State‐of‐the‐Art Strategies for Circular RNA in Cancers: Opportunity and Challenge
Zehao Ding, Zai Luo, Liao Zhang, Shaopeng Zhang, Renchao Zhang, Zhengjun Qiu, Chen Huang

TL;DR
Circular RNAs are emerging as key regulators in cancer, offering new diagnostic and therapeutic opportunities despite challenges in their study and application.
Contribution
This review provides a comprehensive overview of circRNA functions, biogenesis, and therapeutic potential in cancer, emphasizing recent advances and future directions.
Findings
CircRNAs act beyond microRNA sponges, functioning as protein scaffolds and peptide templates in cancer.
They play roles in tumorigenesis, immune evasion, and tumor–microbiome interactions, making them promising biomarkers.
Emerging AI tools and engineered therapies like CAR-T are being explored to harness circRNA potential in precision oncology.
Abstract
Circular RNAs (circRNAs) are characterized by their covalently closed structure, remarkable stability, and precise spatiotemporal regulation, evolving from once‐overlooked transcriptional byproducts to pivotal molecular regulators. In addition to their well‐established function as microRNA sponges, circRNAs serve as protein scaffolds, transcriptional modulators, and even templates for functional peptide synthesis. This review synthesizes recent breakthroughs across the entire circRNA life cycle, encompassing biogenesis, degradation, nucleocytoplasmic transport, and extracellular vesicle‐mediated secretion, while systematically analyzing their multifaceted involvement in tumorigenesis, immune evasion, metastatic dissemination, programmed cell death, and tumor–microbiome crosstalk. We highlight their exceptional potential as liquid biopsy biomarkers and critically assess translational…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
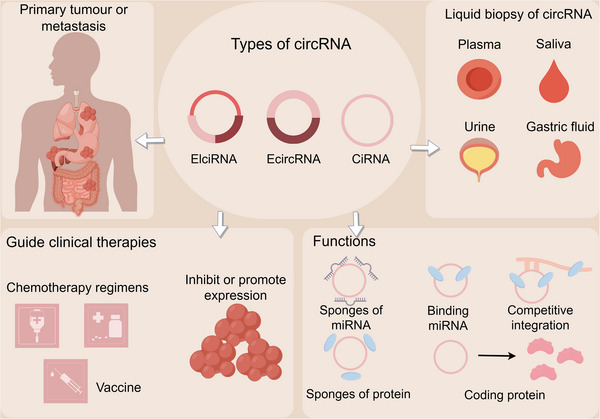 FIGURE 1
FIGURE 1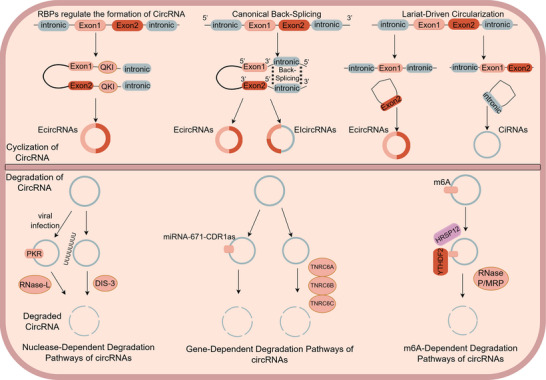 FIGURE 2
FIGURE 2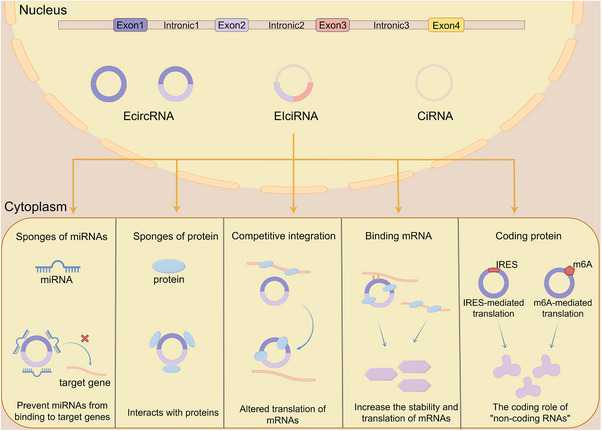 FIGURE 3
FIGURE 3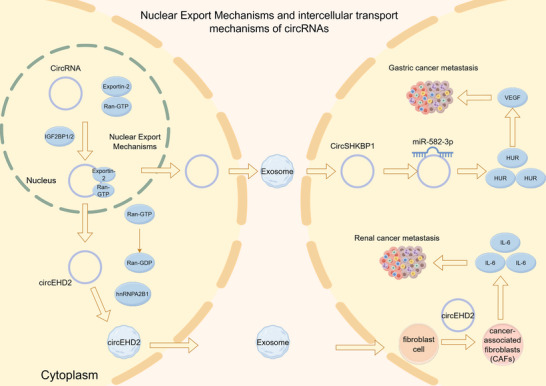 FIGURE 4
FIGURE 4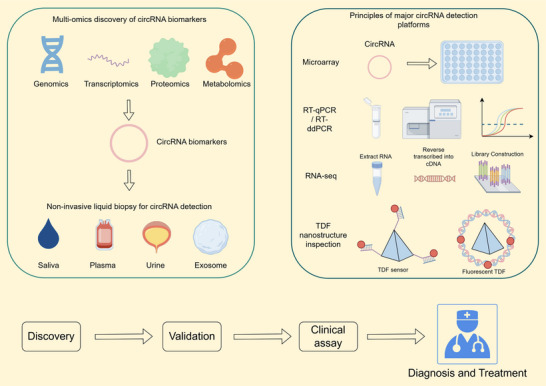 FIGURE 5
FIGURE 5| CircRNA | Cancer type | Sample type | Expression | Functional mechanism | Clinical relevance | References |
|---|---|---|---|---|---|---|
| circSHKBP1 | Gastric cancer | Tissue/exosome | Upregulated | Promotes angiogenesis and metastasis via miR‐582‐3p/HUR/VEGF | Diagnostic/prognostic | [ |
| circEHD2 | Renal cancer | Serum exosome | Upregulated | Activates CAF transition via hnRNPA2B1‐mediated IL‐6 secretion | Prognostic | [ |
| circUHRF1 | Hepatocellular carcinoma | Exosome | Upregulated | Sponges miR‐449c‐5p → TIM‐3 ↑ → NK cell exhaustion | Immune evasion/prognostic | [ |
| circTUBGCP4 | Colorectal cancer | Exosome | Upregulated | Promotes endothelial migration and tube formation | Prognostic | [ |
| circRNF216 | Colorectal cancer | Tissue | Downregulated | Sponges miR‐576‐5p → ↑ZC3H12C → ↑CD8+ T‐cell infiltration | Diagnostic/tumor suppression | [ |
| circp53 (hsa_circp53_0041947) | Multiple myeloma | Tissue | Downregulated | Encodes p53‐209aa → activates mitochondrial apoptosis | Diagnostic/therapeutic | [ |
| circSATB1 | Colorectal cancer | Tissue | Upregulated | RNF25/FKBP8/mTOR/EMT axis → metastasis | Prognostic | [ |
| circFAM13B | Nasopharyngeal carcinoma | Tissue | Downregulated | Inhibits lymphangiogenesis/↑5‐year survival | Prognostic | [ |
| circTFRC | Gastric cancer | Tissue | Upregulated | Recruits ELAVL1 → stabilizes SCD1 mRNA → ferroptosis resistance | Therapeutic target | [ |
| circMAT2B | Hepatocellular carcinoma | Tissue | Upregulated | miR‐338‐3p/PKM2 axis → glycolysis ↑ | Metabolic reprogramming/prognostic | [ |
| circACC1 | Colorectal cancer | Tissue | Upregulated | Forms complex with AMPK β/γ subunits → fatty acid metabolism ↑ | Prognostic | [ |
| circ_0000003 | Tongue SCC | Tissue | Upregulated | miR‐330‐3p/GLS axis → ↑α‐KG, ↑ATP → proliferation ↑ | Prognostic | [ |
| circ_0000808 | NSCLC | Tissue | Upregulated | miR‐1827/SLC1A5 axis → ↑glutamine metabolism | Prognostic | [ |
| circCDYL | Breast cancer | Tissue | Upregulated | Sponges miR‐1275 → ↑ATG7/ULK1 → promotes autophagy | Prognostic | [ |
| circHIPK2 | Colorectal cancer | Tissue | Upregulated | FUS–EIF4A3 complex → ↑TAZ translation | Early diagnostic biomarker | [ |
| Cancer type | Experimental model | Target circRNA | Specific pathway | References |
|---|---|---|---|---|
| Gastric cancer | Cell line, CDX | circMTHFD2L | circMTHFD2L/CM‐248aa/SET/PP2A/AKT;ERK;P65 | [ |
| Gastric cancer | Cell line, CDX | circHIPK3 |
circHIPK3/miR‐508‐3p/Bcl‐2/beclin1/SLC7A11 | [ |
| Nasopharyngeal carcinoma | Cell line, CDX | circFIP1L1 | circFIP1L1/miR‐1253/EIF4A3/PTEN | [ |
| Lung adenocarcinoma | Cell line, CDX | cEMSY | cEMSY/TDP‐43/mtDNA/cGAS/STING | [ |
|
Breast cancer (triple negative) | Cell line, PDO | circPVT1 | circPVT1/miR‐33a‐5p/c‐MYC/GLS1 | [ |
| Ovarian cancer | Cell line, CDX, PDO | hsa_circ_0010467 | AUF1/hsa_circ_0010467/miR‐637/LIF/STAT3 | [ |
| Breast cancer (HER2‐low) | Cell line, PDX | crVDAC3 | crVDAC3/HSPB1 | [ |
| Hepatocellular carcinoma | Cell line, CDX, PDX | circMDK | circMDK/miR‐346;miR‐874‐3p/ATG16L1/PI3K–AKT–mTOR | [ |
| Breast cancer (HER2‐negative) | Phase I clinical trial | CircFAM53B | circFAM53B/IRES‐driven translation/FAM53B‐219aa peptide/HLA‐I&II presentation/antigen‐specific T cell activation/tumor killing | [ |
| Large‐cell lymphoma (ALK‐positive anaplastic) | Clinical trial | / | / | [ |
- —National Natural Science Foundation of China10.13039/501100001809
- —Young Scientists Fund of the National Natural Science Foundation of China
- —2021 Shanghai ‘Rising Stars of Medical Talent’ Youth Development Program: Outstanding Youth Medical Talents
- —Key Projects of the Interdisciplinary Program of Medicine and Engineering at Shanghai jiao Tong University
- —Shanghai Rising‐Star Program Sailing Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircular RNAs in diseases · Cancer Mechanisms and Therapy · MicroRNA in disease regulation
Introduction
1
CircRNAs constitute a unique class of noncoding RNAs (ncRNAs), distinguished by their circular structure formed through covalently closed loops of multiple nucleotides [1]. The first observation of circRNA was made in an RNA virus in 1976 [2]. Initially, circRNA was regarded as a result of aberrant splicing, a perspective largely shaped by technological limitations and other factors. However, with the ongoing advancements in sequencing technology and bioinformatics, it has become evident that circRNA constitutes an abundant and evolutionarily conserved class of RNA, demonstrating widespread and intricate tissue‐, cell type‐, or stage‐specific expression patterns [3, 4]. Unlike linear RNAs, circRNAs undergo noncanonical splicing to form a covalently closed loop structure without free 3′ or 5′ ends. This unique structure makes them less susceptible to degradation by exonucleases and thus more stable than linear RNAs [5]. In recent years, numerous studies have confirmed that circRNAs possess critical biological functions and play a vital role in the onset and progression of diseases. For instance, specific subsets of circRNAs have been shown to promote tumor proliferation, migration, and metabolic reprogramming [6, 7, 8, 9]. Moreover, translation‐competent circRNAs encode functional proteins that regulate oncogenic signaling pathways [10, 11]. Recent advances have further elucidated their roles in mediating therapeutic resistance and highlighted their potential as engineered circRNA vaccines for emerging immunotherapeutic strategies [12, 13, 14, 15, 16]. Emerging evidence from various oncological studies indicates that certain circRNAs show significant potential as novel tumor biomarkers, potentially outperforming traditional markers like CEA [17, 18, 19].
Global cancer epidemiology continues to be a critical research priority. According to the latest World Health Organization assessment, there were 19.96 million new cancer cases and 9.74 million cancer‐related deaths worldwide in 2022 [20]. In 2013, two groundbreaking studies published consecutively in Nature clarified the molecular mechanisms by which circRNAs function as competitive endogenous RNAs (ceRNAs). These studies provided the first definitive evidence that circRNAs regulate miRNA activity through their sponging effects [21, 22]. These landmark discoveries reshaped our understanding of circRNAs, elevating them from perceived “splicing artifacts” to a diverse category of endogenous regulatory RNA molecules. As circRNA research has advanced, their multifaceted biological roles have been increasingly elucidated. In 2017, two studies published in Molecular Cell provided the first definitive evidence that endogenous circRNAs have protein‐coding potential in vivo [23, 24]. Specifically, circ‐ZNF609, originating from myoblasts, was demonstrated to undergo cap‐independent translation, yielding a functional protein [23]. This groundbreaking discovery not only challenges the conventional understanding of circRNAs but also opens new avenues in ncRNA and proteomics research. In 2022, Wei and colleagues achieved a significant breakthrough by developing a circRNA‐based vaccine effective against SARS‐CoV‐2 and its variants of concern. This innovation not only provided a promising new vaccine candidate for COVID‐19 but also established a groundbreaking technological platform. The success of this approach has since facilitated rapid advancements in circRNA‐based therapeutics and vaccine development, with applications extending to oncology research [25].
