL Milli-Q water only (control). Ten second instar Ae. triseriatus larvae were introduced into each microcosm at the time of treatment. Larval survival rate was recorded after 7 days of exposure. The experiment was replicated five times, and the data were analyzed using one-way analysis of variance (ANOVA). Significance was defined as p < 0.05. Tukey's HSD was used for post-hoc pairwise comparisons.
To perform microbial community analysis, the microcosms were set up as described above. Fourth-instar larvae were collected for sampling to obtain enough midgut contents after a ten-day incubation (Supplementary Figure 3). Upon adult emergence, mosquitoes were caught, and were surface-sterilized by three rinses in 70% ethanol followed by one rinse in sterile water. Midguts were removed under sterile conditions using finely sharpened, number 5 watchmaker's forceps (Daigger Scientific, New Jersey, USA), placed into 200 μL of sterile phosphate-buffered saline (PBS), and pooled from six individuals per sample. For larvae, at least 12 replicates were analyzed in the TA-treated group and 10 in the control group (Supplementary Figure 3). For adults, 14 TA-treated and 19 control samples were processed. Each pooled set was homogenized with a sterile pestle. Water samples were collected by centrifuging at 15,000 rpm for 10 min at 4 °C; resulting pellets were stored at −70 °C until further use. Leaf materials after removing surface water were flash-frozen in liquid nitrogen, and ground with a sterile pestle. For water and leaf samples, five replicates were included for each treatment group (Supplementary Figure 3).
DNA extraction, library construction, and 16S rRNA sequencing
2.2
All dissections and DNA extractions were conducted in a laminar flow biosafety cabinet to avoid contamination. Mosquito midgut tissues, pellets from water and leaf materials were resuspended in 200 μL of lysis buffer, and DNA was extracted using the DNeasy Blood & Tissue Kit (Qiagen) according to the manufacturer's protocol. DNA concentration was quantified using Qubit™ dsDNA HS Assay Kits, and integrity was confirmed by PCR with primers 63F (CAGGCCTAACACATGCAAGTC) and 1387R (CGGAACATGTGWGGCGGG). Amplicon tagging and 16S rRNA gene sequencing were performed at the Research Technology Support Facility (RTSF) at Michigan State University. The V4 region of the bacterial 16S rRNA gene was amplified using primers 515f (GTG CCA GCM GCC GCG GTA A) and 806r (NNN NNN GGA CTA CHV GGG TWT CTA AT), with unique 6-bp error-correcting barcodes included in the reverse primer. Amplicons were purified, pooled, and sequenced using the Illumina MiSeq platform with a 2 × 250 bp paired-end format and 500-cycle v2 reagent kit. Base calling was performed with Illumina Real Time Analysis (RTA) v1.18.54, and output files were demultiplexed and converted to FASTQ format using Illumina Bcl2fastq v1.8.4.2.3.
Bioinformatics analysis and statistical analysis
2.3
To obtain high-quality sequences, bases at the head or tail with Phred quality scores below Q30 were trimmed, and sequences shorter than 100 bp were discarded. Raw sequence data were processed in QIIME2 (version 2024.2) (Bolyen et al., 2019), including sequencing and PCR error reduction, and denoising with DADA2 (filtering, dereplication, chimera removal, and merging of paired-end reads) to generate amplicon sequence variants (ASVs) for taxonomic analysis. ASVs representing less than 0.1% of the total sequences across all samples or annotated as chloroplast or mitochondrial contaminants were removed. Sequencing depth was normalized to the minimum read count among samples. Taxonomic classification was performed using a Naive Bayes classifier trained on the SILVA 138.2/16S rRNA database (https://www.arb-silva.de/). Alpha diversity was calculated in Mothur (version 1.30.2) (https://mothur.org/) using the Chao1 richness and Shannon diversity indices based on randomly subsampled sequences from each sample. Statistical differences in alpha diversity were assessed using the Kruskal–Wallis test. Differences were considered statistically significant when the p-value was less than 0.05. Beta diversity was assessed using PERMANOVA (999 permutations) based on Bray–Curtis distances, weighted UniFrac, and Jaccard dissimilarities, implemented in the vegan package (version 2.4-1) in R (version 3.3.1). Microbial biomarkers for each experimental group were identified using LEfSe (version 1.0) (Segata et al., 2011).
