Public health surveillance of Vibrio cholerae in travellers returning to the United Kingdom
Ching-Ying J. Poh, David R. Greig, Ella V. Rodwell, Claire Jenkins

TL;DR
This study analyzed Vibrio cholerae cases in UK travelers to understand its spread and risk factors, highlighting the need for better surveillance and global cooperation.
Contribution
The study provides a detailed UK-based analysis of V. cholerae isolates from travelers, identifying key trends and recommending improved surveillance strategies.
Findings
Cholera toxin-producing ST69 was the most common sequence type among isolates.
Travel restrictions during the pandemic led to a significant drop in reported cases in 2020 and 2021.
India, Pakistan, and Kenya were the most frequently reported travel destinations for V. cholerae cases.
Abstract
Introduction. Vibrio cholerae is a diverse species of bacteria that causes watery diarrhoea, vomiting and stomach cramps and is the aetiological agent of cholera. Gap statement. Despite the global upsurge in notifications of cholera and concerns over the impact of climate change, systematic analysis of national and international surveillance data describing the microbiology and epidemiology of V. cholerae is sparse. Aim. We reviewed the microbiology and epidemiology of V. cholerae isolated from travellers returning to the UK. Methodology. All human isolates of V. cholerae detected from 2004 to 2024 were extracted from UK Health Security Agency reference laboratory surveillance systems. Microbiological data were analysed and linked to available epidemiological data and genome sequences for all isolates from 2016 to 2024. Results. There were 984 notifications of V. cholerae from 2004…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
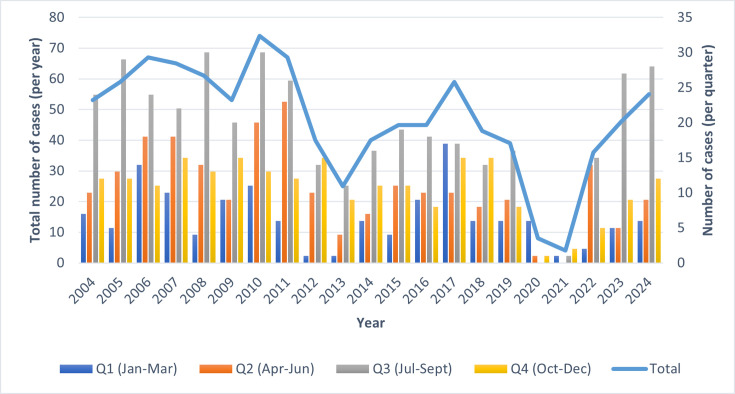 Fig. 1
Fig. 1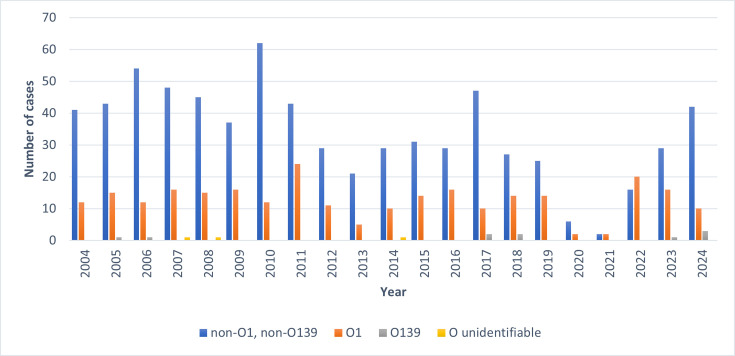 Fig. 2
Fig. 2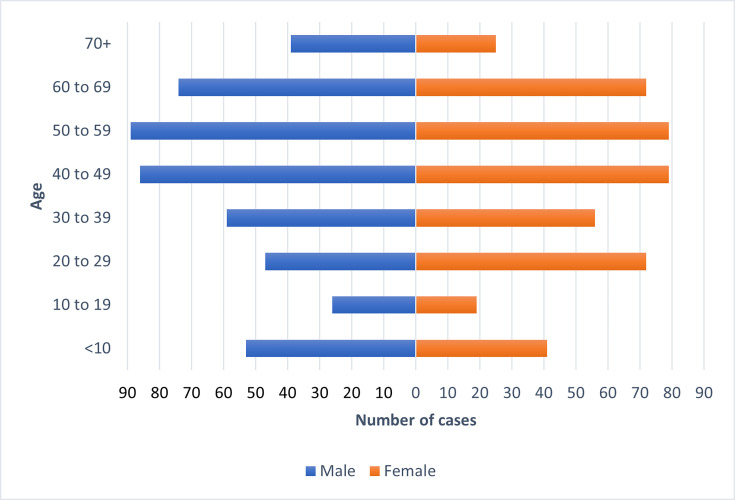 Fig. 3
Fig. 3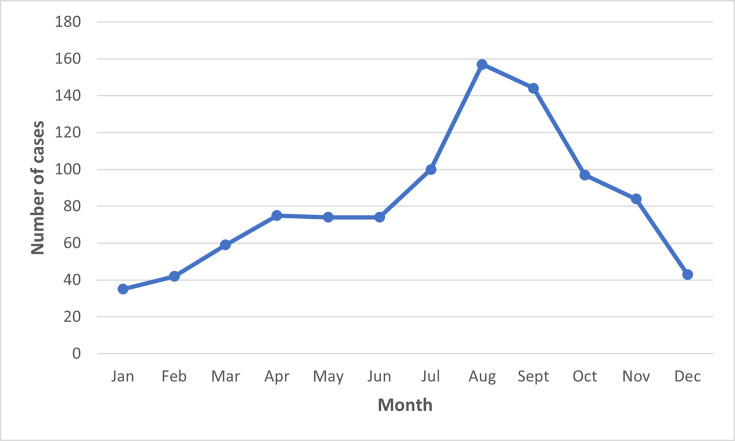 Fig. 4