This review offers a comprehensive overview of the evolving field of circRNAs, covering their biogenesis, classification, degradation, and regulatory factors. We delve into their functional mechanisms, emphasizing their roles in tumorigenesis and their emerging protein‐coding potential. Additionally, we discuss the clinical potential of circRNAs, particularly as biomarkers for early cancer detection and as novel therapeutic targets (Figure 1). Last, we highlight the transformative impact of artificial intelligence on circRNA research and address the challenges in translating these findings into clinical practice.
Overview of circular RNA in tumors. Various types of circRNAs can be detected through liquid biopsies from plasma, saliva, urine, and gastric fluid, providing insights into the status of both primary and metastatic tumors. By functioning as sponges for miRNAs and proteins, engaging in competitive interactions, and encoding peptides, circRNAs have the potential to guide clinical therapies, including chemotherapy regimens, vaccines, and strategies to modulate circRNA expression.
Biosynthesis of CircRNA
2
The biogenesis, degradation, and intercellular communication of circRNAs exhibit remarkable mechanistic diversity in humans. Different subtypes of circRNAs undergo different modes of formation and turnover [26]. A comprehensive understanding of these processes is essential for elucidating the context‐dependent functions of circRNAs across various diseases. Therefore, we summarize the current knowledge on the major pathways governing circRNA circularization, degradation, as well as their nuclear export and extracellular transport (Figure 2).
The mechanisms and processes of cyclization and degradation of circRNA. The upper panel illustrates the circularization of precursor mRNAs into circRNAs through three mechanisms: RBP‐mediated exon circularization, canonical back‐splicing of flanking introns, and lariat‐driven circularization. The lower panel summarizes the major degradation pathways of circRNAs, including: (1) nuclease‐dependent pathways activated by viral infection; (2) gene‐dependent degradation, such as miR‐671‐directed CDR1as decay mediated by TNRC6 family proteins; and (3) N6‐methyladenosine (m6A)‐dependent degradation.
Circularization Mechanisms of CircRNA
2.1
Canonical Back‐Splicing
2.1.1
Similar to linear RNAs, circRNAs are also derived from precursor RNAs and transcribed by RNA polymerase II. However, their splicing process is distinct: the pre‐mRNA forms a loop through covalent bonding between the 5′ splice donor site downstream of an exon and the 3′ splice acceptor site upstream [27]. This loop then undergoes intron removal by cleavage to produce exonic circRNAs (EcircRNAs). During this reverse splicing process, some introns may remain uncleaved, resulting in the formation of exon–intron circRNAs (EIcircRNAs) [28].
Lariat‐Driven Circularization
2.1.2
Unlike the canonical back‐splicing mechanism, lariat‐driven circularization begins with exon‐skipping events during pre‐mRNA splicing. In this process, flanking introns are ligated into circular structures, while the skipped exons form m^7^G‐capped lariat intermediates. Subsequent back‐splicing then produces mature EcircRNAs. Multiple studies have shown that SF3B1 impairs exon recognition, thereby promoting intronic EIcircRNA biogenesis through exon‐skipping‐dependent circularization [29, 30]. The lariat‐driven formation of intronic circRNAs (CiRNAs) fundamentally differs from that of EcircRNAs. During canonical splicing, intronic lariats are typically degraded. However, if they contain a 7‐nucleotide GU‐rich motif at the 5′ splice site and an 11‐nucleotide C‐rich sequence near the 3′ end, these lariats evade debranching in standard linear splicing and instead fold into circular conformations via back‐splicing (Figure 2) [31, 32].
RNA‐Binding Proteins‐Dependent Cyclization Mechanism
2.1.3
Certain RNA‐binding proteins (RBPs) enhance circRNA biogenesis by specifically recognizing motifs within the flanking introns of pre‐mRNAs. Through either dimerization induction or complementary sequence bridging, these RBPs promote the spatial approximation of upstream and downstream back‐splice sites [33]. This structural rearrangement establishes an optimal configuration for the spliceosome to catalyze back‐splicing and generate circRNAs (Figure 2) [34]. The initial dimerization event allows RBPs to bridge adjacent exons, as illustrated by the Quaking protein, which forms homodimers through its C‐terminal (C‐term) domain, effectively clamping down on target motifs in a spatially constrained manner [35]. Furthermore, the FUS (fused in sarcoma) protein binds to the intronic regions flanking back‐splicing junctions (BSJs), dynamically modulating circRNA biogenesis. It exhibits context‐dependent dual functionality, acting as both a splicing activator and repressor [36]. Additionally, certain RBPs compete for binding to splice sites or regulatory elements, thereby suppressing the formation of linear mRNA and indirectly enhancing back‐splicing. For example, Kadener et al. reported that the muscleblind (MBL) protein binds to sequences surrounding the third intron of its own pre‐mRNA, promoting the production of the circular transcript circMbl while simultaneously inhibiting the canonical splicing of the corresponding linear isoform [37].
Classification of CircRNA
2.2
Most of the literature categorizes circRNAs into three main types: EcircRNAs, EIcircRNAs, and CiRNAs [38]. Recent studies have demonstrated that circRNAs encompass not only antisense circRNAs but also intergenic circRNAs [39]. The majority of identified circRNAs are exon circRNAs and intronic circRNAs. Although the splicing of circRNAs primarily occurs in the nucleus, EcircRNAs are predominantly exported to the cytoplasm, whereas CiRNAs remain sequestered in the nucleus (Figure 3) [27, 39].
Classification and function of circular RNA. In the nucleus, precursor mRNAs undergo back‐splicing to generate EcircRNAs, exon–EIcircRNAs, and ciRNAs. Upon export to the cytoplasm, circRNAs engage in various regulatory processes, including: (1) functioning as molecular sponges for miRNAs and RNA‐binding proteins; (2) competitively modulating mRNA translation; (3) enhancing the stability and translational efficiency of target transcripts; and (4) serving as templates for protein or micropeptide synthesis through IRES‐ or m6A‐mediated cap‐independent translation.
Mechanisms of CircRNA Degradation
2.3
Nuclease‐Dependent Degradation Pathways of CircRNAs
2.3.1
Previous studies have established that RNase L, an interferon‐induced endoribonuclease, mediates comprehensive degradation of circRNAs during viral infection [40]. Clinical observations have revealed marked decreases in circRNA levels in patients with systemic lupus erythematosus and chronic inflammatory skin disorders [40]. Under normal physiological conditions, circRNAs form intramolecular double‐stranded structures (16–26 bp) that interact with and inhibit dsRNA‐activated protein kinase (PKR), thereby preserving immune equilibrium. During viral infection, viral dsRNA stimulates 2′–5′‐oligoadenylate synthetases to generate 2–5A, which activates RNase L's endonucleolytic function and initiates circRNA breakdown [41]. Simultaneously, PKR‐mediated phosphorylation of eIF2α suppresses overall protein synthesis to inhibit viral proliferation. Importantly, this degradation pathway, which is specifically induced during viral infection, does not fully explain circRNA turnover under physiological circumstances [41]. A recent study by Chen et al. uncovered a novel degradation pathway mediated by DIS3 (chromosome transmission fidelity protein 3), a bifunctional endo/exoribonuclease that maintains circRNA homeostasis in noninfected cells [42].
DIS3 encodes a highly conserved ribonuclease with dual catalytic activities: 3′–5′ exoribonuclease activity mediated by its RNB domain and endoribonuclease activity through the PIN (PilT N‐terminus) domain [43]. Genetic studies demonstrate that DIS3 depletion leads to a significant upregulation of over 60% of circRNA, while showing minimal impact on their linear counterparts [44]. This DIS3‐mediated circRNA degradation is evolutionarily conserved, occurs in the cytoplasm, and strictly depends strictly on DIS3's endonucleolytic activity, yet remains independent of the RNA exosome complex. Additionally, sequence enrichment analysis indicates that DIS3 preferentially targets circRNAs containing U‐rich motifs, suggesting sequence‐specific recognition [45].
Gene‐Dependent Degradation Pathways of CircRNAs
2.3.2
In contrast to enzyme‐mediated degradation pathways, multiple genes have been identified as participating in circRNA decay through distinct mechanisms [46, 47]. Ago2, a ubiquitously expressed member of the Argonaute protein family, plays a key role in this process. CDR1as, a functionally important circRNA containing over 60 miR‐7 binding sites with critical implications in various pathologies, demonstrates near‐perfect complementarity to miRNA‐671 [48]. This structural feature facilitates experimental confirmation of Ago2‐mediated cleavage and degradation of CDR1as upon recognition of the miRNA‐671–CDR1as complex (Figure 2). Additionally, GW182 has been shown to participate in circRNA degradation. RNAi library screening in Drosophila DL1 and S2 cells revealed significant enrichment of specific circRNAs following GW182 depletion. Systematic genetic screens have uncovered protein‐coding genes that functionally regulate circRNA degradation pathways. Genome‐wide RNAi screening in Drosophila DL1 and S2 cells identified GW182 as a pivotal regulator of circRNA turnover. GW182 depletion markedly increased the abundance of circular isoforms originating from the dati and laccase2 transcripts. Structural analysis demonstrated that GW182 possesses six functionally distinct domains: an Ago‐binding domain, ubiquitin‐associated domain (UBA), glutamine‐rich domain (Q‐rich), middle region (Mid), RNA‐recognition motif, and C‐term region. Systematic deletion mutagenesis revealed that specifically disrupting the Mid domain eliminated the circRNA destabilization effect, highlighting its essential role in circRNA degradation. In humans, GW182 orthologs consist of three genes: TNRC6A, TNRC6B, and TNRC6C. Knockdown of each ortholog consistently increased circRNA accumulation, demonstrating their conserved function in circRNA decay pathways [48].
N6‐Methyladenosine‐Dependent Degradation Pathways of CircRNAs
2.3.3
N^6^‐methyladenosine (m^6^A) is a prevalent internal modification found in eukaryotic mRNAs and ncRNAs. Its canonical function in circRNAs involves regulating RNA stability through the recruitment of specific reader proteins and degradation complexes [49]. YTHDF2 is an evolutionarily conserved RBP characterized by a canonical YTH (YT521‐B homology) domain that specifically recognizes and binds to m^6^A‐modified RNA [50]. YTHDF2 recognizes m^6^A modifications and recruits the adaptor protein HRSP12, along with the RNase P/MRP endonuclease complex, to specific sites on circRNAs, where HRSP12 binds upstream of the YTHDF2 binding site and RNase P/MRP cleaves at downstream positions (Figure 2). This coordinated recruitment results in precise circRNA cleavage and subsequent rapid degradation [51, 52].
Nuclear Export Mechanisms of CircRNAs
2.4
CircRNAs are primarily generated within the nucleus through backsplicing. However, their functional roles, such as serving as miRNA sponges, interacting with proteins, or undergoing translation, require precise subcellular localization, mainly in the cytoplasm, although a subset remains in the nuclear. Therefore, the nuclear export processes of circRNAs are tightly regulated.
Ran‐GTP, a small GTPase, forms an intracellular concentration gradient that provides directional cues for various nucleocytoplasmic transport processes [53]. As a nuclear export receptor, Exportin‐2 (XPO2) mediates the translocation of RNA molecules from the nucleus to the cytoplasm. Notably, XPO2 serves as the sole export receptor for circRNA nuclear export, though it does not directly bind circRNA. This process relies critically on the adaptor proteins IGF2BP1 and/or IGF2BP2, which are mRNA localization‐associated factors. These adaptors demonstrate increased circRNA binding affinity in the presence of Ran‐GTP. Thus, circRNAs are exported via a Ran‐GTP‐dependent pathway, where XPO2 functions as the export receptor while IGF2BP1 (or IGF2BP2) physically links the circRNA cargo to the XPO2/Ran‐GTP complex. Upon cytoplasmic hydrolysis of Ran‐GTP to Ran‐GDP, the circRNA is subsequently released (Figure 4) [54].
Intercellular communication and function of circular RNA. CircRNAs are exported from the nucleus to the cytoplasm via pathways involving exportins and Ran GTPase. They are subsequently packaged into exosomes and transported to neighboring cells. This exosome‐mediated intercellular communication enables circRNAs to reshape the tumor microenvironment and promote cancer metastasis.
The Intercellular Transport Mechanisms of CircRNA
2.5
Emerging evidence suggests that circRNAs not only function intracellularly but also participate in intercellular communication through extracellular secretion and uptake by neighboring or distant cells, playing pivotal roles in developmental, immune, and oncogenic processes [55, 56]. This intercellular transport is primarily mediated by extracellular vesicles (EVs).
Exosomes, a major EV subtype ranging from 40 to 160 nm in diameter, are secreted by various cell types and serve as vehicles for transferring lipids, proteins, DNA, and ncRNAs [57]. Following uptake by recipient cells, exosome‐derived circRNAs can exert regulatory functions, most notably through their miRNA sponge activity. By competitively binding to miRNAs, these circRNAs prevent miRNA interaction with target gene 3′‐untranslated regions (UTRs), effectively modulating downstream signaling pathways implicated in cancer progression and metastasis (Figure 4) [58, 59, 60]. For instance, Xie et al. revealed that exosomal circSHKBP1 promotes gastric cancer metastasis by sponging miR‐582‐3p, which subsequently elevates HUR expression. This upregulation stimulates VEGF secretion and angiogenesis in gastric cancer tissues [61]. Similarly, Liu et al. discovered that circEHD2, which is highly expressed in renal cancer tissues and metastatic patient serum exosomes, is selectively packaged into exosomes through hnRNPA2B1 binding. Following exosomal transfer to fibroblasts, circEHD2 induces their transformation into cancer‐associated fibroblasts (CAFs). These activated CAFs then secrete IL‐6, creating a prometastatic microenvironment that drives renal cancer progression [62] (Figure 4).