Microbial functions were predicted using PICRUSt2 (v2.2.0-b) (Douglas et al., 2020), and functional differences between treatment groups were assessed using FAPROTAX (version 1.2.11) (Louca et al., 2016). Predicted genes were annotated against the Enzyme Commission (EC) database (https://enzyme.expasy.org/), the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (Kanehisa, 2000; Kanehisa et al., 2016, 2025), and the MetaCyc Metabolic Pathway Database (https://metacyc.org/). Group differences were analyzed in STAMP (https://beikolab.cs.dal.ca/software/STAMP), with two-group comparisons performed using the Wilcoxon test followed by false discovery rate (FDR) correction. Statistical significance was defined as p < 0.05.
Sequence data accession number
2.4
Raw paired-end reads per sample without barcode and primer bases were submitted to the Sequence Read Archive of the NCBI (PRJNA1320966).
Results
3
Effects of TA and antibiotic treatment on larval mosquito
3.1
Larval Ae. triseriatus survival declined with increasing TA concentrations, with 100% mortality observed at 1.8 mg/mL TA (Figure 1A) (p < 0.05). Larval survival was high and equivalent between controls and kanamycin alone but was lower with TA (at 35 mg/mL) and lowest with TA and kanamycin in combination (Figure 1B) (p < 0.05), indicating a synergistic negative effect on larval survivorship.
Impact of TA and antibiotic treatment on mosquito larval survival. (A) Survival of Ae. triseriatus larvae decreased with increasing TA concentrations. Larvae at the 2nd instar stage were exposed to TA, and surviving individuals were counted after 1 week. (B) Km treatment reduced microbial protection against TA toxicity. Larvae at the 2nd instar stage were reared under four conditions: control (no TA or antibiotics), TA alone (0.35 mg/ml), Km alone (100 μg/ml), and TA + Km. Km alone had no effect on survival. Statistical analysis was performed using one-way ANOVA, with Tukey's HSD for post-hoc pairwise comparisons; an asterisk () indicates a statistically significant difference at p < 0.05.*
Effects of TA treatment on bacterial composition in insect guts and microcosms
3.2
After filtering, denoising, merging, and chimera removal, 3,313,743 high-quality reads were obtained, with a median depth of 44,183 reads per sample. Taxonomic profiling assigned sequences to at least 38 bacterial phyla. The five most abundant were Pseudomonadota, Bacteroidota, Bacillota, Planctomycetota, and Actinomycetota (Figure 2A). Across all habitats (rearing water, leaf surfaces, larval midguts, and adult midguts) Pseudomonadota predominated (45.3–93.3%), regardless of TA treatment. Bacteroidota ranked second (1.61–33.4%). In rearing water, TA addition remarkably increased the relative abundance of Pseudomonadota (from 45.3% to 92.4%) while sharply reducing Bacteroidota (from 33.4% to 1.6%), indicating strong TA-mediated inhibition of this bacterial group. Bacillota abundance declined in TA-treated water (from 12.7% to 5.8%), Verrucomicrobia nearly disappeared (from 4.1% to 0), and Actinomycetota decreased (from 0.9% to 0.2%). Similar patterns were observed on leaves and in midguts, except Bacillota increased on TA-treated leaves (from 3.0% to 7.9%). Planctomycetota were more common in larvae (10.2%) and adults (5.9%) from non-TA treatments but remained rare in water (0.3%) and leaves (0.3%).
Relative abundance of bacterial communities in microcosms and mosquito midguts with and without TA treatment. (A) Bacterial community composition at the phylum level. (B) Community composition at the genus level. C indicates control (no TA) and T indicates TA treatment. Ad_Ctr, adult mosquitoes without TA; Ad_Tan, adult mosquitoes with TA; La_Ctr, larval mosquitoes without TA; Lar_Tan, larval mosquitoes with TA; Leaf_Ctr, leaf without TA; Leaf_Tan, leaf with TA; Wa_Ctr, water without TA; Wa_Tan, water with TA.
Genus-level composition varied widely between treatments and sample types (Figure 2B). In control water, ~37.5% of reads were unclassified at the genus level (“others”); dominant genera included Pseudomonas, Flectobacillus, Clostridium, unclassified Comamonadaceae, and Cytophaga. TA-treated water showed a marked shift. Pseudomonas nearly doubled in abundance (35.6%) while Flectobacillus, Clostridium, and Cytophaga declined to negligible levels (Figure 2B). Conversely, unclassified Erwinaceae, unclassified Enterobacteriaceae, Burkholderia–Caballeronia–Paraburkholderia, and unclassified Enterobacteriales were enriched (Figure 2B). In control leaves, Pseudomonas was rare (0.4%); dominant genera included Flavobacterium, unclassified Comamonadaceae, unclassified Enterobacteriales, Ralstonia, and Variovorax. TA-treated leaves were dominated by Pseudomonas, unclassified Erwinaceae, and unclassified Enterobacteriaceae, resembling TA-treated water communities. In control larval midguts, 55.4% of reads were unclassified (“others”), with Pedobacter, Pseudomonas, Rurimicrobium, Elizabethkingia, and Ralstonia as key taxa. TA treatment doubled Pseudomonas abundance, while Elizabethkingia and Pedobacter remained similar to controls; Sphingobacterium, Serratia, and Variovorax were also common (Figure 2B). Adult midguts in controls were dominated by unclassified Yersiniaceae, Serratia, unclassified Isosphaeraceae, unclassified env.OPS_17, Escherichia–Shigella, and Legionella. In TA-treated adults, Pseudomonas, unclassified Enterobacteriales, and unclassified Yersiniaceae increased markedly (33.3%, 23.8%, and 14.0%, respectively).