Fig. 4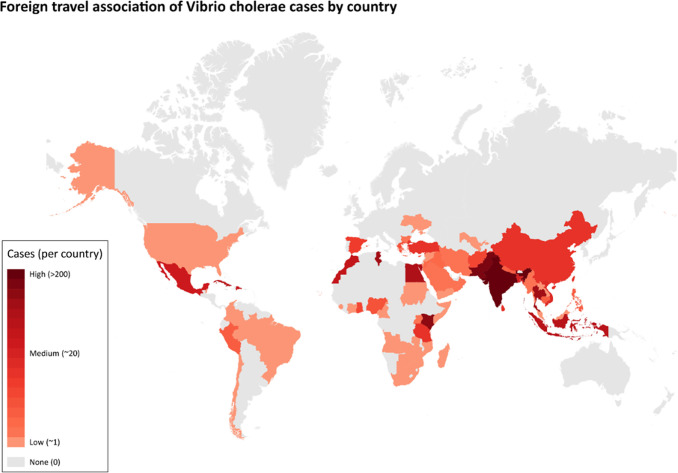 Fig. 5
Fig. 5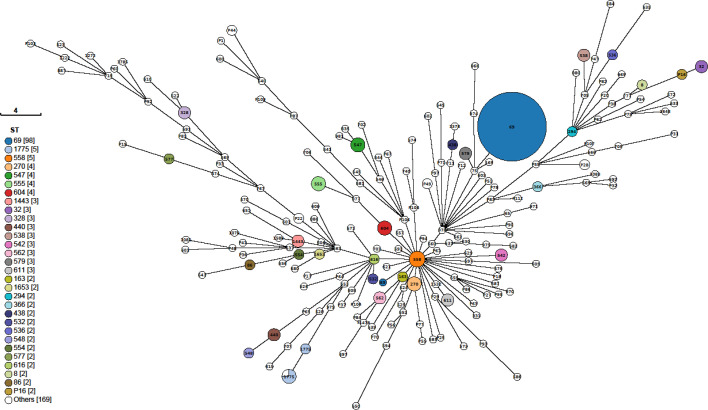 Fig. 6
Fig. 6| UKHSA region | UKHSA centre | Cases (%) |
|---|---|---|
| London | London | 108 (11) |
| Midlands and East of England | East Midlands | 67 (6.8) |
| East of England | 92 (9.4) | |
| West Midlands | 56 (5.7) | |
| North of England | North East | 58 (5.9) |
| North West | 144 (14.7) | |
| Yorkshire and Humber | 111 (11.3) | |
| South of England | South East | 220 (22.4) |
| South West | 123 (12.6) | |
| Wales* | – | 1 (0.1) |
| Continent | No. (%) |
|---|---|
| Africa | 243 (24.7) |
| Asia | 564 (57.3) |
| Europe | 23 (2.3) |
| North America | 62 (6.3) |
| South America | 7 (0.7) |
| Y – not stated | 15 (1.5) |
| None | 14 (1.4) |
| Unknown | 56 (5.7) |
|
|
|
- —http://dx.doi.org/10.13039/100018336 National Institute for Health Research Health Protection Research Unit
- —http://dx.doi.org/10.13039/100018336 National Institute for Health Research Health Protection Research Unit
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Travel-related health issues · Yersinia bacterium, plague, ectoparasites research
Introduction
Vibrio cholerae is a diverse species of bacteria that causes symptoms ranging from mild to watery diarrhoea, vomiting and stomach cramps and is the aetiological agent of cholera. Cholera is an acute gastrointestinal infection where patients pass large volumes of stool that can resemble rice water, leading to severe dehydration that can kill within hours if left untreated [12]. Cholera is caused by cholera toxin produced by the prophage-encoded ctx genes, which are present in a limited number of V. cholerae serogroups, most commonly serogroup O1 [3]. There are over 200 different lipopolysaccharide O antigens, or serogroups, of *V. cholerae *[4]. Although the non-O1 serogroups are associated with a milder form of gastroenteritis, extraintestinal infections can occur, including sepsis, wound infections and otitis media [57].