Characteristics of CircRNAs
3
The key characteristics of circRNAs encompass their remarkable structural stability, distinct tissue‐ and disease‐specific expression, and significant spatiotemporal heterogeneity across various cell types, tumor regions, and microenvironmental components. These attributes underpin the functional roles of circRNAs, influencing their expression patterns and regulatory functions in cancer and other diseases [63]. Our review integrates these three fundamental characteristics and elucidates the underlying principles that support their potential as biomarkers and their broader clinical applications.
High Stability
3.1
CircRNAs are generated through the backsplicing of pre‐mRNA, resulting in covalently closed circular structures that lack the 5′ cap and 3′ poly(A) tail found in linear RNAs. This unique topology enables circRNAs to evade deadenylation, decapping, and exonucleolytic degradation. Quantitative studies in mammary gland cells have revealed a median circRNA half‐life of 18.8 to 23.7 h, in stark contrast to the 4.0–7.4‐h half‐life observed for their homologous linear RNAs [64]. Furthermore, comparative half‐life analysis has shown that m^6^A‐modified circNSUN2 exhibits a stability of over 24 h in colorectal cancer cells, sharply contrasting with the approximately 4‐h half‐life of its linear transcript counterpart [65]. Consequently, circRNAs are considered to be stably preserved in bodily fluids such as plasma, exosomes, and urine. The noninvasive detection of circRNA expression changes in these fluids allows for effective monitoring of dynamic circRNA alterations in cancer patients. Through genome‐wide screening, Zhang et al. identified a biomarker panel consisting of five circRNAs, which can be used for the early noninvasive detection of pancreatic ductal adenocarcinoma (PDAC). Notably, when combined with CA19‐9, the diagnostic performance is significantly enhanced [66].
Tissue Specificity
3.2
Extensive studies have confirmed that circRNAs are ubiquitously expressed across eukaryotic tissues, with notable enrichment in mammalian brain and skeletal muscle [21, 67, 68]. Tumor‐specific circRNA expression exhibits significant heterogeneity: for instance, circFAM53B is specifically enriched in breast cancer tissues but undetectable in normal mammary glands, a pattern also observed in melanoma [69]. CircE7 is the only identified HPV‐encoded circRNA, exclusively expressed in HPV‐positive head and neck squamous cell carcinoma (HNSCC) [70]. Additionally, circZKSCAN1 is highly expressed in normal liver, where it maintains cellular homeostasis, but is significantly downregulated in hepatocellular carcinoma (HCC). Interestingly, this circRNA is significantly upregulated in tumor tissues such as colorectal and lung ADCs [71]. A study on cervical cancer identified 215 circRNAs that were upregulated in SCC but downregulated in ADC, with 50 showing inverse regulation patterns [72]. Moreover, numerous circRNAs exhibit dysregulated expression patterns across various cancer types. For example, CircHIPK3 (hsa_circ_0000284), a circular RNA (circRNA) derived from the exons of the HIPK3 gene, is widely expressed in diverse tissues and cell types. It is significantly upregulated in several malignancies, including gastric, colorectal, and breast cancers, but notably downregulated in bladder cancer [73]. Overall, circRNAs display distinct tissue‐specific expression patterns in vivo. Comprehensive sequencing analyses of various tumor tissues enable researchers to elucidate the complex regulatory functions of specific circRNAs in tumor initiation and progression, as well as their dynamic roles across diverse biological contexts [74].
Spatiotemporal Heterogeneity
3.3
The exceptional stability and tissue specificity of circRNAs provide a solid foundation for functional studies, while their expression patterns exhibit spatiotemporal dynamics across various biological contexts.
Multiple tumor studies have confirmed the spatiotemporal heterogeneity of circRNAs. For instance, in gliomas, the most prevalent primary malignant brain tumors with a poor prognosis, circRNA sequencing revealed a NEIL3‐derived circRNA that is upregulated in tumor tissues and shows expression levels positively correlated with pathological grades (I–IV) [75]. A groundbreaking study published in Nature Communications utilized spatial analysis technologies to investigate the distribution of CDR1as in colon cancer tissues. Surprisingly, the researchers discovered that ciRS‐7 was not localized within cancer cells but was highly expressed in the stromal cells of the tumor microenvironment (TME). This finding challenges the conventional view that CDR1as is an endogenous molecule of cancer cells and underscores the spatial heterogeneity of circRNAs across different tumor regions [76]. Moreover, circRNA profiles exhibit significant functional heterogeneity between tumor cells and the TME. In tumor cells, circRNAs primarily regulate malignant behaviors such as proliferation, invasion, metastasis, metabolic reprogramming, epithelial–mesenchymal transition (EMT), and drug resistance. By contrast, circRNAs within TME cells mainly participate in exosome‐mediated intercellular communication, affecting processes like immunosuppression, fibrosis, and angiogenesis [77]. Shi et al. reported that exosomal circUHRF1 in liver cancer suppresses natural killer (NK) cell function and induces NK cell exhaustion by inhibiting miR‐449c‐5p, thereby upregulating TIM‐3 expression [78]. Similarly, Kang et al. demonstrated that exosomes derived from CRC cells promote vascular endothelial cell migration and tube formation by inducing filopodia formation and endothelial tip cell orientation. Silencing circTUBGCP4 in CRC cell‐derived exosomes (CRC‐CDEs) effectively inhibited endothelial cell migration, tube formation, tip cell formation, and CRC metastasis [79].
Functional Mechanisms of CircRNAs
4
The diverse regulatory mechanisms of circRNAs form a crucial foundation for their roles in cancer research and clinical applications, allowing them to exert their biological functions through multiple pathways [26]. Our review summarizes their primary functional mechanisms, which include serving as ceRNAs to modulate gene expression, interacting with RBPs to influence transcriptional programs and protein activity, and generating biologically active peptides through noncanonical translation (Figure 3).
Competing Endogenous RNA
4.1
The role of circRNAs has garnered significant attention. miRNAs are a class of endogenous small RNAs, typically 20–24 nucleotides in length. Multiple miRNAs can coregulate the expression of individual genes, thereby fine‐tuning gene expression and participating in various physiological activities [80]. Numerous studies have demonstrated that circRNAs function as molecular sponges by competitively binding to miRNAs [81, 82, 83]. This interaction blocks miRNA‐target mRNA binding, thereby relieving the suppression of target genes. Consequently, circRNAs precisely modulate downstream signaling pathways. This intricate circRNA–miRNA–mRNA regulatory axis exerts profound biological effects on tumorigenesis and progression across diverse cancer types [84]. For instance, circ‐CDYL accelerates early‐stage HCC progression by sponging miR‐892a and miR‐328‐3p, leading to the upregulation of hepatoma‐derived growth factor (HDGF) and hypoxia‐inducible factor asparaginyl hydroxylase (HIF1AN). This mechanism activates the HDGF‐mediated PI3K–AKT–mTORC1/β‐catenin pathway while suppressing HIF1AN‐regulated NOTCH2 signaling [85]. Additionally, Mou et al. identified significantly downregulated circRNF216 expression in colorectal cancer tissues and cells. Functioning as a ceRNA, circRNF216 sequesters miR‐576‐5p, thereby alleviating its suppression of the target gene ZC3H12C and reducing N‐cadherin levels [86]. Furthermore, circRNF216 enhances CD8+ T cell infiltration by upregulating ZC3H12C, ultimately inhibiting colorectal cancer progression. Downregulation of circCCDC9 was observed in gastric cancer cells, and functional analysis demonstrated its direct binding to miR‐6792‐3p, which alleviates the suppression of the tumor suppressor gene CAV1 [87].
CircRNA–RBP Interaction Patterns
4.2
RBPs constitute a class of proteins that specifically interact with RNA molecules. By recognizing distinct RNA sequences or structures, RBPs regulate the entire RNA life cycle, including posttranscriptional processes, and their functional dysregulation is closely associated with various disease pathologies [88, 89].
First, circRNAs can specifically bind RBPs, thereby influencing their interactions with other proteins. For example, circ0006646, a circRNA associated with HCC and poor prognosis, binds nucleolin (NCL) and disrupts its interaction with the E3 ubiquitin ligase TRIM21. This suppression of K48‐linked ubiquitination and degradation stabilizes the oncogenic protein NCL. Accumulated NCL inhibits p53 translation in vivo, ultimately promoting cancer cell metastasis [90, 91]. In laryngeal SCC, circMTCL1 directly binds the RBP C1QBP, inhibiting its ubiquitination‐mediated degradation. This complex coactivates Wnt/β‐catenin signaling, driving disease progression in vivo [92]. Second, circRNAs can interact with specific proteins to dysregulate their native functions or elicit novel biological effects. CircHuR suppresses the proliferation, invasion, and metastasis of gastric cancer by inhibiting HuR transcription through blocking the binding of the transcription factor CNBP to the HuR gene promoter [93]. Circ‐hnRNPU, an exon‐derived circRNA from heterogeneous nuclear ribonucleoprotein U (hnRNPU), physically interacts with the non‐POU domain‐containing octamer‐binding protein (NONO). This interaction induces cytoplasmic retention of NONO, thereby suppressing tumor progression through downregulation of glycosyltransferases. The underlying mechanism involves inhibiting both nuclear NONO‐mediated c‐Myc transcriptional activation and cytoplasmic NONO‐promoted mRNA stabilization [94]. Third, circRNAs can bind specific cis‐regulatory elements to modulate transcription factors or epigenetic modifications, thereby altering gene expression. For instance, circRHOT1 recruits and anchors TIP60 (a histone acetyltransferase) to the NR2F6 promoter region, activating its transcription and consequently promoting HCC proliferation and metastasis [95]. Additionally, circMRPS35 specifically recruits KAT7 to the FOXO1/FOXO3a promoters, inducing H4K5 acetylation to activate transcription. This process orchestrates a tumor‐suppressive transcriptional network involving p21, p27, and E‐cadherin, thereby inhibiting gastric cancer proliferation and metastasis [96].
CircRNA Encode Proteins
4.3
In addition to their previously recognized functions, recent studies have revealed that some circRNAs can undergo translation. Advances in bioinformatics platforms and next‐generation sequencing technologies have enabled the identification of numerous circRNAs containing open reading frames (ORFs) capable of encoding proteins [97]. Due to the absence of 5′ caps and 3′ termini, circRNA translation is facilitated through cap‐independent mechanisms. Internal ribosome entry sites (IRES), located within the 5′ untranslated regions of mRNAs, enable translation initiation in the absence of canonical initiation factors, either partially or completely [96]. The IRES‐dependent translation mechanism is widespread among circRNAs. CircRNAs can initiate translation by directly recruiting ribosomes through the insertion of a synthetic IRES upstream of the start codon. Fan et al. discovered that numerous short IRES‐like elements are significantly enriched in endogenous circRNAs, serving as trans‐acting factors that promote cap‐independent translation [98]. Another mechanism of circRNA translation is mediated by m^6^A, the most prevalent internal RNA modification in eukaryotes. Yang et al. found that a single m^6^A site effectively recruits the 43S complex to initiate translation, aided by the YTH domain family protein 3 (YTHDF3) and eIF4G2. The translation is inhibited by the m^6^A demethylase FTO but enhanced by the adenosine methyltransferases METTL3/14 [99]. Pan et al. identified that circ‐YAP is significantly upregulated in colorectal cancer with liver metastasis and is associated with poor prognosis. This circRNA encodes a novel 220‐amino acid truncated protein, termed YAP‐220aa, which competitively binds LATS1, leading to YAP dephosphorylation and nuclear translocation. Consequently, this process activates prometastatic genes [100]. Additionally, circCAPG encodes a 171‐amino acid protein that binds to the kinase STK38, disrupting its interaction with the E3 ubiquitin ligase SMURF1. This disruption suppresses MEKK2 ubiquitination and degradation, leading to sustained activation of the MEK/ERK signaling pathway, thereby promoting proliferation and metastasis in triple‐negative breast cancer (TNBC) cells [101].
Biological Function of CircRNAs in Tumors
5
Emerging evidence indicates that circRNAs play pivotal regulatory roles in cancer by modulating key biological processes, including tumor cell proliferation, immune evasion, invasion, and metastasis, and various forms of cell death. Additionally, circRNAs are intricately involved in metabolic reprogramming, cellular senescence, and epigenetic regulation. They also influence the remodeling of the TME through interactions with the microbiota [77]. By summarizing these mechanisms, we can elucidate how circRNAs coordinate tumor initiation, progression, and metastasis across multiple biological levels (Table 1).
The Role of CircRNAs in Tumor Growth and Proliferation
5.1
Cellular genesis and demise are governed by tightly regulated, genetically programmed pathways [112]. In normally developing tissues, these processes remain under homeostatic control, whereas some cells undergo clonal expansion and acquire additional genetic and phenotypic alterations that enhance their survival and proliferation [113]. These transformations are primarily driven by multifactorial interplay among epigenetic and metabolic reprogramming, microenvironmental cues, and other determinants. The resultant alterations in gene expression or mutations confer competitive advantages over normal cells, leading to the clonal expansion of tumor subpopulations with acquired malignant traits. Ultimately, iterative cycles of mutation, selection, and proliferation culminate in tumor progression and oncogenic sequelae [114, 115].