Alpha and beta diversity affected by TA in the insect guts and microcosms
3.3
Building on our initial observations of microbial community composition, we next assessed whether alpha diversity was influenced by TA treatment (Figure 3). Bacterial richness, as measured by the Chao1 index, was significantly reduced in TA-treated adult guts compared to controls (p < 0.05) (Figure 3A). A similar decrease in richness was observed in larval samples following TA exposure (Figure 3B). In contrast, no significant richness differences were detected between TA-treated and untreated leaf samples (p > 0.05) (Figure 3C), nor in water samples (Figure 3D). However, when comparing all non-TA-treated samples (water, leaf, larvae, and adult grouped as controls) against all TA-treated samples (Figure 3E), a significant difference emerged (p < 0.05). Additionally, leaf samples generally exhibited higher Chao1 values than water samples, independent of treatment. When TA treatment was disregarded, no significant differences in richness were found between treated and untreated adults or larvae (p > 0.05) (Figure 3F).
Chao1 richness of gut microbiota across Aedes mosquito developmental stages and environmental microcosms. (A) Adults with and without TA treatment. (B) Larvae with and without TA treatment. (C) Leaf samples with and without TA treatment. (D) Water samples with and without TA treatment. (E) Combined samples (adults, larvae, leaves, water) with and without TA treatment. (F) Comparison across adults, larvae, leaves, and water, regardless of TA treatment. Diversity metrics are based on total ASVs. Statistical differences were assessed using the Kruskal–Wallis test. * indicates p < 0.05; ns, not significant (p > 0.05). Ad_Ctr, adult mosquitoes without TA; Ad_Tan, adult mosquitoes with TA; La_Ctr, larval mosquitoes without TA; Lar_Tan, larval mosquitoes with TA; Leaf_Ctr, leaves without TA; Leaf_Tan, leaves with TA; Wa_Ctr, water without TA; Wa_Tan, water with TA.
Significant differences in species diversity, assessed via the Shannon index, were found between non-TA-treated controls and TA-treated adults (p < 0.05), with TA-treated larvae also showing reduced diversity relative to controls (Figures 4A, B). However, no significant diversity differences (p > 0.05) were observed between TA-treated and control leaves, nor between TA-treated and control water samples (Figures 4C, D). When ignoring TA treatment, Shannon diversity did not differ significantly (p > 0.05) among adult and larval mosquitoes or between leaf and water samples (Figures 4E, F).
Shannon diversity index of gut microbiota across Aedes mosquito developmental stages and environmental microcosms. (A) Adults with and without TA treatment. (B) Larvae with and without TA treatment. (C) Leaf surfaces with and without TA treatment. (D) Water samples with and without TA treatment. (E) Combined samples (adults, larvae, leaves, water) with and without TA treatment. (F) Comparison across adults, larvae, leaves, and water, regardless of TA treatment. Diversity metrics are based on total ASVs. Statistical differences were assessed using the Kruskal–Wallis test. * indicates p < 0.05; ns, not significant (p > 0.05). Ad_Ctr, adult mosquitoes without TA; Ad_Tan, adult mosquitoes with TA; La_Ctr, larval mosquitoes without TA; Lar_Tan, larval mosquitoes with TA; Leaf_Ctr, leaves without TA; Leaf_Tan, leaves with TA; Wa_Ctr, water without TA; Wa_Tan, water with TA.