Clinical management of cholera involves oral rehydration therapy, supplemented by treatment with antimicrobials, if required to reduce the severity and duration of the disease [8]. Empirical treatment includes tetracycline, fluoroquinolones, macrolides, co-trimoxazole and, less commonly, extended spectrum beta-lactams [9]. The emergence of antimicrobial resistance in V. cholerae has led to increasing global public health concern and poses new challenges in managing cholera treatment [1011].
Humans become infected with V. cholerae via contaminated water, and limited access to clean water and/or sanitation facilities is a driver of transmission. The consequences of a humanitarian crisis, such as the disruption of water and sanitation systems, or the displacement of populations to inadequate and overcrowded camps, can increase the risk of outbreaks. It is estimated that there are 1.3 to 4.0 million cases and 21,000 to 147,000 deaths due to cholera each year [12]. V. cholerae has caused seven pandemics of cholera (the seventh is on-going) and remains a significant global threat to public health [13]. Cholera is endemic across Africa and Asia, and outbreaks are common, resulting in a large healthcare burden in these regions [1415].
The UK Standards for Microbiology Investigations for Gastroenteritis recommends testing faecal specimens for V. cholerae in individuals with gastroenteritis reporting recent travel (within 2 to 3 weeks) to countries where cholera is endemic, patients with suspected cholera and/or individuals reporting epidemiological links to outbreaks caused by the consumption of contaminated seafood (Gastroenteritis S7 Syndromic Documents) [16]. Consequently, the true incidence of domestically acquired V. cholerae infection in the UK remains unknown, and almost all isolates of enteric origin are associated with travellers' diarrhoea. There is no evidence that the pandemic serogroup V. cholerae O1 is endemic in the UK, although non-O1 serogroups have been detected in UK rivers and coastal waters [17].
In 2018, the UK Health Security Agency (UKHSA) implemented whole-genome sequencing (WGS) for the routine surveillance of V. cholerae [18]. The aim of this study was to review the microbiology and epidemiology characteristics of V. cholerae isolated from travellers returning to the UK.
Methods
Epidemiological data
Patient information, including sex, age and recent travel, was collected from laboratory request forms upon submission. These data were stored in the Gastro Data Warehouse, an in-house UKHSA database for storing and linking patient demographic and microbiological typing data. Data on symptoms were limited, stating only that the patient had either gastrointestinal symptoms or an extraintestinal infection. There were no data on the severity of symptoms or patient outcomes. A case was defined as an individual that had V. cholerae isolated from their faecal specimen or an extraintestinal site.
Microbiology
Faecal specimens submitted for microbiological testing from individuals hospitalized and community cases with symptoms of gastrointestinal infection reporting travel within 4 weeks to endemic regions were tested for V. cholerae. Positive cultures are referred to the Gastrointestinal Bacteria Reference Unit (GBRU) for confirmation and typing. Prior to 2016, phenotypic identification was performed by inoculating a combination of over 40 biochemical substrates. Utilization of the substrate was identified by a colour change or gas production within the media. The ability (or inability) of the bacteria to utilize the substrates was recorded as a positive (or negative) result and compared with that of published tables categorizing the known reactions of Vibrio species. Isolates of V. cholerae were agglutinated with antisera raised to O1 (Ogawa and Inaba) and O139 (Bengal) antisera to determine the serogroup.