Certain circRNAs modulate tumor proliferation primarily through dysregulating metabolic pathways. For instance, Mo et al. identified a novel circRNA, circRNF13, which is pathologically downregulated in NPC tissues and cell lines. Mechanistically, circRNF13 upregulates SUMO2 protein expression, suppresses glycolytic flux, and consequently inhibits the AMPK–mTOR signaling axis, ultimately attenuating tumor proliferation [116]. In HCC studies, Mei et al. demonstrated that circEPB41(2) interacts with the m^6^A demethylase FTO to modulate mRNA stability of histone deacetylase SIRT6, thereby facilitating transcriptional activation of lipid metabolism‐associated genes. Experimental validation revealed that circEPB41(2) knockdown significantly suppresses HCC cell proliferation and attenuates tumor growth in vivo [117]. Furthermore, circRNAs modulate tumor growth by regulating cell cycle progression and apoptosis. Yu et al. identified that the TP53 gene produces circp53 (hsa_circp53_0041947) with coding function. Clinical analyses revealed significant downregulation of circp53 in multiple myeloma patients, correlating with reduced survival. Mechanistically, the translation product circp53‐209aa activates mitochondrial apoptosis through a unique pathway. The researchers also engineered an EV‐based delivery platform (E7/Her2–Lamp2b–EV) for targeted circp53 delivery, which potently suppressed xenograft tumor growth and enhanced bortezomib therapeutic efficacy [102].
The Role of CircRNAs in Tumor Immune Escape
5.2
The immune system plays a crucial role in identifying and removing aberrant cells, preserving cellular homeostasis, and preventing malignant transformation [118]. Tumor immune evasion constitutes a pathological condition where tumors bypass immune surveillance mechanisms to sustain uncontrolled growth. This process involves multiple strategies such as altered antigen presentation, creation of immunosuppressive microenvironments, and suppression of immune cell function, collectively enabling cancer cell survival [119, 120, 121]. Emerging evidence highlights specific circRNAs as key modulators of these immune evasion pathways.
Functionally, hsa_circ_0007991 acts as a competitive ceRNA that binds to and sequesters miR‐505‐3p, leading to increased expression of the endoplasmic reticulum chaperone CANX. This molecular interaction not only promotes tumor proliferation but also diminishes CD8^+^ T cell cytotoxicity. Additional investigations confirmed the cytoplasmic stability of this circRNA, which enhances immune evasion through dual activation of the TGF‐β receptor/SMAD2 axis and PD‐L1 signaling pathways [122]. Similarly, the highly expressed circRNA hsa_circ_0136666 in gastric cancer facilitates immune evasion by modulating the hsa_circ_0136666/miR‐375/PRKDC axis. This regulatory network promotes PRKDC‐mediated phosphorylation of PD‐L1, preventing its proteasomal degradation and resulting in abnormal PD‐L1 accumulation. Miu et al. showed that LNP‐encapsulated siRNA targeting hsa_circ_0136666 effectively reduces recruitment of immunosuppressive MDSCs and Treg cells, substantially improving the efficacy of anti‐PD‐L1 therapy [123]. Moreover, hsa_circ_0020397 binds to and neutralizes miR‐138, thereby upregulating both PD‐L1 and telomerase reverse transcriptase (TERT). This mechanism enhances tumor survival while weakening T cell‐mediated cytotoxicity. Additionally, circPIAS1 inhibits STAT1 phosphorylation, suppressing ferroptosis and consequently diminishing the effectiveness of anti‐PD‐1 treatment [124]. Targeting these circRNAs presents a promising strategy for overcoming tumor immune evasion, potentially opening new avenues for clinical cancer therapy.
The Role of CircRNAs in Tumor Invasion and Metastasis
5.3
Although primary tumors pose a significant threat, their danger to patients is relatively limited compared with tumor invasion and metastasis, which are major causes of clinical mortality [125, 126]. Specific circRNAs play crucial roles in metastatic progression by modulating EMT, promoting angiogenesis and lymphangiogenesis, and establishing premetastatic niches.
Liver metastasis is the most common distant metastatic pattern in CRC. Research has shown that circSATB1 promotes CRC cell metastasis both in vitro and in vivo. Mechanistically, circSATB1 orchestrates RNF25‐mediated ubiquitin‐dependent degradation of FKBP8, thereby derepressing FKBP8's inhibition of the mTOR pathway, activating EMT, and enhancing tumor invasion and hepatic dissemination [103]. Additionally, circAKT3 and circAQR facilitate tumor metastasis in prostate and thyroid carcinomas, respectively, through EMT modulation in vivo [127, 128]. Beyond EMT, tumor angiogenesis is a critical prerequisite for invasion and metastasis. Pathological angiogenesis not only supplies oxygen and nutrients to tumor cells but also aids in the clearance of metabolic waste. Jiang et al. identified significant upregulation of circFNDC3B in oral SCC, which positively correlates with lymph node metastasis. Functional analyses in vitro and in vivo demonstrate that circFNDC3B promotes vascular endothelial tubulogenesis and induces lymphatic endothelial cell migration and tube formation, thereby enhancing tumor metastasis [129]. Conversely, circFAM13B suppresses lymphangiogenesis and lymph node metastasis in NPC. Tissue microarray analysis of 255 patients revealed a 35% improvement in 5‐year survival rates among those with high circFAM13B expression, and multivariate analysis confirmed its status as an independent prognostic factor [104]. Leveraging multiomics insights into circRNA regulatory networks will facilitate the development of novel therapeutics and diagnostic biomarkers targeting tumor invasion and metastasis.
The Role of CircRNAs in Tumor Cell Death
5.4
Cellular death is a fundamental mechanism for maintaining tissue homeostasis, and its dysregulation is intricately linked to cancer pathogenesis [104, 130]. Targeting tumor cell death represents a promising therapeutic strategy, and growing evidence indicates that circRNAs play pivotal regulatory roles in cell death pathways. Hsa_circ_0003141 is significantly upregulated in HCC tissues, where it functions as a molecular sponge for miR‐1827, derepressing UBAP2 inhibition and thus suppressing the mitochondrial apoptotic pathway [131]. CircTFRC drives ferroptosis resistance in gastric cancer by recruiting the RBP ELAVL1 to stabilize SCD1 mRNA through 3′UTR binding. This enhances monounsaturated fatty acid synthesis, suppresses lipid peroxidation, and ultimately antagonizes ferroptosis. In xenograft models, circTFRC siRNA treatment reduced tumor volume by 58% and pulmonary metastatic nodules by 72%, highlighting its therapeutic potential for gastric cancer intervention [105].
The Role of CircRNAs in the Tumor Microbiome
5.5
Tumors harbor a complex microbial ecosystem comprising bacteria, fungi, and viruses, collectively termed the intratumoral microbiota. This microbial community plays a crucial role in tumor progression by modulating immune regulation, inflammatory responses, and metabolic reprogramming [132]. To date, such microbial communities have been detected in at least 33 different human cancer types [133]. Notably, broad‐spectrum antibiotic treatment induces gut dysbiosis, which in turn disrupts the TME. Employing deep sequencing combined with animal models and fecal microbiota transplantation. Zhang et al. revealed that mmu_circ_0000730 suppresses mmu‐miR‐466i‐3p to upregulate the oncogenic transcription factor SOX9 in lung cancer stem cells. These findings highlight the gut microbiota's potential to regulate cancer progression and metastasis through the IL‐11‐mediated circRNA/miRNA/SOX9 signaling axis [134]. Although emerging evidence demonstrates that gut microbiota influences neuropsychiatric disorders via specific circRNAs (e.g. circ_0001239 and circHIPK2), the role of circRNAs at the tumor‐microbiota interface remains poorly understood [135]. Further mechanistic studies are urgently needed to elucidate the functional relationships between circRNAs, microbial communities, and tumor pathogenesis.
CircRNA and Metabolic Reprogramming
5.6
Metabolic reprogramming is a hallmark of tumor cells, characterized by alterations in glucose, amino acid, and lipid metabolism to support rapid proliferation, stress adaptation, and microenvironmental remodeling [76]. Accumulating evidence suggests that circRNAs serve as key regulators of these metabolic pathways by functioning as miRNA sponges, interacting with proteins, or encoding short peptides [136, 137, 138].
Enhanced glycolysis is a prominent feature of metabolic reprogramming in cancer cells, enabling them to meet increased energy and biosynthetic demands. Bioinformatic analyses by Wang et al. revealed that circMAT2B is significantly upregulated in HCC tissues and cell lines. Patients with high circMAT2B expression exhibited reduced overall survival. Mechanistic studies demonstrated that circMAT2B acts as a molecular sponge for miR‐338‐3p, thereby upregulating pyruvate kinase M2 (PKM2), a key glycolytic enzyme directly targeted by miR‐338‐3p. This circMAT2B/miR‐338‐3p/PKM2 axis enhances glycolysis and promotes HCC progression [106]. In lipid metabolism, Wu et al. discovered that circACC1 stabilizes and enhances the enzymatic activity of the AMPK holoenzyme by forming a ternary complex with its regulatory β‐ and γ‐subunits. This mechanism simultaneously promotes fatty acid synthesis and oxidation, providing energy and biosynthetic precursors to sustain the rapid proliferation of CRC cells. In tumor xenograft models, circACC1 knockdown suppressed tumor growth, while its overexpression enhanced tumor progression [107]. Glutamine metabolism is one of the most extensively studied aspects of amino acid reprogramming in cancer, with multiple circRNAs identified as pivotal regulators. Yao et al. demonstrated that circ_0000003 acts as a molecular sponge for miR‐330‐3p, thereby relieving its inhibition of glutaminase (GLS). This upregulates GLS expression, enhances glutamine consumption, increases α‐ketoglutarate (α‐KG) production, and elevates ATP generation, collectively driving proliferation, migration, and invasion in tongue SCC [108]. Similarly, Wang et al. reported that circ_0000808 facilitates non‐small cell lung cancer (NSCLC) progression by sequestering miR‐1827, which releases the suppression of solute carrier family 1 member 5 (SLC1A5), a key glutamine transporter. Consequently, glutamine uptake, glutamate, and α‐KG production are increased, ultimately promoting NSCLC proliferation, migration, and invasion [109]. Although numerous circRNAs play indispensable roles in metabolic reprogramming, the complexity of metabolic networks in tumor cells necessitates further fundamental research to elucidate their context‐specific mechanisms. Such investigations are essential for establishing the foundation for clinical translation and for developing novel therapeutic strategies targeting tumor metabolism.
CircRNA Regulates Cellular Senescence
5.7
Cellular senescence represents a fundamental biological process that persists throughout an organism's lifespan, where maintaining the delicate equilibrium between senescent cell clearance and new cell proliferation is critical for physiological homeostasis. Emerging evidence highlights this process as a pivotal pathogenic driver in diverse diseases, ranging from neurodegenerative disorders and cardiovascular conditions to carcinogenesis, with circRNAs emerging as key players within these regulatory networks [139, 140, 141].
As established in prior research, specific circRNAs exert their biological functions through protein interactions. Notably, circDNA2v demonstrates significant upregulation in colorectal cancer, where it binds to IGF2BP3 protein and protects it from ubiquitin‐mediated degradation, thereby prolonging its half‐life. The stabilized IGF2BP3 subsequently binds to and stabilizes proto‐oncogene c‐Myc mRNA, amplifying its expression. Importantly, circDNA2v inhibition results in c‐Myc downregulation and triggers tumor cell senescence [140]. Additionally, circFoxo3 interacts with crucial cell cycle regulators such as cyclin‐dependent kinase 2 (CDK2) and the CDK inhibitor p21, forming a ternary circFoxo3–p21–CDK2 complex that impedes cell cycle progression from G1 to S phase, ultimately promoting cellular senescence [142]. Certain circRNAs contribute to senescence through molecular sponge mechanisms. Liang et al. revealed that circCDYL functions as a molecular sponge for miR‐1275, consequently upregulating ATG7 and ULK1 expression to enhance autophagy and accelerate breast cancer progression [110]. However, the dual role of senescent tumor cells, exhibiting both protumorigenic and antitumorigenic effects, is mediated by complex interactions between various senescence‐associated secretory phenotype factors and the immune microenvironment. While targeting circRNAs to induce tumor cell senescence presents a promising antitumor strategy, further functional characterization of relevant circRNAs remains essential to validate their clinical potential [143].
CircRNA Regulates Epigenetic Modifications
5.8
Epigenetic modifications are heritable alterations in gene function that occur without changes to the underlying DNA sequence. These include DNA and RNA methylation, histone modifications, and posttranscriptional regulation mediated by ncRNAs. Such mechanisms allow cells sharing identical genetic backgrounds to acquire distinct functional identities. Increasing evidence indicates that circRNAs are integral to these processes through complex interactions with DNA, proteins, and other RNAs, thereby exerting profound effects on the initiation and progression of tumors [144].