For beta diversity analysis, we performed PCoA to assess similarities in microbial structures among TA treatment, rearing systems and development stages. PCoA analyses were based on Bray-Curtis distance, Weighted UniFrac and Jaccard, respectively (Figure 5). For Bray-Curtis distance analysis (Figure 5A), the dissimilarity revealed clear separation between TA-treated and control microbial communities (ANOSIM: R = 0.67, p = 0.001). The first two PCoA axes explained 25.05% of the total variance (PCoA1: 15.15%, PCoA2: 9.90%). Samples clustered primarily by TA treatment, with separation along axis 1 except for adult mosquito samples. Moreover, both Weighted UniFrac (ANOSIM: R = 0.42, p = 0.001) and Jaccard (ANOSIM: R = 0.68, p = 0.001) distance analyses supported significant structural differences between TA-treated and control microbiome (Figures 5B, C), agreeing with Bray-Curtis analysis. From the three different dissimilarity analyses, we also observed there were distinct gut microbial communities across developmental stages (larvae vs. adults) and in rearing systems (leaf vs. water) (Figure 5). Moreover, PERMANOVA analysis indicated that TA treatment (p = 0.001), rearing systems (p = 0.028), and developmental stages (p = 0.001) were all significant drivers of beta diversity (Table 1).
Principal coordinate analysis (PCoA) of bacterial community structure. (A) Bray–Curtis distances. (B) Weighted UniFrac distances. (C) Jaccard distances. Distances between symbols on each ordination plot indicate relative dissimilarities in community composition. Ad_Ctr, adult mosquitoes without TA; Ad_Tan, adult mosquitoes with TA; La_Ctr, larval mosquitoes without TA; Lar_Tan, larval mosquitoes with TA; Leaf_Ctr, leaves without TA; Leaf_Tan, leaves with TA; Wa_Ctr, water without TA; Wa_Tan, water with TA.
Linear discriminant analysis effect size (LEfSe) analysis
3.4
LEfSe analysis identified bacterial taxa differentially enriched across mosquito life stages and microcosms, spanning from phylum to genus level (Figures 6A, B). Only taxa with an LDA score greater than 2.0 were included. Three candidate markers in TA-treated water were found (Figure 6A): order Pseudomonadales, family Pseudomonadaceae, and genus Pseudomonas. In contrast, family Caulobacteraceae was enriched exclusively in control water. Two markers such as genus Methylorubrum and class Planctomycetes were enriched in control larvae, while genus Sphingomonas was a marker for control leaf. Additionally, family Caulobacterales served as a characteristic marker in control adults. No marker taxa were detected in treated leaf or treated adult groups. The cladogram from phylum to species was used to further understand the distribution of these different taxa at different taxonomic levels (Figure 6B). The differences in phylogenetic distribution showed that the microbial communities in each sample group are compositionally distinct.
LEfSe analysis showing significant differences of microbial species from phylum to genus. (A) LEfSe Bar diagram for different samples with LDA scores higher than 2.0. A higher LDA score represents that this bacterial taxon has a greater contribution to the differences. (B) Cladogram indicates the phylogenetic distribution of microbial communities in insect guts and rearing system and their TA-treated samples. Yellow nodes represent microbial taxa with no significant difference between different life stages, while other color nodes represent microbial taxa that are significantly enriched at those life stages. Ad_Ctr, adult mosquitoes without TA treatment; Ad_Tan, adult mosquitoes with TA treatment; La_Ctr, larval mosquitoes without TA treatment; Lar_Tan, larval mosquitoes with TA treatment; Leaf_Ctr, leaf without TA treatment; Leaf_Tan, leaf with TA treatment; Wa_Ctr, water without TA treatment; Wa_Tan, water with TA treatment.
Functional prediction analysis reveals potential differences in microbial community metabolic function
3.5
PICRUSt2 predicted six major functional pathways (Level 1) based on KEGG (Kanehisa, 2000; Kanehisa et al., 2016, 2025) (Figure 7A). These functional categories were enriched in metabolism, environmental information processing, genetic information processing, cellular processes, human diseases, and organismal systems (Figure 7A). Among all groups, metabolism was the predominant predicted function (72.91–76.5%), followed by environmental information processing (6.07–9.55%) and genetic information processing (4.80–6.50%). It is noted that more “environmental information processing” functions were predicted in the TA-treated samples. The most abundant pathways at Level 2 included carbohydrate metabolism, global and overview maps, amino acid metabolism, energy metabolism, and metabolism of cofactors and vitamins (Figure 7B). However, COG functional classification did not clearly resolve the predicted functional differences (Supplementary Figure 4). Therefore, FAPROTAX was employed to refine functional predictions, focusing on processes such as plant cell wall degradation, nitrogen and carbon cycling, and nutrient assimilation, based on individual microbial profiles. This annotation identified 30 functional categories within the insect guts and rearing systems (Supplementary Figure 5). The top five enriched pathways were chemoheterotrophy, aerobic chemoheterotrophy, fermentation, and ureolysis (Supplementary Figure 5). We further analyzed the different predicted function enriched in treatment for further comparison (Figures 8, 9). Notably, cellulolysis and intracellular parasite-related functions were more abundant in water controls without TA compared to TA-treated water (Figure 8). The only significantly different pathway between TA-treated and untreated leaf surface samples was cellulolytic activity (Figure 9C). It seemed that “methanol oxidation” and “methylotrophy” were more abundant in larval and adult mosquitoes without TA, which likely benefits insect physiology (Figures 9A, B). Genes involved in nitrogen metabolism were commonly detected in both larval and adult mosquitoes (Figures 9A, B). Overall, TA-treated insects showed a reduced proportion of nitrogen fixation and methanol metabolism genes. However, several functions such as aerobic chemoheterotrophy showed higher abundance in TA-treated water and larval samples (Figures 9B, D).