Whole-genome sequencing
All viable cultures of V. cholerae submitted to the GBRU between January 2016 and December 2024 were sequenced (see Table S1, available in the online version of this article). Genomic DNA was extracted and indexed using Nextera XT DNA and Nextera Flex sample preparation kits (Illumina) and sequenced using the Illumina HiSeq 2500 and NextSeq 1000 platforms at UKHSA. Trimmomatic (v0.36) [19] was used to trim sequence adapters. Reads with a PHRED score below 30 and a read length of less than 50 bp were discarded, along with their paired reads. FASTQ reads from all sequences in this study can be found at the UKHSA Pathogens BioProject at the National Center for Biotechnology Information (BioProject number PRJNA438219).
Sequence typing and virulence profiling
Sequence type (ST) assignment was performed using MOST v2.18 [20] https://github.com/phe-bioinformatics/MOST. Any multilocus sequence type (MLST) gene sequences that did not match the existing alleles were submitted to pubMLST (see https://pubmlst.org/vcholerae/) for assignment of a new allelic type. Similarly, new allelic profiles were also submitted to the database for assignment of a new ST. GrapeTree was employed to generate a minimum spanning tree (MSTree-V2). This was visualized in the GrapeTree platform and annotated with ST derived from SRST2.
Using GeneFinder v2.9 (https://github.com/ukhsa-collaboration/gene_finder) which utilizes Bowtie2 v2.1 [21] to perform sequence alignment, FASTQ reads were mapped to the virulence regulator gene toxR (GenBank accession no. [KF498634](https://www.ncbi.nlm.nih.gov/nuccore/KF498634)), the cholera toxin gene ctxA (GenBank accession no. [AF463401](https://www.ncbi.nlm.nih.gov/nuccore/AF463401)), wbeO1 and wbfO139 (GenBank accession numbers [KC152957](https://www.ncbi.nlm.nih.gov/nuccore/KC152957) and [AB012956](https://www.ncbi.nlm.nih.gov/nuccore/AB012956)) encoding the somatic O antigens O1 and O139, tcpA classical and tcpA El Tor gene sequences (GenBank accession numbers [M33514](https://www.ncbi.nlm.nih.gov/nuccore/M33514) and [KP187623](https://www.ncbi.nlm.nih.gov/nuccore/KP187623)). The best match to each target was reported with metrics, including coverage, depth and nucleotide similarity in XML format for quality assessment. toxR is found in all V. cholerae isolates and is regarded as a marker for species identification. The ctxA encoding cholera toxin is most often, although not exclusively, associated with V. cholerae serotypes O1 and O139 and is a characteristic of these pandemic lineages [22]. Variants of tcpA can be used to identify the classical and El Tor biotypes [22]. For in silico predictions, only results that matched to a gene determinant at >80% nucleotide identity over >80% target gene length were accepted.
Results
Overview of the dataset
In total, there were 985 isolates of V. cholerae isolated from 984 patients presenting with symptoms of diarrhoeal illness in England submitted to the GBRU between January 2004 and December 2024 (Fig. 1). From 2004 to 2024 (excluding 2020 and 2021), on average, 51 confirmed V. cholerae isolates are referred to GBRU for identification and typing each year. The greatest number of isolates was received in 2010 (n=74), while the lowest number was received in 2020 (n=8) and 2021 (n=4) due to travel restrictions imposed because of the COVID-19 pandemic. Of the 12 cases in 2020 and 2021, 9 were reported either before the travel restrictions were imposed in March 2020 or after the restrictions were relaxed in May 2021. The three cases identified during the travel ban were V. cholerae non-O1 serogroups; they did not report recent travel outside the UK, and for two of the patients, V. cholerae was isolated from extraintestinal sites, including a skin swab and a tracheal aspirate.
Number of V. cholerae cases (organized by quarter) referred to the Gastrointestinal Bacteria Reference Unit between January 2004 and December 2024 (n=984). Coloured bars represent different quarters per year. Solid line represents total annual cases.