Regarding DNA methylation regulation, FECR1, a circRNA derived from the FLI1 gene, has been shown to bind CpG islands in the promoters of both FLI1 and DNMT1. By recruiting the TET1 demethylase, FECR1 promotes site‐specific DNA demethylation and transcriptional activation. Simultaneously, FECR1 suppresses DNMT1 expression, leading to reduced global DNA methylation levels and enhanced invasiveness of breast cancer cells [145]. Additionally, certain circRNAs can modulate chromatin architecture and influence gene transcription by regulating histone modifications. Chromatin immunoprecipitation sequencing data indicate that circZKSCAN1 alters chromatin states through the repression or activation of specific histone marks, thereby contributing to the progression of melanoma [146]. m^6^A is the most prevalent internal modification in eukaryotic mRNAs, and certain circRNAs also undergo m^6^A modification. This epitranscriptomic mark significantly influences circRNA stability and function. For example, in NSCLC, m^6^A‐modified circIGF2BP3 (hsa_circ_0079587) downregulates the immune checkpoint protein PD‐L1, thereby impairing antitumor immune responses. Conversely, m^6^A‐modified circNSUN2 promotes colorectal cancer liver metastasis by facilitating the assembly of a circNSUN2/IGF2BP2/HMGA2 ternary complex [147]. Super‐enhancers (SEs), genomic regions with exceptionally strong enhancer activity, have been implicated in the regulation of circRNA biogenesis. SEs are marked by dense occupancy of transcription factors, cofactors, and histone modifications such as H3K27ac. A recent study demonstrated that SEs enhance both the diversity and abundance of circRNA isoforms from host genes by promoting RBP recruitment and modulating transcriptional elongation. Importantly, the same study identified a pan‐cancer tumor‐suppressor signature termed CircRNA Isoform Reduction from Shortened Enhancers in Cancer, which exhibits strong prognostic value in lung ADC [148].
Collectively, these findings highlight the central role of circRNAs in epigenetic and transcriptional regulation. By bridging DNA, RNA, and protein regulatory layers, circRNAs contribute to tumor initiation, progression, and metastasis, underscoring their potential as diagnostic biomarkers and therapeutic targets.
CircRNAs Serve as Promising Biomarkers for Early Tumor Diagnosis
6
Temporal Dynamics of CircRNA Expression in Early Tumor Progression
6.1
The development of cancer is a complex and prolonged process, typically progressing from normal tissue through inflammatory responses and precancerous lesions to early‐stage cancer, often spanning decades [149]. Consequently, investigating the molecular mechanisms underlying this multistep progression is critically important.
CRC currently has the highest incidence among gastrointestinal malignancies and is strongly associated with ulcerative colitis (UC) [150]. In clinical practice, early detection primarily relies on endoscopic examination; however, due to limitations in patient compliance and variability in endoscopist expertise, a significant proportion of early‐stage tumors remain undiagnosed. Recent research has revealed that circRNAs exhibit stage‐specific expression dynamics with distinct characteristics during the progression from inflammatory bowel disease to colorectal tumorigenesis. In UC patients, the downregulation of multiple circRNAs, including circCCND1, circ_0001021, and circHECTD1, is observed in colonic epithelial tissues. Mechanistic studies show that these circRNAs contribute to intestinal epithelial barrier integrity [151, 152, 153]. Additionally, circHIPK2 is moderately elevated in intestinal epithelial cells during active UC and further increases in the colonic adenoma stage. Mechanistic studies demonstrate that FUS mediates the formation of a circHIPK2–EIF4A3 complex, which promotes TAZ translation and ultimately drives epithelial cell proliferation in colitis and colorectal carcinogenesis [111]. Similarly, cervical cancer development progresses through a well‐defined multistep continuum, evolving from normal cervical epithelium to HPV infection/inflammation, cervical intraepithelial neoplasia (CIN), and ultimately early‐stage invasive carcinoma [154]. Notably, recent research has shown a stepwise upregulation of circ‐LDHA expression across normal cervical tissues, CIN lesions, and cervical carcinomas. This circRNA maintains persistent elevation under HPV infection and Acinetobacter lwoffii costimulation, thereby orchestrating malignant transformation via the circ‐LDHA/miR‐34a/HMGB1 signaling axis [155].
Noninvasive Detection of CircRNA
6.2
Advances in multiomics technologies, including genomics, proteomics, and metabolomics, have facilitated the discovery and clinical translation of numerous promising biomarkers for oncology [156]. Research indicates that circRNA expression is closely associated with tissue homeostasis, cellular dynamics, and tumorigenesis, positioning circRNAs as viable biomarkers for various pathologies, including cancer [26, 157]. In clinical practice, noninvasive biomarkers and liquid biopsy techniques are widely used for real‐time monitoring of disease progression and therapeutic response [158]. CircRNAs exhibit stable expression and relatively high abundance in biofluids such as saliva, plasma, serum, and exosomes, making them ideal candidates for noninvasive liquid biopsy‐based cancer detection [159, 160]. Several primary noninvasive methods for circRNA detection include microarray analysis, RNA‐sequencing (RNA‐seq), reverse transcription quantitative polymerase chain reaction (RT‐qPCR), and reverse transcription droplet digital PCR (RT‐ddPCR) (Figure 5).
Flow chart of clinical translation research on circRNA. Multiomics approaches are employed to identify candidate circRNA biomarkers that can be detected in noninvasive liquid biopsy specimens, including saliva, plasma, urine, and exosomes. These biomarkers are subsequently quantified using various detection platforms, such as microarrays, RT‐qPCR/RT‐ddPCR, RNA‐seq, and nanostructure‐based sensors. Following validation and the development of clinical assays, these biomarkers may ultimately aid in disease diagnosis and treatment.
Microarray‐based circRNA detection relies on specific hybridization between predesigned oligonucleotide probes and target circRNAs. The process begins with the selective elimination of linear RNAs via RNase R digestion, followed by hybridization of fluorescence‐labeled circRNAs to immobilized BSJ‐targeting probes on the microarray chip [161]. Scanning the fluorescence signal intensity enables semi‐quantitative analysis of circRNA expression profiles [162]. While microarrays are cost‐effective and efficient, their ability to detect new transcripts is limited due to predesigned templates, and their accuracy is compromised at extremely low or high gene expression levels. Consequently, microarrays are often used as a preliminary screening step before conducting quantitative circRNA analyses.
RNA‐seq has become a prevalent tool in molecular biology research. Unlike microarrays, RNA‐seq can discover novel circRNAs and facilitate large‐scale testing in clinical samples. The core principle involves identifying unique back‐splicing patterns derived from the circular conformation of circRNAs. This process starts with digesting total RNA with RNase R to selectively degrade linear RNAs, followed by purification and construction of strand‐specific whole‐transcriptome libraries. Subsequent fragmentation of RNA templates enables the synthesis of double‐stranded cDNA fragments for sequencing. Advanced algorithms, such as CIRI and CIRI‐deep, have been developed for circRNA detection. CIRI uses a BWA–MEM‐based circular alignment methodology to identify BSJs by detecting soft‐clipping signals in paired‐end mapping data. Its latest version incorporates a convolutional neural network (CNN) module to filter false positives through sequence‐context features, such as splice site conservation and RNA secondary structure [163]. However, due to the absence of poly(A) tails in circRNAs, conventional transcriptomic library preparation methods such as polyA enrichment fail to efficiently capture them, resulting in the loss of circRNA expression information in most traditional single‐cell and spatial transcriptomic datasets. To address this limitation, researchers have developed CIRI‐deep, a deep learning model that circumvents dependence on traditional back‐splicing signals and instead utilizes specific cis‐regulatory elements and trans‐acting factors as input features. Evaluation results demonstrate that CIRI‐deep enables reliable prediction of differentially spliced circRNAs in transcriptomic sequencing data, thereby achieving accurate cell‐type‐specific circRNA resolution at single‐cell and spatial levels [164].
The NanoString nCounter platform enables direct digital counting of RNA molecules without amplification, providing highly specific and sensitive quantification. By employing a pair of probes (a capture probe and a reporter probe) designed to span the BSJ, the technology ensures precise discrimination between circRNAs and their linear counterparts. During solution hybridization, these probes bind specifically to the target circRNA, forming a stable hybrid complex. After purification and immobilization, the complexes are immobilized onto a cartridge for subsequent digital imaging and quantification [165]. This amplification‐free approach avoids PCR‐induced biases, delivering more direct and reliable measurements with high specificity and reproducibility. Furthermore, the platform supports multiplex detection, enabling simultaneous quantification of hundreds of circRNAs in a single run. Its compatibility with diverse clinical specimens, including plasma, urine, and formalin‐fixed paraffin‐embedded tissues, underscores its translational potential [166]. Despite these advantages, the NanoString nCounter system is relatively costly, especially for large sample cohorts, and relies on custom‐designed probes specific to the BSJ, which require extended design and synthesis timelines. These factors hinder its widespread adoption, particularly in resource‐limited laboratories [167].
RT‐qPCR is extensively used for the detection and validation of circRNAs. It employs different primers flanking the BSJ locus to amplify circRNA fragments and detect circRNAs. A standard procedure involves treating total RNA extracted from the sample with RNase R, followed by RT to generate the complementary DNA of the circRNA, and subsequently determining and analyzing the fluorescent signal [168]. This technology offers superior sensitivity, detection speed, and time efficiency for circRNA detection, making it well suited for clinical laboratories. However, RT‐qPCR has significant errors in accurately quantifying trace nucleotides in plasma and may overestimate circRNA concentrations due to strand displacement and roll‐over replication during RT [169, 170]. To address these challenges, an assay utilizing RT‐ddPCR was developed. RT‐ddPCR is an emerging assay technology with higher sensitivity and accuracy than conventional PCR, which has been demonstrated to be the most suitable method for quantifying circRNA in tissues and plasma [168]. RT‐ddPCR is unaffected by standard curves and amplification kinetics, allowing for absolute quantification and avoiding the impact of differences in PCR amplification efficiency. It requires a small sample size and exhibits high detection sensitivity, making it appropriate for clinical samples [171]. However, these conventional methods necessitate skilled personnel, specialized laboratory facilities, and processing times exceeding 24 h, making rapid and sensitive circRNA detection challenging. To address this, Yao et al. developed a strategy using tetrahedral DNA frameworks for selective recognition and high‐affinity capture of intact circRNAs. This approach involves constructing multivalent DNA nanostructure probes spatially complementary to circRNA conformations. By conjugating two‐photon fluorophores to the probes, it enables real‐time imaging of freely migrating circRNAs in living systems while simultaneously facilitating dynamic tracking during circRNA delivery processes [172].
Exploiting the unique circular structure of circRNAs, researchers have developed a method that selectively amplifies circular templates without amplifying linear RNAs, known as RT‐rolling circle amplification (RT‐RCA). In this approach, circRNAs are first reverse‐transcribed into cDNA using specifically designed primers, followed by circularization of the cDNA template via ligases or similar enzymes. DNA polymerase then continuously extends along the circular template, generating long tandem repeat sequences. The amplified products are subsequently detected [173]. Zhu et al. combined RT‐RCA with a carefully designed networked hybridization chain reaction (HCR) to develop a circRNA detection method based on isothermal networked HCR. In this approach, the networked structure forms a stable mesh with RT‐RCA products containing multiple tandem repeats under isothermal conditions, significantly enhancing the fluorescent signal. Experimental results demonstrated that the optimized networked HCR system exhibits higher detection efficiency and excellent selectivity for DNA strands containing multiple repeat sequences. Notably, this method allows accurate detection of specific circRNAs in authentic biological samples without RNase R enrichment, providing a simple and effective platform for the detection of low‐abundance circRNAs [174]. However, this detection method has so far been limited to circRNAs in specific tissues, and the impact of abundant interfering substances in complex biological samples, such as blood or saliva, remains unclear. Future studies should aim to expand the application of RT‐RCA to various biological specimens, including body fluids and feces, while enhancing its resistance to interference. Additionally, improving the efficiency of simultaneous detection of multiple circRNAs will be essential to meet the demands of high‐throughput and complex sample analyses [173, 175].
CircRNA and Encoded Proteins as Therapeutic Targets
7
There is increasing evidence that circRNAs and the proteins they encode function as ceRNAs by sequestering miRNAs and upregulating downstream genes. This mechanism ultimately contributes to various aspects of tumor progression, metastasis, and enhances therapeutic resistance by inhibiting cell death processes such as apoptosis and ferroptosis. Consequently, these specific circRNAs and their associated pathways are anticipated to serve as novel therapeutic targets, offering additional opportunities for clinical intervention (Table 2).
Therapeutic Targeting Across Models
7.1
Numerous studies utilizing in vitro cell cultures and cell line‐derived xenograft (CDX) models have established that targeting circRNAs or their encoded proteins can effectively suppress tumor progression and metastasis, while also reversing tumor cell resistance to chemotherapy, radiotherapy, and immunotherapy. For instance, CM‐248aa, a protein encoded by circMTHFD2L, has been shown to inhibit gastric cancer cell proliferation and metastasis when overexpressed in gastric cancer cell lines, CDX models, and lung metastasis models [176]. Research on cisplatin resistance mechanisms revealed that silencing circHIPK3 in cisplatin‐resistant gastric cancer cells and CDX models suppresses tumor progression and metastasis by activating the miR‐508‐3p/Bcl‐2/beclin1/SLC7A11 axis to promote ferroptosis, thereby reducing cisplatin resistance [177]. Similarly, elevated circFIP1L1 expression stabilizes PTEN mRNA via the miR‐1253/EIF4A3 axis, inducing apoptosis in NPC cells and enhancing radiosensitivity, as demonstrated in CDX models [178]. Notably, the circRNA cEMSY has been identified as an immunogenicity enhancer both in vitro and in vivo (in immunosuppressed tumor models), where it triggers immunogenic cell death in lung ADC cells, thereby amplifying antitumor immune responses and potentiating the efficacy of immune checkpoint inhibitors [179].