Prediction of KEGG functions of bacteria at different developmental stages, with TA-treatment and in the microcosms. (A) Level 1 of predicted function categories is shown in each group. (B) Level 2 of predicted function categories is shown in each group. Ad_Ctr, adult mosquitoes without TA treatment; Ad_Tan, adult mosquitoes with TA treatment; La_Ctr, larval mosquitoes without TA treatment; Lar_Tan, larval mosquitoes with TA treatment; Leaf_Ctr, leaf without TA treatment; Leaf_Tan, leaf with TA treatment; Wa_Ctr, water without TA treatment; Wa_Tan, water with TA treatment.
Functional differences among all groups. The relative proportions of FAPROTAX-predicted functions were compared across mosquito developmental stages, TA treatments, and microcosm samples. p-values are shown along the right Y-axis to indicate significant differences among the selected groups. Ad_Ctr, adult mosquitoes without TA treatment; Ad_Tan, adult mosquitoes with TA treatment; La_Ctr, larval mosquitoes without TA treatment; Lar_Tan, larval mosquitoes with TA treatment; Leaf_Ctr, leaf without TA treatment; Leaf_Tan, leaf with TA treatment; Wa_Ctr, water without TA treatment; Wa_Tan, water with TA treatment.
Paired comparation of Functional differences. The relative proportions of FAPROTAX-predicted functions were compared. (A) Adults with and without TA treatment. (B) Larvae with and without TA treatment. (C) Leaf surfaces with and without TA treatment. (D) Water samples with and without TA treatment. Ad_Ctr, adult mosquitoes without TA treatment; Ad_Tan, adult mosquitoes with TA treatment; La_Ctr, larval mosquitoes without TA treatment; Lar_Tan, larval mosquitoes with TA treatment; Leaf_Ctr, leaf without TA treatment; Leaf_Tan, leaf with TA treatment; Wa_Ctr, water without TA treatment; Wa_Tan, water with TA treatment.
Discussion
4
Previous research on tannin-insect interactions largely focused on their impacts on the growth and physiology of herbivorous pests, with relatively little attention paid to aquatic insects (Barbehenn and Peter Constabel, 2011; Isayama et al., 2021; Tan et al., 2022; Zhang et al., 2023a). In this study, we investigated the effects of TA on mosquito larval development, gut microbiota composition, and environmental microbial communities within experimental rearing microcosms. Our research centered on the feeding ecology of Ae. triseriatus larvae in container habitats, where early-life interactions with microbial communities can influence adult fitness and vector competence (Merritt et al., 1992; Kaufman et al., 1999; Xu et al., 2008; Chen et al., 2014). Larval Ae. triseriatus feed by filtering and browsing suspended particulate matter, detritus of plant leaves, and microbial biofilms found on submerged surfaces such as plant material and container walls (Merritt et al., 1992). In natural tree holes, microbial community composition was shaped by both leaf litter decomposition and larval mosquito activity (Paradise et al., 2008). However, little is known about how mosquito larvae tolerate or metabolize plant-derived polyphenol compounds such as tannins, which commonly accumulate in these environments (Rey et al., 1999). The western tree hole mosquito was negatively affected by exposure to tannins, but any ameliorating effects of microorganisms on the effects were not investigated (Mercer, 1993; Mercer and Anderson, 1994). Overall, our findings demonstrated TA influenced both mosquito survival and the structure of their associated microbial communities. These insights expand our understanding of how ecological and chemical factors shape mosquito–microbiome interactions and suggest that naturally occurring compounds such as tannins could be leveraged to manipulate mosquito-associated microbiota as part of integrated biocontrol strategies against mosquito populations and vector-borne diseases (Isayama et al., 2021; Tan et al., 2022).