The majority of isolates belonged to the non-O1, non-O139 group (n=706, 71.7%), and 266 (27.0%) belonged to the O1 serogroup (Fig. 2). V. cholerae O139 was uncommon, with only 10 (1.0%) cases. There were three cases (0.3%) where the serogroup could not be determined and were categorized as ‘O unidentifiable’. For V. cholerae O1 cases (n=266), the majority had the El Tor biotype (n=235, 88.3%), while there were 31 cases (11.7%) with no identified biotype. There were no V. cholerae O1 cases with the classical biotype.
Number of V. cholerae cases (categorized by serogroup) referred to the Gastrointestinal Bacteria Reference Unit between January 2004 and December 2024 (n=984).
Descriptive epidemiology
The age–sex distribution of all V. cholerae cases included in this study where age and sex data were available (n=916/984=93%) indicated a balanced sex ratio, although the number of male cases was greater (n=473, 51.6%) compared to female (n=443, 48.4%). The median age for females was 45 years (IQR: 26–58), while the median age for males was 46 years (IQR: 28–59). Additionally, the highest proportion of cases belonged to the 50 to 59 age group for males and females combined (n=168, 18.3%), including for females only (n=79, 8.6%) and males only (n=89, 9.7%). The second highest proportion of cases was observed in the 40 to 49 age group (n=165, 18.0%). The lowest proportion of cases was observed in the 10 to 19 age group for males and females combined (n=45, 4.9%), males only (n=26, 2.8%) and females only (n=19, 2.1%) (Fig. 3).
Age–sex distribution of total V. cholerae cases reported to UKHSA [isolates originating from England (n=916), where date of birth, sample date and sex were available].
Seasonal variation in the number of reported V. cholerae cases was observed, with incidence highest between the summer months of July and September (Fig. 1), forming a late summer peak in August (n=157) and lower levels during off-peak seasons from December to February (Fig. 4).
The number of V. cholerae cases (n=984) by months between 2004 and 2024 in England.
V. cholerae cases were detected in all regions of England and one in Wales, with the South of England having the highest frequency (n=343, 35.0%) – South East (n=220); South West (n=123). The North of England had the second highest frequency of cases (n=313, 31.9%), specifically the North West UKHSA centre (n=144) and Yorkshire and Humber (n=111). Excluding the Wales case, the lowest number of reported cases was in the West Midlands (n=56, 5.7%) and the North East of England (n=58, 5.9%) (Table 1, Fig. 5).
V. cholerae cases associated with foreign travel outside of the UK, where a specific country was stated (n=868). Countries are colour-coded in proportion to the frequency of cases associated, i.e. the higher the frequency of cases, the darker the colour.
The geographical distribution of cases typically clustered around major cities, notably, London, Manchester, Newcastle and Leeds (Fig. 5). One case resided in Wales, while another case resided in the Isle of Man (not shown).
Of the 984 cases, 914 (92.9%) were associated with foreign travel, 14 (1.4%) were not associated with foreign travel, and foreign travel was unknown for 56 cases (5.7%). Overall, foreign travel was most commonly associated with Asia (n=564, 57.3%) and Africa (n=243, 24.7%) (Table 2). The top three travel destinations were India (n=239), Pakistan (n=135) and Kenya (n=68) (Fig. 6).
Minimum spanning tree describing the 7-gene MLST of V. cholerae samples submitted to UKHSA between 2015 and 2024, where MLST profiles were available (n=348). Annotated with labels to describe the ST, and coloured to reflect each ST.
Population structure of V. cholerae isolated in the UK from returning travellers
Sequence typing data were available for 351 isolates, of which 229 isolates belonged to the non-O1, non-O139 serogroup, 115 isolates were O1, and 7 were O139. Of these 351 isolates, 66 (18.8%) were novel STs and were assigned either a provisional ST (pST) or were designated as ‘novel ST’. Overall, ST69 was the predominant ST (n=99, 28.2%), followed by ST1775 (n=7, 2.0%) and ST558 (n=5, 1.4%)Fig. 7. There were 180 different STs associated with the non-O1, non-O139 serogroup, while 7 STs were associated with the O139 serogroup (ST163, ST544, ST586, ST707, ST1222, pST93 and pST103). There were 14 different STs associated with the O1 serogroup, of which ST69 was the predominant ST (n=98) (Table S1).