Preclinical investigations employing patient‐derived organoid (PDO) and patient‐derived xenograft (PDX) models have further underscored the clinical translational potential of targeting circRNA‐related pathways in cancer therapy, although no related drugs have advanced to clinical trials. For example, circPVT1 depletion sensitizes breast cancer cells to glutaminase inhibitor‐induced cytotoxicity through the miR‐33a‐5p/Myc/GLS1 axis, a finding validated in TNBC‐derived organoids [180]. In ovarian cancer cells, CDX models, and PDOs, silencing hsa_circ_0010467 markedly enhances cisplatin's antitumor effects by modulating the AUF1/hsa_circ_0010467/miR‐637/LIF/STAT3 axis [181]. Additionally, in HER2‐low PDX models, paritaprevir binds VDAC3‐derived circRNA, promoting HSPB1 protein ubiquitination and degradation while elevating ferroptosis levels, thereby overcoming trastuzumab deruxtecan resistance in HER2‐low breast cancers [182].
Through a comprehensive approach involving four HCC animal models (subcutaneous, metastatic, PDX, and orthotopic), Du et al. demonstrated that the poly(β‐amino ester)‐based circMDK siRNA delivery system (PAE–siRNA complex) effectively silences circMDK, inhibiting HCC progression and metastasis in vivo [183].
CircRNA Reshaping CAR‐T Therapeutic Areas
7.2
CircRNAs are reshaping landscape and practicing CAR‐T therapy at multiple levels. Numerous studies have shown that, at the adaptive immunity level, circRNAs can impair T‐cell function by upregulating immune checkpoint molecules or diminish their antitumor activity by reducing T‐cell infiltration into tumor tissues [147, 186, 187, 188]. At the innate immunity level, circRNAs can induce macrophage polarization toward the M2 (tumor‐promoting) phenotype and suppress the tumor‐killing function of NK cells [78, 189]. Given that CAR technology functions by activating these immune cells, circRNAs may attenuate CAR‐T cell and associated immune cell activity through these mechanisms, thereby diminishing the antitumor efficacy of CAR technology [63].
Conversely, a growing body of research highlights the unique application potential of circRNAs in adoptive cell therapy. On one hand, circRNAs can directly participate in CAR‐T preparation processes. Their covalently closed‐loop structure confers high stability and precise gene regulatory functions, enhancing the persistence and specificity of CAR‐T cells in cancer treatment. Specifically, compared with traditional mRNA‐based CAR‐T preparation methods, using circRNAs as a delivery system for precise insertion of CAR transgenes offers higher gene integration efficiency, enhanced gene tagging accuracy, and reduced production costs [190, 191, 192]. Additionally, circRNAs can minimize off‐target effects by reducing nonspecific binding of CAR‐T cells to healthy tissues through acting as miRNA sponges, or by enhancing the targeting accuracy of CAR‐T cells via increasing the expression of tumor‐specific receptors on CAR‐T cells and upregulating the levels of target antigens (such as HER2, CD19) on tumor cell surfaces [190, 193, 194]. When delivered in a vaccine, circRNAs can enhance the therapeutic efficacy of CAR‐T cells by regulating CAR‐T cell proliferation, infiltration, and exhaustion processes. Furthermore, they can optimize CAR‐T therapy by providing supplemental antigens or functioning as adjuvant enhancers.
A recent study demonstrated that delivering circRNACAR encoding anti‐HER2 CAR alongside immunocellular‐targeting lipid nanoparticles (LNPs) promotes the in vivo formation of a panCAR system encompassing CAR‐T, CAR‐NK, and CAR‐macrophage cells, significantly inhibiting tumor growth. The combined use of circRNA vaccines encoding the corresponding HER2 antigen enhances circRNACAR‐mediated antitumor therapy by increasing CAR‐T cell proliferation, promoting CAR‐T cell infiltration, and attenuating CAR‐T cell exhaustion [195]. Numerous prior studies have confirmed that circRNAs can reduce T‐cell exhaustion by acting as miRNA sponges, interacting with RBPs, regulating activation‐induced cell death, and activating T‐cell survival‐related transcription factors [194, 196, 197]. They also create a favorable environment for CAR‐T cell infiltration and activation by downregulating extracellular matrix protein expression and modulating cytokine and chemokine levels within the TME [194]. Moreover, circRNA vaccines may provide additional antigenic signals by encoding tumor‐specific antigens, thereby enhancing endogenous T‐cell immunity to assist CAR‐T cells in eliminating antigen‐deficient tumor cells [69, 198, 199]. CircRNAs may also function as “adjuvant enhancers”; for example, circFOREIGN activates type 1 conventional dendritic cells (cDC1s), enhancing antigen presentation efficiency, which may boost CAR‐T cell activation and proliferation capacity [198, 200]. Precisely due to circRNA's stable expression patterns and multifaceted regulatory functions, it is highly suitable as a biomarker for predicting CAR‐T outcomes [201].
Delivery Systems
7.3
Currently, cancer treatments utilizing circRNAs can be categorized into three primary approaches. The first approach involves delivering siRNAs, antisense oligonucleotides, or CRISPR–Cas13a to silence oncogenic circRNAs. The second approach entails constructing and delivering overexpression vectors for tumor‐suppressing circRNAs. The third approach involves delivering circRNAs that encode functional proteins to participate in cancer therapy [159].
The key to these circRNA‐based cancer therapies lies in the precise and efficient delivery of the target molecules to the desired tissues or cells. Common delivery systems for transporting circRNAs include LNPs, exosomes, viruses, peptide vectors, and composite nanoparticles [202]. Most studies have utilized adenoviral and lentiviral vectors as viral delivery systems to target circRNAs; however, due to their limited gene‐carrying capacity, high costs, and potential biosafety concerns, an increasing number of studies are now focusing on LNPs or engineered exosomes for circRNA delivery [203].
CircRNA Vaccine
7.3.1
CircRNA vaccines represent a novel class of vaccines capable of inducing the expression of antigens that elicit adaptive immune responses. The circRNAs used in these vaccines primarily possess protein‐coding functions. Compared with traditional mRNA vaccines, circRNA vaccines typically produce higher and more sustained antigen levels for several reasons.
First, circRNAs are covalently closed loop structures that resist degradation by RNA exonucleases, making circRNA vaccines structurally stable and easier to store [199]. Second, the lack of free ends in circRNAs avoids recognition by innate immune receptors such as RIG‐I, thereby reducing immunogenicity and cytotoxicity [204, 205]. Third, the roll‐over translation mechanism of circRNAs prolongs the expression time, making them an ideal vector for efficient protein expression [204, 205].
LNP Delivery System
7.3.2
LNPs have emerged as a predominant delivery vehicle for circRNA‐targeting therapeutics. These nanoparticles typically comprise four key components: ionizable cationic lipids, phospholipids, cholesterol, and polyethylene glycolated lipids [206]. The ionizable cationic lipids facilitate RNA encapsulation within LNPs through electrostatic interactions with negatively charged RNA molecules [207]. In a notable study, Xu et al. employed a high‐throughput combinatorial approach to develop optimized LNPs for efficient circRNA delivery to lung tumors. Their findings demonstrated that a single intratumoral injection of IL‐12 circRNA‐loaded LNPs significantly enhanced immune cell infiltration within the TME and induced marked tumor regression in the LLC1 murine tumor model [208]. Furthermore, Zhou et al. pioneered an innovative therapeutic platform utilizing in vitro circRNA (ivcRNA) for osteoarthritis treatment. Their strategy involves local intra‐articular administration of LNP‐encapsulated ivcRNA encoding therapeutic proteins. This delivery system exhibits exceptional stability and translational efficiency, enabling sustained expression of therapeutic proteins such as MSI2 and SOX5—outperforming m^1^Ψ‐modified linear mRNA in both durability and protein yield. The approach shows significant efficacy in mitigating osteoarthritis progression, representing a promising advancement in circRNA‐based therapeutic strategies [209].
Exosome Delivery System
7.3.3
Exosomes are endogenous EVs actively secreted by cells. Tumor‐derived exosomal circRNAs have been demonstrated to regulate immune cell antitumor activity and modulate immune checkpoint protein expression, while also promoting angiogenesis, participating in cancer metabolism, and contributing to tumor proliferation, metastasis, and drug resistance. Notably, various cellular components within the TME—including immune cells, CAFs, adipocytes, and mesenchymal stem cells—can influence tumor progression through exosomal circRNA secretion. These properties make exosomal circRNAs promising diagnostic and prognostic biomarkers for diverse tumor types [159].
Beyond naturally occurring exosomal circRNAs, engineered exosomes have emerged as effective vehicles for delivering therapeutic agents that directly target circRNA functions. Compared with synthetic delivery systems such as LNPs and nanoparticles, exosomes exhibit superior biocompatibility, reduced immunogenicity, and enhanced chemical stability, rendering them highly suitable as delivery vectors [210, 211]. For instance, Wang et al. developed exosomes loaded with si‐ciRS‐122, which effectively suppressed colon cancer cell proliferation and reversed oxaliplatin resistance [212].
Synthesis of CircRNA in Therapy
7.4
The large‐scale synthesis of circRNA is crucial for advancing the widespread adoption of circRNA therapeutics.
In vitro circRNA synthesis primarily employs three methods: chemical synthesis, enzyme‐mediated synthesis using DNA or RNA ligases, and nuclease‐based approaches relying on self‐splicing introns [213]. Chemical synthesis is suitable for producing small circRNAs of high purity but involves toxic reagents, poses potential biosafety risks, and is characterized by low efficiency and high cost. Enzymatic synthesis using DNA or RNA ligases is more appropriate for preparing medium‐to‐long circRNAs, offering advantages such as high cyclisation efficiency, minimal introduction of extraneous sequences, and low immunogenicity. A recent study developed an in vitro circularization strategy based on cis‐acting ligase ribozyme (RzL). RzL autonomously catalyses RNA circularization during transcription without GTP involvement and without by‐product generation, significantly enhancing circRNA synthesis efficiency and product purity [214]. Nuclease‐based approaches using self‐splicing introns are suitable for synthesizing circRNAs of varying sizes (up to 5 kb). Their reaction conditions are straightforward, making them suitable for large‐scale production, and the synthesized circRNA products are highly safe. Wang et al. constructed a novel chimeric permuted intron‐exon system. By engineering type I self‐splicing introns, they significantly enhanced the circularization efficiency of in vitro circRNA synthesis [215]. Du et al. developed a novel circRNA synthesis method based on trans‐Rzl‐based circularization, enabling efficient preparation of long‐chain circRNAs exceeding 8000 nt. This approach not only eliminates the need for bacterial sequence introduction but also supports the simultaneous incorporation of multiple RNA modifications. This system employs virus‐derived IRES for rolling‐circle translation (RCT), enhancing protein expression efficiency by over 7000‐fold [216]. Researchers have also proposed CIRC (complete self‐splicing intron for RNA circularization), a novel intron‐mediated circRNA synthesis method. Utilizing complete type I and type II introns, it achieves efficient, rapid RNA circularization while producing scarless, low‐immunogenicity circRNAs [217]. Furthermore, Zhu et al. designed a novel circRNA‐promoting RNA (cpRNA) that targets the flanking reverse complementary matches (RCMs) within pre‐mRNAs to enhance exon circularization and thereby promote circRNA biogenesis. Additionally, cpRNA specifically upregulates tumor‐suppressive circRNAs such as circZKSCAN1 and circSMARCA5, leading to the inhibition of HCC cell proliferation and migration, and offering a potential therapeutic strategy for diseases associated with low circRNA expression. However, its effectiveness is limited to circRNAs containing RCMs, which restricts its broader applicability [218].
Beyond in vitro synthesis, circRNA can also be generated in vivo. For instance, the nuclease‐based Tornado system efficiently produces intracellular circRNA, and when combined with virus‐like particles (VLPs), it enables sustained protein expression [219]. Further studies report that the Tornado system (twister‐optimized RNA for durable overexpression) enables efficient circRNA synthesis within mammalian cells, exhibiting significantly higher efficiency than conventional backsplicing methods. Using VLPs delivery systems, it achieves more sustained target protein expression, offering novel strategies for circRNA in vivo applications [189]. Tong et al. developed two novel circRNA synthesis strategies: type II intron‐mediated in vitro circularization and in vivo circularization utilizing the intracellular RtcB ligase. They established an efficient purification method without requiring high‐performance liquid chromatography, significantly enhancing circRNA yield and purity while reducing immunogenicity [220].
CircRNA structures typically comprise IRES, coding regions, and regulatory hairpin loops. Optimizing these structures enhances protein expression, reduces immunogenicity, and improves stability, ultimately boosting the overall efficacy of circRNA therapeutics [213]. For example, Chen et al. achieved a several‐hundred‐fold increase in circRNA protein yield by systematically optimizing five key components of synthetic circRNAs: the vector topology, 5′ and 3′ UTRs, IRES, and synthetic aptamers recruiting translation initiation machinery. Embedding an eIF4G‐recruiting aptamer (Apt‐eIF4G) within a specific IRES domain was found to significantly enhance circRNA translation efficiency [221].
The purification process for synthetic circRNA is critical for its clinical applications. A team recently developed a multistep purification strategy combining enzymatic treatment (RNase R) with cellulose chromatography (e.g., WMC), which efficiently removes immunogenic impurities (such as dsRNA and 5'‐ATP) from synthetic circRNA. This significantly reduces innate immune responses and increases circRNA yield by over tenfold. CircRNA purified via this strategy demonstrated over 300% efficiency in induced pluripotent stem cell reprogramming and achieved sustained, potent antitumor expression in CAR‐T cells, establishing a high‐quality RNA platform for regenerative medicine and cancer immunotherapy [222].