Our results showed that elevated TA concentrations significantly reduced larval survival rate. Although TA likely exerts its effects primarily by directly damaging the midgut epithelium of Aedes mosquitoes as reported previously (Mercer, 1993; Rey et al., 1999), we cannot rule out additional pathways through which TA may influence larval physiology (Tan et al., 2022; Hosseini et al., 2025). For example, TA was shown to increase reactive oxygen species while decreasing antioxidant and detoxification enzyme activities in hosts (Tan et al., 2022). In addition, dietary tannins were reported to change growth, behavior, and the gut microbiome of larval amphibians (Maurer et al., 2024). Because larval Aedes mosquitoes depend on microbial and detrital food resources that are frequently coated with plant-derived tannins, they are continuously exposed to these chemical stressors (Merritt et al., 1992; Norman and Walker, 2018, 2019). Such exposure may lower the nutritional quality of available food, disrupt gut microbial communities, and impose sublethal physiological costs that collectively slow larval growth and development. Another possible pathway for TA-induced toxicity involves changes in water chemistry, especially reductions in dissolved oxygen (DO) levels, which may result from increased microbial respiration during the breakdown of tannin-rich organic matter (Oliva et al., 2024). Soaking American beech leaves for 3 days released tannins up to 120 mg/L, substantially lowering DO (Oliva et al., 2024). Reduced DO imposed metabolic stress, increased respiration costs, and impaired larval growth and survival (Pino-Otín et al., 2023). Whether these mechanisms apply to mosquito larvae and other aquatic insects remains to be determined.
Taken together, the findings from this study and previous work suggest that tannins may influence mosquito survival through multiple interacting pathways, including direct physiological stress, reduced food quality, disruption of the gut microbiome, and alterations in water chemistry. These results highlight the potential of tannins as natural compounds for modulating mosquito populations and contributing to integrated vector management strategies. A limitation of this study is that it primarily focused on microbiome changes, without fully examining the physiological, chemical, and molecular mechanisms underlying the interactions between mosquitoes and their microbiota under high tannic acid stress. In particular, the proposed water-quality effects—such as reduced dissolved oxygen driven by increased microbial respiration during tannin degradation—remain speculative, as these parameters were not directly measured. Likewise, although previous studies indicate that tannins may elevate ROS levels and suppress antioxidant or detoxification enzyme activity, these pathways were not assessed here and therefore cannot be used to infer causality in the observed larval mortality patterns. Incorporating water-chemistry measurements, respiration assays, and physiological biomarkers of oxidative stress in future work would help clarify these potential mechanisms and strengthen the overall interpretation of tannin-mediated effects.
Exogenous toxicants are known to perturb insect gut microbiota, disrupting physiological homeostasis and impairing host growth and development (Wang et al., 2024). Consistent with this, TA exposure significantly altered the richness, diversity, and taxonomic composition of gut microbiota in both larval rearing systems and the insects themselves, with TA-treated groups exhibiting reduced Chao1 richness and lower Shannon diversity indices compared to controls. PCoA analysis revealed clear differences in microbial community structures, highlighting a strong TA-driven influence on community composition. These shifts suggested that TA exposure selectively favored microbes capable of detoxification or otherwise supporting host health under chemical stress. Despite its antimicrobial properties, many bacteria exhibit tannin resistance (Goel et al., 2011; Reyes et al., 2017; Farha et al., 2020; Rogowska-van der Molen et al., 2023). Microbes have evolved diverse mechanisms for TA degradation in natural environments (Unban et al., 2020; Rogowska-van der Molen et al., 2023). Notably, the relative abundance of Pseudomonas spp. consistently increased across insect midguts, rearing water, and leaf surfaces under sublethal TA exposure compared to controls. In agreement with this observation, Tan et al. reported a higher relative abundance of Pseudomonas in TA-treated larvae compared to untreated controls (37.43% vs. 33.59%) (Tan et al., 2022). In Pseudomonas protegens Pf-5, TA exposure modulated iron homeostasis, upregulating genes for heme uptake and siderophore biosynthesis and transport, which may reduce reactive oxygen species generation and enhance bacterial survival (Lim et al., 2013). Additionally, certain Pseudomonas species can metabolize TA and its hydrolysis product gallic acid as sole carbon sources (Chowdhury et al., 2004). TA's antimicrobial and protein-binding properties may inhibit tannin-sensitive bacteria, reducing competition and selectively enriching for tannin-tolerant taxa like Pseudomonas (Farha et al., 2020). TA may alter the microenvironment within mosquito larval guts or associated habitats (e.g., pH, nutrient availability), indirectly favoring Pseudomonas proliferation (Maurer et al., 2024). As predominant mosquito commensals, Pseudomonas spp. played key roles in shaping larval gut communities and protecting hosts from toxic metals and insecticides (Silva et al., 2024), with disruptions to these populations linked to increased larval mortality and impaired development (Tikhe and Dimopoulos, 2022). Collectively, these findings suggest that TA exposure drives a selective restructuring of mosquito gut microbiota, promoting tannin-tolerant taxa like Pseudomonas, which may provide detoxification functions and confer mutualistic benefits to the host. However, without quantifying the total bacterial load (e.g., via CFU counts or qPCR) across TA treatments and controls, it remains unclear whether Pseudomonas contributes to TA detoxification. Further studies would be required to determine the relationship between absolute abundance and intensity of detoxification of tannins.
Microbes within Burkholderia–Caballeronia-Paraburkholderia, unclassified Erwinaceae, and unclassified Enterobacteriaceae were also promoted by TA addition (Figure 2). Members of the Erwinaceae family, such as Pantoea and Erwinia, were commonly plant-associated bacteria (Walterson and Stavrinides, 2015; Wasendorf et al., 2022). Many plant-associated bacteria possessed tannin-tolerant or tannin-degrading enzymes including tannase (tannin acyl hydrolase) and polyphenol oxidases to survive in these phenolic-rich environments (Rogowska-van der Molen et al., 2023; Govindarajan et al., 2024). For instance, P. agglomerans was reported to tolerate polyphenols and degrade certain gallotannins (Xu et al., 2023). Burkholderia exhibited a dramatic increase in relative abundance in water samples treated with TA compared to controls. It was also found in Aedes mosquito habitats (Kaufman et al., 1999; Xu et al., 2008). This marked enrichment suggested a strong tolerance or adaptive response of Burkholderia to TA exposure, potentially mediated by specific genetic determinants that confer resistance or degradation capabilities. Serratia spp., another common commensal bacterium associated with mosquitoes (Chen et al., 2017), demonstrated a modest increase in relative abundance in TA-treated larval mosquitoes and persisted in adult mosquitoes. However, the physiological functions and ecological roles of Serratia in mosquito biology remain poorly characterized, warranting further investigation. The elevated abundance of Pseudomonas, Serratia, and Burkholderia collectively implies that these taxa may harbor intrinsic or acquired genes enabling resistance to the antimicrobial and protein-binding effects of TA, thereby facilitating their persistence and proliferation in TA-enriched environments (Zeida et al., 1998; Pepi et al., 2010).
In contrast, TA exposure resulted in a notable decrease in the relative abundance of Flavobacterium across both microcosms and mosquitoes (Kaufman et al., 1999; Xu et al., 2008). The reduction of Flavobacterium is particularly significant, as this genus represents an important commensal gut bacterium in mosquitoes, commonly acquired and enriched from larval habitats (Chen et al., 2014). Prior studies demonstrated that Flavobacterium played critical roles in mosquito nutrition and fitness (Chen et al., 2014); its removal delayed larval growth, reduced adult longevity, and impaired overall mosquito fitness (Giraud et al., 2022). While we observed a decrease in Flavobacterium abundance following TA exposure, we did not directly measure changes in nutritional quality or host physiology. Therefore, any potential effects of TA on mosquito nutrition or development remain hypothetical. Nonetheless, the antimicrobial activity of TA against this genus suggests that tannin presence in breeding sites could indirectly influence microbial community composition, which may, in turn, have consequences for host health and development. Investigating these dynamics further is important for understanding the complex interactions between environmental chemicals, microbiota, and mosquito physiology.
LEfSe analysis identified Pseudomonadaceae and the genus Pseudomonas as markers enriched in TA-treated water, highlighting their role in TA degradation or tolerance. In contrast, Caulobacteraceae were enriched only in non-treated water. It features a distinctive life cycle with motile swarmer and sessile stalked cells, enabling effective colonization of aquatic surfaces (Abraham et al., 2014). Widely distributed in various habitats, Caulobacteraceae contribute to nutrient cycling and organic matter decomposition (Abraham et al., 2014; Dasgupta et al., 2024), particularly in nutrient-poor environments such as water in native tree hole or disposed tires. In control larvae, two additional markers appeared: genus Methylorubrum and class Planctomycetes. Bacteria Methylorubrum are facultative methylotrophs that utilize single-carbon compounds like methanol, supporting carbon cycling and potentially host nutrition (Zhang et al., 2023b). Planctomycetes are involved in nitrogen and carbon cycling and may help maintain gut microbial balance (Storesund et al., 2020). Sphingomonas species (Showering et al., 2022), common in various insect microbiomes including mosquitoes, were markers in TA-treated larvae. Known for degrading aromatic compounds, Sphingomonas likely aid in detoxifying tannin-rich environments (Zhao et al., 2017). They were also found in herbivorous insects, assisting in plant material digestion (Pecourt et al., 2025). Lastly, Caulobacteraceae was a marker in leaf-reared control adults. LEfSe analysis did not discover significant marker taxa in two groups such as TA-treated leaf or treated adult groups. This usually means that those groups do not have bacterial taxa with strong enough differential abundance (LDA score > 2.0) compared to others, or the microbial communities are more similar in these groups.