Currently, several clinical trials are exploring the therapeutic potential of specific circRNAs in cancer treatment. A clinical trial (NCT06530082) is currently evaluating the safety and tolerability of the CircFAM53B‐219aa DC vaccine, both as a monotherapy and in combination with camrelizumab, for the treatment of HER2‐negative advanced breast cancer. Previously, the same research team demonstrated the tumor‐suppressive effects of CircFAM53B‐219aa in vitro and in animal models [184]. Furthermore, an ongoing clinical trial (NCT05934045) is investigating the role of circRNAs as circulating biomarkers for predicting treatment resistance in ALK‐positive anaplastic large‐cell lymphoma, as well as their potential utility as therapeutic targets [185].
Opportunities in the Clinical Translation of CircRNAs
8
Applications and Prospects of CircRNA‐Based Vaccines
8.1
Currently available vaccines fall into five main categories: inactivated vaccines, viral vector vaccines, DNA vaccines, mRNA vaccines, and emerging circRNA‐based vaccines, each representing distinct technological platforms for immunization strategies. Traditional inactivated vaccines, exemplified by inactivated hepatitis B vaccines, are produced using physical and chemical methods to neutralize pathogens. These vaccines offer high safety profiles, enhanced stability, and straightforward manufacturing processes, though they exhibit relatively weaker immunogenicity [223, 224]. Viral vector vaccines, which use replication‐deficient adenoviral vectors to deliver pathogen antigen genes, provide advantages such as convenient storage conditions and efficient delivery. However, pre‐existing immunity can compromise their clinical efficacy [225]. DNA vaccines benefit from intrinsic DNA stability and the elimination of ultra‐low temperature storage requirements, but the potential for genomic integration poses risks for germline transmission [226]. Due to these limitations, the first three vaccine platforms have constrained applicability, positioning RNA‐based therapeutics at the forefront of both fundamental research and clinical translation.
mRNA vaccines, delivered via LNPs encapsulating antigen‐encoding transcripts, undergo cytoplasmic translation to elicit potent immunostimulatory effects, including robust antigen‐specific humoral and cellular immune responses that confer broad and durable immunity [227]. Critically, their rapid development cycle, requiring only genetic sequence information, enables prompt deployment during pandemics such as COVID‐19. However, mRNA vaccines also carry potential risks, as demonstrated by a study showing that the in vitro incorporation of N1‐methylpseudouridine (m1Ψ) into mRNA can induce ribosomal frameshifting, leading to the production of frameshifted products during the translation of BNT162b2 vaccine mRNA in both murine models and human recipients [228].
The covalently closed circular architecture of circRNAs confers high stability with extended half‐lives compared with linear RNAs. During translation, circRNAs utilize IRES to initiate RCT, enabling sustained generation of repetitive antigen epitopes that enhance immunogenicity while reducing dsRNA production. Consequently, circRNA‐based vaccines represent a rapidly advancing research domain [199].
In cancer immunotherapy, Lin et al. developed the world's first intranasal circRNA vaccine by encapsulating circRNAs in LNPs to elicit localized mucosal immunity. This vaccine induced potent antitumor T cell responses in animal models, specifically eliminating pulmonary tumors in mice while mitigating systemic adverse effects typically associated with intravenously administered mRNA vaccines. It also augmented CAR‐T cell responses to enhance therapeutic efficacy against tumor cells expressing specific tumor‐associated antigens [198]. Furthermore, wang et al. pioneered a circRNA‐based neoantigen vaccine expressing the HCC‐specific neoantigen Ptpn2_I383T. This vaccine promotes DC maturation and enhances DC‐mediated T cell priming, thereby potentiating T cell cytotoxicity against tumor cells. In murine tumor models, the vaccine demonstrated robust therapeutic and prophylactic efficacy while conferring long‐term protective immunity. Synergistic strategies integrating circRNA vaccines with chemotherapeutic agents are also emerging. Zhang et al. developed a circRNA–DC vaccine encoding tumor antigens FAPα and survivin, which elicited potent CD8^+^ T cell responses and significantly suppressed tumor growth. Combining this DC vaccine with gemcitabine substantially prolonged median survival in pancreatic cancer murine models [229].
More recently, a research team from China developed a carrier‐free, scalable, and programmable circRNA platform embedded with aptamers (Apt‐circRNA). Unlike traditional delivery systems, Apt‐circRNA does not require LNPs or exogenous adjuvants; instead, it achieves targeted delivery through its built‐in aptamer structure. When administered subcutaneously as a tumor vaccine, Apt‐circRNA can efficiently target DCs and be transported to draining lymph nodes, thereby eliciting a potent and safe antitumor immune response. Preclinical and clinical studies have already demonstrated its remarkable targeted immunogenicity and excellent safety profile, establishing a solid foundation for the future development and clinical translation of aptamer‐based circRNA vaccines [230].
Technological Innovations: Single‐Cell CircRNA Sequencing Reveals Tumor Heterogeneity
8.2
Accurately deciphering biological systems and their dysregulation in disease requires multiparametric analysis at the single‐cell level. While single‐cell sequencing has become essential in oncology research due to advancements in multiomics profiling platforms, analyzing circRNAs at single‐cell resolution remains challenging. The key limitations include: (1) the inherently low abundance of circRNAs, which often leads to frequent dropout events in conventional single‐cell workflows; and (2) the high sequence homology between circRNAs and their linear parental transcripts, necessitating both specific enrichment strategies and sophisticated computational discrimination algorithms. Zhao et al. achieved high‐resolution profiling and comprehensive characterization of circRNAs at single‐cell resolution by systematically integrating 171 published single‐cell full‐length transcriptome datasets. Their analysis of single‐cell datasets from 20 breast cancer patients revealed significantly higher circRNA expression in both normal and malignant cells from low‐grade tumors with favorable prognosis compared with high‐grade TNBCs. CircRNA abundance progressively increased during the epithelial‐to‐intermediate EMT, but paradoxically decreased in late EMT stages. Notably, circRNA expression in cancer cells dropped below the baseline levels observed in the mesenchymal phase of normal cells, highlighting tumor‐specific circRNA heterogeneity during mesenchymal transformation [231].
Translational Priorities: Integrating Databases Across Cancer Types
8.3
The rapid advancement of high‐throughput sequencing technologies has facilitated the development of multiple comprehensive circRNA databases dedicated to cancer research, significantly advancing pan‐cancer circRNA investigations. By integrating circRNA expression profiles, molecular signatures, and clinical information across diverse cancer types, these databases provide invaluable resources for systematically exploring the functional roles of circRNAs in oncogenesis. Here, we summarize key publicly available circRNA databases and highlight their core features.
The CSCD database series represents one of the earliest resources dedicated to tumor‐specific circRNAs. Its upgraded version, CSCD2, compiles over 100,000 circRNAs identified from more than 1000 cancer and normal samples. Its core strength lies in constructing interactive regulatory networks that enable intuitive exploration of circRNA–miRNA/RBP interactions. Additionally, CSCD2 provides precise predictions of full‐length circRNA sequences and ORFs [232].
TCCIA is a pioneering database that bridges circRNAs with cancer immunotherapy, integrating over 4000 tumor samples from 25 independent immunotherapy cohorts. The platform employs standardized reprocessing of raw RNA‐seq data and clinical phenotypes, coupled with systematic circRNA detection, enabling users to interactively screen circRNAs significantly associated with immunotherapy response and investigate their correlations with T‐cell infiltration, immune exclusion, and other TME phenotypes [233].
MiOncoCirc is a pioneering database that systematically links circRNAs with clinical phenotypes across cancer types, integrating data from over 2000 clinical specimens, including primary tumors, metastatic lesions, and rare subtypes. Its distinctive advantage lies in directly capturing patient‐derived circRNA expression profiles within TMEs, thereby circumventing potential biases inherent in cell line models. The implementation of exon‐capture sequencing technology by the research team demonstrates enhanced circRNA detection capacity compared with conventional workflows, offering superior sensitivity and specificity to facilitate circRNA biomarker discovery [234].
Protein‐coding circRNAs play pivotal roles in disease pathogenesis, and circDB represents the first dedicated database cataloging these functional circRNAs. It integrates over 35,000 human exon‐derived circRNAs with comprehensive annotations, including exon composition, genomic origins, molecular size, sequence length, and full‐length circular transcripts. To assess protein‐coding potential, the database systematically predicts IRES elements and ORFs exceeding 300 bp, providing the two top‐ranking IRES motifs for each entry based on computational scoring algorithms [235].
Beyond these established resources, novel circRNA databases continue to emerge, including circRNADisease, which encompasses nononcological disorders, and exoRBASE, dedicated to circRNAs in human blood exosomes. These databases enable researchers to explore circRNA–disease associations while also serving as cross‐validation platforms for assessing circRNA coding potential [236].
The Role of Artificial Intelligence in CircRNAs
8.4
Over the past decade, artificial intelligence has ushered in a new era in medicine, becoming an indispensable tool for clinical decision support and biomedical predictive analytics. By leveraging robust computational capabilities, AI efficiently processes and analyzes the high‐throughput sequencing data generated by modern biology [237]. Various deep learning architectures enable the prediction of novel drug targets and disease risks, while AI‐driven frameworks play crucial roles in circRNA screening, sequence annotation, and functional characterization [238, 239].
CircRNA Profiling: High‐Fidelity Identification and Association Mapping
8.4.1
Traditional circRNA screening methods typically require months to years from discovery to functional validation, often introducing potential biases due to technical variations [240, 241]. In contrast, AI‐driven approaches leverage deep learning frameworks to establish end‐to‐end predictive pipelines, significantly enhancing screening efficiency. This methodology begins with the curation of experimentally validated circRNA–disease associations from databases, followed by feature extraction and selection. The subsequent application of CNNs and autoencoders enables the prediction of novel circRNA–disease relationships, which are ultimately validated through targeted experiments or multiomics sequencing [242, 243].
Li et al. developed CDASOR, a deep learning framework that integrates convolutional and recurrent neural networks for circRNA–disease association prediction. CDASOR uses continuous k‐mer bag‐of‐words encoding to generate low‐dimensional circRNA sequence embeddings. These representations are processed by a 1D CNN for local feature extraction and then analyzed using bidirectional long short‐term memory (BiLSTM) networks to capture long‐range dependencies. Validation against published literature confirmed six out of ten top‐ranked predictions in case studies, demonstrating CDASOR's ability to generate biologically relevant associations [244]. However, existing deep learning models face limitations such as insufficient feature representation and high dependency on training data. To address these issues, a research team from China developed AGDFCDA, a computational framework that implements dual‐feature extraction strategies for circRNA–disease association prediction. This approach uses fully connected neural networks to reduce feature redundancy and initially extract latent representations of circRNAs and diseases. It then employs adaptive graph convolutional networks to learn comprehensive circRNA–disease embeddings for advanced feature extraction, and finally performs association prediction through multilayer perceptron architectures [245].
Sequence Prediction: Optimizing Cyclization Efficiency and Translation Capability
8.4.2
Optimization of conventional linear mRNAs typically focuses on the 5′/3′ UTRs and ORFs to achieve sustained and efficient translation. In contrast, circRNAs contain IRES motifs, enabling cap‐independent translation initiation beyond ORF elements. Properly folded functional IRES motifs facilitate the recruitment of the preinitiation complex, which is essential for efficient circRNA translation [216]. Additionally, optimizing the secondary structures and codon usage in circRNA sequence design significantly enhances transcript stability while concurrently improving translational efficiency and antibody responses [246]. To address the challenges of circRNA sequence optimization, Zhang et al. developed circDesign, an algorithmic platform for circRNA structural prediction and sequence design. This platform extends their previously established AI framework, originally designed for linear mRNA sequence engineering. The model initially generates diverse mRNA sequence libraries from target amino acid sequences, then filters the ORF regions of circRNAs based on minimal free energy and codon adaptation index criteria. It assembles optimized sequences into complete circRNA molecules, evaluates IRES positional entropy and structural ensemble diversity within these constructs, and ultimately validates the designed circRNAs through in vitro and in vivo screening platforms. Experimental validation demonstrated that algorithm‐designed circRNAs exhibited significantly extended half‐lives, indicating enhanced nuclease resistance. Moreover, the CR3 circRNA isoform showed the highest abundance in both transcriptomic and ribosome footprint profiling, highlighting its efficient translation capability [247].
Functional Prediction: Analysis of Interaction Networks
8.4.3
Current research on circRNA functional mechanisms primarily focuses on their interactions with proteins, making the prediction of circRNA–protein interaction sites and the assessment of their binding affinity increasingly crucial for advancing this field. Traditional experimental methods, such as CLIP‐seq, are limited by their ability to capture only single RBP binding sites within specific cellular contexts, often requiring lengthy and costly procedures [248]. Additionally, protein–RNA interactions dynamically change in response to alterations in the cellular microenvironment, while studies on protein‐mediated RNA regulation require binding information under consistent conditions [249].
Deep learning‐based prediction of circRNA–protein binding sites provides a viable alternative to overcome these economic and temporal constraints. CircRIP, a framework that integrates stacked codon‐based encoding and hybrid deep learning architectures, uses RNA sequences alone to predict RBP binding sites on circRNAs [250]. CSCRSites, another deep learning model, incorporates nucleotide positional information for the classification of cancer‐specific circRNA–RBP binding sites [251]. HCRNet, a novel end‐to‐end framework utilizing bidirectional encoder representations from transformers to contextualize circRNA sequences, demonstrates superior performance [252]. The recently proposed CRIECNN model, which combines a CNN, a BiLSTM layer, and a self‐attention mechanism for feature refinement, achieves a mean AUC of 0.9577 on benchmark datasets, showcasing excellent accuracy and performance in predicting circRNA–RBP interactions for both full‐length sequences and fragments [253]. Moreover, the recently introduced CoPRA model, developed through a multi‐institutional consortium, has gained significant attention for its innovative integration of protein language models and RNA language models within a sophisticated architecture designed for protein–RNA binding affinity prediction. Benchmark evaluations show that CoPRA achieves state‐of‐the‐art performance across various datasets, positioning it as a promising framework for future investigations into circRNA–RBP interactions [254].
Challenges in the Clinical Translation of CircRNAs
9
Challenges in the Standardized Study of CircRNAs
9.1
While the rapid advancement of high‐throughput sequencing technologies and algorithmic innovations has significantly accelerated circRNA research, these developments also introduce challenges in establishing standardized experimental and analytical frameworks. At the library preparation stage, there is considerable divergence between RNase R‐treated and untreated datasets. Critical digestion conditions, such as enzyme concentration, experimental time, and reaction temperature, significantly impact the results [255]. Additionally, specific RNA structures, including G‐quadruplexes, histone mRNAs, and small nuclear RNAs, exhibit intrinsic resistance to RNase R digestion [256, 257]. Furthermore, despite the availability of numerous computational algorithms for circRNA detection, there are substantial variations in sensitivity across different methods. A 2023 benchmarking study comparing 16 distinct algorithms revealed identification discrepancies ranging from 1372 to 58,032 circRNAs per algorithm within identical cell lines [256]. Consequently, integrating multiple algorithmic tools is essential. CircComPara2 incorporates up to eight circRNA detection tools alongside various analytical modules [258]; however, such multitool integration has not yet established itself as a universal gold‐standard computational framework, necessitating comprehensive benchmarking studies and algorithmic innovations.
Inherent differences in sequencing technologies also pose significant challenges for circRNA normalization. Conventional short‐read sequencing (second‐generation sequencing) generates reads typically ranging from 50 to 300 nucleotides in length [259], enabling high‐throughput data acquisition. However, these short reads are generally incapable of spanning the full BSJ, necessitating reliance on split‐alignment strategies to identify discontinuous sequences flanking the BSJ. This approach is associated with higher false‐positive rates and a dependency on existing reference genome annotations [260]. In contrast, long‐read sequencing (third‐generation sequencing) offers distinct advantages in detecting full‐length circRNAs and characterizing isoforms. This technology can directly span entire circRNA molecules to capture complete sequences, thereby enabling the detection of a greater number of alternative circularization events and internal alternative splicing events. However, long‐read sequencing faces technical limitations such as lower base‐call accuracy and read truncation, which may compromise the quantification of splice isoforms [261]. Additionally, the high costs associated with these technologies present challenges in standardizing research protocols.
Recent advancements address these issues through Longcell, an analytical pipeline developed at Stanford University School of Medicine. This framework incorporates four core technological innovations that achieve Spearman correlation coefficients exceeding 0.85 for single‐cell isoform quantification accuracy using nanopore long‐read data, positioning it as an indispensable analytical tool for long‐read sequencing applications [262]. Finally, discrepancies, redundancies, and inconsistencies in circRNA annotations (including IDs, sequences, genomic locations, host genes, and expression profiles) across public databases, coupled with the absence of standardized nomenclature, result in multiple designations for identical circRNAs, thus posing significant challenges for researchers integrating and cross‐referencing results from diverse sources.
Ethical Issues in the Clinical Translational Research of CircRNA
9.2
Clinical translational research serves as a crucial bridge between fundamental medical discoveries and front‐line clinical practice. Current efforts in circRNA translation primarily focus on diagnostic biomarkers, cancer immunotherapy, drug development, and innovative vaccine design. However, the field is still in its infancy, with most circRNA‐related studies remaining at the preclinical stage [74, 116, 263]. Consequently, there is an urgent need for standardized ethical frameworks to address the associated moral and regulatory challenges.
The most prominent application of circRNAs in clinical translation is in disease diagnosis and biomarker development. Due to their structural stability and abundance in peripheral blood, urine, and exosomes, circRNAs have been extensively studied as noninvasive biomarkers for gastrointestinal cancers, urological tumors, and other diseases [264, 265]. Nevertheless, privacy and data security have become critical concerns in the clinical implementation of circRNA‐based biomarkers, raising ethical issues about protecting sensitive patient information from unauthorized access and misuse [266, 267]. Therefore, obtaining informed consent before collecting biospecimens and associated data, such as pathological tissues, plasma, and urine, is essential. For biomarker research, the informed consent process should involve thorough consultations between patients and relevant institutions, such as hospitals and laboratories, rather than just a signature on a form. Given the rapidly evolving nature of circRNA research and the potential for future secondary use of samples, adopting a dynamic consent framework may better safeguard patient rights and autonomy [268]. Furthermore, the clinical translation of circRNAs in cancer immunotherapy and drug development faces substantial ethical and regulatory challenges, largely due to differing approval pathways across jurisdictions. In the United States, biological products require premarket approval from the United States Food and Drug Administration (US FDA). In Europe, advanced therapy medicinal products, including gene‐, tissue‐, and cell‐based therapies, are regulated under regulation (EC) No. 1394/2007 and must obtain centralized marketing authorization from the European Medicines Agency [269]. In China, the National Medical Products Administration oversees the development, production, distribution, and use of pharmaceuticals, including biological drugs [270]. Navigating innovative therapies through these diverse regulatory frameworks presents significant challenges, necessitating continuous adaptation through sustained dialogue with all stakeholders.
Moreover, due to the unique survival characteristics of cancer patients, most current circRNA‐based therapeutic regimens have yet to demonstrate improvements in long‐term survival, leaving considerable uncertainty regarding their enduring benefits and risks. To ensure participant safety, it is essential to establish unified clinical translation protocols through multinational, multicenter collaborations [271].
Conclusions and Perspective
10
Initially, circRNAs were considered mere “noise” in the translation process. However, with technological advancements, circRNAs have been found to have multiple functions, including acting as miRNA sponges, regulating parental gene transcription, and serving as protein translation templates [272]. These diverse functions are associated with various physiological and pathological processes. Due to their unique structural characteristics, circRNAs exhibit stable in vivo expression and tissue specificity, making them potential biomarkers [101, 273].
Elucidating the detailed biological functions of circRNAs requires high‐precision spatial mapping of their subcellular localization. Super‐resolution microscopy techniques, such as stochastic optical reconstruction microscopy and photoactivated localization microscopy, have become the primary methodologies for subcellular circRNA localization and functional characterization [274, 275]. However, super‐resolution imaging demands sophisticated instrumentation and meticulous calibration, leading to significant economic and temporal costs. This highlights the need for technological refinements and cost‐containment strategies. Multimodal imaging, which integrates optical microscopy with mass spectrometry imaging, offers a promising approach by enabling concurrent mapping of circRNA spatial localization and interactions, thus providing comprehensive insights into biological processes from molecular to tissue levels. Importantly, these techniques generate vast datasets alongside high‐precision images, necessitating the development of advanced machine and deep learning algorithms to optimize data processing, automate analysis pipelines, and enable real‐time interpretation of imaging data.
Given their superior structural integrity compared with mRNAs and their pleiotropic functional modalities in vivo, circRNAs are promising candidates for engineered therapeutic vectors. Although current technologies can synthesize both short and long circRNA sequences, previous studies have shown that RNAs generated through different circularization methods can elicit markedly distinct immune responses in humans [16, 276]. This underscores the need for context‐specific circRNA synthesis platforms to ensure predictable outcomes across diverse biological applications. Additionally, circRNA delivery poses significant challenges. While LNP systems are established delivery platforms, their encapsulation efficiency for circRNAs is substantially lower than for linear mRNAs due to the larger molecular size of circular transcripts. Multiple biotechnology firms are optimizing LNP formulations to enhance delivery efficacy [277]. Targeted delivery is another critical hurdle, as the inherent hepatotropism of LNPs limits therapeutic applications to hepatic tissues, whereas most diseases require delivery to extrahepatic organs [278]. Consequently, developing exosomes, which are naturally evolved intercellular communication vehicles, as delivery vectors represents a compelling alternative. Several companies are now focusing on exosome‐based delivery technologies, despite the engineering complexities and substantial manufacturing costs.
Over the past decade, AI has emerged as a powerful tool in biomedical research, particularly with the advent of large language models like ChatGPT and Claude, alongside advanced deep learning algorithms. Conventional approaches for screening and validating therapeutic circRNA candidates typically require months to years, whereas AI‐driven pipelines enhance efficiency by over 50%. Chinese research teams have made substantial advancements in circRNA vaccine development through AI‐driven multimodal optimization [279]. Future efforts necessitate expanded translational investigations into circRNA‐based therapeutics across diverse oncological contexts. Integrating multiomics data, including single‐cell sequencing, epigenomic profiles, and protein interaction networks, through AI frameworks enables robust prediction of circRNA targets with therapeutic potential. However, fragmented medical data across isolated databases and stringent privacy constraints create significant data barriers, impeding AI model development due to prohibitive training costs. Establishing curated, publicly accessible multiomics repositories spanning diverse diseases is therefore imperative. Furthermore, the opaque decision‐making processes of deep neural networks necessitate rigorous validation of model interpretability through SHapley Additive exPlanations frameworks and experimental verification.
Collectively, circRNAs demonstrate superior structural and functional attributes in oncology research, driving a paradigm shift from traditional biomarkers, anticancer therapies, and mRNA vaccines toward multimodal circRNA‐based therapeutics. In Q4 2024, the US FDA granted Investigational New Drug (IND) clearance for the Phase I/IIa RXRG001 trial (SPRINX‐1 study), representing the first US FDA‐approved circRNA therapy globally. Furthermore, China's Center for Drug Evaluation (CDE) accepted the IND application for BioCrown's circRNA drug TI‐0093 in June 2025. This investigational injection targets HPV16‐positive advanced recurrent or metastatic solid tumors, constituting the only circRNA antitumor agent in IND‐stage development. Sustained research investment and substantial funding remain imperative to ensure translational feasibility.
Author Contributions
Zehao Ding, Zai Luo, and Liao Zhang conceptualized the manuscript, performed the literature search, wrote the original draft, and revised and edited the manuscript. Shaopeng Zhang revised and edited the manuscript. Renchao Zhang, Zhengjun Qiu, and Chen Huang supervised, revised, and edited the manuscript. All authors read and approved the final manuscript.
Funding Information
CH is supported by the National Natural Science Foundation of China (No.82472921 and No. 82072662), Key Projects of the Interdisciplinary Program of Medicine and Engineering at Shanghai Jiao Tong University (No. 24×010301419), and the 2021 Shanghai ‘Rising Stars of Medical Talent’ Youth Development Program: Outstanding Youth Medical Talents. ZL is sponsored by Shanghai Rising‐Star Program Sailing Program (No. 24YF2734700) and Young Scientists Fund of the National Natural Science Foundation of China (Grant No. 82504100).
Ethics Statement
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1W. Li , J.‐Q. Liu , M. Chen , et al., “Circular RNA in Cancer Development and Immune Regulation,” Journal of Cellular and Molecular Medicine 26, no. 6 (2020): 1785–1798.33277969 10.1111/jcmm.16102 PMC 8918416 · doi ↗ · pubmed ↗
- 2D. Kolakofsky , “Isolation and Characterization of sendai Virus DI‐RN As,” Cell 8, no. 4 (1976): 547–555.182384 10.1016/0092-8674(76)90223-3 · doi ↗ · pubmed ↗
- 3B. P. Nicolet , S. Engels , F. Aglialoro , V. D. Akker E , V. Lindern M , and M. C. Wolkers , “Circular RNA Expression in human Hematopoietic Cells Is Widespread and Cell‐type Specific,” Nucleic Acids Research 46, no. 16 (2018): 8168–8180.30124921 10.1093/nar/gky 721PMC 6144802 · doi ↗ · pubmed ↗
- 4K. Nemeth , R. Bayraktar , M. Ferracin , et al., “Non‐coding RN As in Disease: From Mechanisms to Therapeutics,” Nature Reviews Genetics 25, no. 3 (2023): 211–232.10.1038/s 41576-023-00662-137968332 · doi ↗ · pubmed ↗
- 5I. B. Filippenkov , E. O. Kalinichenko , S. A. Limborska , and L. V. Dergunova , “Circular RN As‐one of the Enigmas of the Brain,” Neurogenetics 18, no. 1 (2016): 1–6.27449796 10.1007/s 10048-016-0490-4 · doi ↗ · pubmed ↗
- 6G. Huang , M. Liang , H. Liu , et al., “Circ RNA hsa_circ RNA_104348 Promotes Hepatocellular Carcinoma Progression Through Modulating mi R‐187‐3p/RTKN 2 Axis and Activating Wnt/β‐catenin Pathway,” Cell Death & Disease 11, no. 12 (2020): 1065.33311442 10.1038/s 41419-020-03276-1PMC 7734058 · doi ↗ · pubmed ↗
- 7F. Yang , Q. Ma , B. Huang , et al., “Circ NFATC 3 promotes the Proliferation of Gastric Cancer Through Binding to IGF 2BP 3 and Restricting Its Ubiquitination to Enhance CCND 1 m RNA Stability,” Journal of Translational Medicine 21, no. 1 (2023): 402.37340423 10.1186/s 12967-023-04235-y PMC 10280940 · doi ↗ · pubmed ↗
- 8B. Fu , W. Liu , C. Zhu , et al., “Circular RNA circ BCBM 1 Promotes Breast Cancer Brain Metastasis by Modulating mi R‐125a/BRD 4 Axis,” International Journal of Biological Sciences 17, no. 12 (2021): 3104–3117.34421353 10.7150/ijbs.58916 PMC 8375234 · doi ↗ · pubmed ↗