Functional predictions using PICRUSt2 revealed that carbohydrate and amino acid metabolism dominated microbial activity in mosquito guts and their habitats, while FAPROTAX identified key functions including chemoheterotrophy and fermentation, essential for nutrient cycling and organic matter breakdown. Targeted experiments could be conducted to validate the functional predictions generated by PICRUSt. For example, targeted metabolomics could be used to confirm predicted metabolic activities, and key Pseudomonas strains could be isolated to experimentally assess their functional roles in the mosquito gut. These approaches would provide direct evidence linking microbial community composition to metabolic function and host interactions. Moreover, cellulolytic activity was higher in untreated water and leaf samples, suggesting that TA inhibits microbes responsible for plant polymer and polyphenol degradation (Makarewicz et al., 2021). This shift likely alters microbial community composition and reduces the capacity to process TA. Methanol oxidation and methylotrophy pathways were more abundant in untreated larvae and adults but reduced under TA treatment, potentially impacting host nutrition and metabolite production beneficial for mosquito development. Conversely, aerobic chemoheterotrophy increased in TA-treated water and larvae, indicating enrichment of microbes capable of tolerating or metabolizing TA. Nitrogen metabolism genes were present across samples, but nitrogen fixation decreased in TA-treated insects, potentially limiting nitrogen availability critical for larval growth. Overall, TA reshaped mosquito-associated microbial functions by suppressing specific degradation pathways while promoting others, thereby influencing microbial detoxification capacity and nutrient support, which likely affected mosquito development and survival in tannin-rich environments. Our results highlight that detoxification processes in Aedes mosquitoes and their microbiota are still poorly understood, which constrains the mechanistic interpretation of TA-induced effects. Future studies incorporating microbial functional assays, host detoxification enzyme profiling, and oxidative stress measurements will be critical to delineating how TA shapes mosquito physiology and survival.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abraham W.-R. Rohde M. Bennasar A. (2014). “The Family Caulobacteraceae,” in The Prokaryotes (Berlin, Heidelberg: Springer Berlin Heidelberg), 179–205.
- 2Barbehenn R. V. Peter Constabel C. (2011). Tannins in plant–herbivore interactions. Phytochemistry 72, 1551–1565. doi: 10.1016/j.phytochem.2011.01.04021354580 · doi ↗ · pubmed ↗
- 3Bolyen E. Rideout J. R. Dillon M. R. Bokulich N. A. Abnet C. C. Al-Ghalith G. A. . (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s 41587-019-0209-931341288 PMC 7015180 · doi ↗ · pubmed ↗
- 4Chen S. Blom J. Walker E. D. (2017). Genomic, physiologic, and symbiotic characterization of Serratia marcescens strains isolated from the mosquito Anopheles stephensi. Front. Microbiol. 8:1483. doi: 10.3389/fmicb.2017.0148328861046 PMC 5561391 · doi ↗ · pubmed ↗
- 5Chen S. Kaufman M. G. Korir M. L. Walker E. D. (2014). Ingestibility, digestibility, and engineered biological control potential of Flavobacterium hibernum, isolated from larval mosquito habitats. Appl. Environ. Microbiol. 80, 1150–1158. doi: 10.1128/AEM.03319-1324296502 PMC 3911232 · doi ↗ · pubmed ↗
- 6Choi J. Yadav S. Wang J. Lorentz B. J. Lourenco J. M. Callaway T. R. . (2022). Effects of supplemental tannic acid on growth performance, gut health, microbiota, and fat accumulation and optimal dosages of tannic acid in broilers. Front. Physiol. 13:912797. doi: 10.3389/fphys.2022.91279736117708 PMC 9478478 · doi ↗ · pubmed ↗
- 7Chowdhury S. P. Khanna S. Verma S. C. Tripathi A. K. (2004). Molecular diversity of tannic acid degrading bacteria isolated from tannery soil. J. Appl. Microbiol. 97, 1210–1219. doi: 10.1111/j.1365-2672.2004.02426.x 15546412 · doi ↗ · pubmed ↗
- 8Coon K. L. Vogel K. J. Brown M. R. Strand M. R. (2014). Mosquitoes rely on their gut microbiota for development. Mol. Ecol. 23, 2727–2739. doi: 10.1111/mec.1277124766707 PMC 4083365 · doi ↗ · pubmed ↗