Of the 351 isolates with WGS typing data, 104 (29.6%) belonged to CC69. Genome-derived MLST indicated that CC69 isolates were consisted of four STs, with the majority of CC69 isolates belonging to ST69 (n=99/104, 95.2%). Additional STs included ST579 (n=3/104, 2.9%), ST75 (n=1/104, 1.0%) and ST169 (n=1/104, 1.0%). Of the CC69 isolates, 43 (41.3%) were associated with foreign travel to Pakistan, 26 (25.0%) to India, and 7 (6.7%) to Bangladesh.
Discussion
In the UK, the majority of faecal specimens from patients presenting to primary healthcare with gastrointestinal symptoms are examined for V. cholerae only if they report recent travel to countries where cholera is endemic (Gastroenteritis S7 Syndromic Documents). The decline in notifications of V. cholerae in England that started in 2011 was most likely linked to a reduction in travel to endemic regions, possibly due to political unrest in certain countries [23]. The number of cases increased in 2014 and remained stable until 2019. In 2020, notifications fell sharply, and this decline in notifications was linked to the travel restrictions put in place during the COVID-19 pandemic. Once the travel restrictions were lifted, notifications quickly returned to pre-pandemic levels [24].
Based on global surveillance data from 2022, the World Health Organization reported a global upsurge of cholera caused by V. cholerae O1, with more cases reported from an increasing number of countries [1025]. There were more outbreaks, notably in Afghanistan, Cameroon, the Democratic Republic of Congo, Malawi, Nigeria, Somalia, Syria and Yemen, and the size of the outbreaks also increased [15,2631]. An upsurge of V. cholerae O1 was also captured in European surveillance systems [10], and in the trends analysis in this study, where the number of notifications of V. cholerae O1 in 2022 was higher than they had been for over 10 years. This increase may be due in part to the relaxation of travel restrictions providing the opportunity for European nationals to visit family and friends living in endemic regions, following the hiatus caused by the COVID-19 pandemic.
Although, in the UK, the notifications of V. cholerae O1 have declined over the last 2 years, the notifications of non-O1 V. cholerae – which includes V. cholerae O139 and V. cholerae non-O1, non-O139 – have continued to increase, with numbers in 2024 at the highest they have been since 2017. Undoubtedly, transmission of V. cholerae is linked to the lack of adequate safe water and sanitation due to underdevelopment, poverty and conflict [29]. However, national and international agencies, including the WHO, the European Food Safety Authority (EFSA) and the Food Standards Agency (FSA) in the UK, have all highlighted concerns that climate change may be a contributing factor. There is good evidence that extreme weather events, such as floods, droughts and cyclones, trigger new outbreaks and worsen existing ones [23233]. Warmer waters are also likely to result in increased Vibrio species that accumulate in fish, shellfish and crustaceans [73435].
Vibrio species have been associated with foodborne illness when contaminated shellfish are consumed raw or lightly cooked [36 37, 38, ]. Individuals with a weak immune system are at risk from V. cholerae O1 and non-O1 serotypes [3942]. Although shellfish are not currently routinely screened for Vibrio species by the food industry, FSA investigated five UK incidents involving Vibrio in shellfish products reported during 2022 and 2023. Four of the shellfish products were imported, while the fifth incident was the first report of V. cholerae from shellfish sourced from UK waters since records began [45, 43]. However, there have been no Vibrio-related foodborne outbreaks reported in the UK in recent times (UKHSA in-house data).
Analysis of the population structure reveals that V. cholerae is a diverse species. The presence of cholera toxin gene ctxA is almost exclusively associated with V. cholerae O1. A notable exception was the strain of V. cholerae serogroup O139 that emerged in the Bay of Bengal region in the early 1990s [44]. However, since that time, notifications of V. cholerae O139 have declined globally, and the isolates V. cholerae O139 detected in this study were negative for ctxA. Although almost all V. cholerae O1 ctxA isolates belonged to ST69, the non-O1 non-toxin-producing serogroups belonged to a large number of different STs, with very few STs comprising more than one isolate. Within the STs that comprise multiple isolates, the associated cases report travel locations in Africa, Asia, the Americas and Europe, indicating that common STs are widely geographically dispersed. Other studies have described the diversity of travel-associated non-pandemic V. cholerae [445].
The age–sex distribution analysis in this study indicates that in UK residents, children under the age of ten and the middle-aged group are most susceptible to infection and/or most at risk of experiencing severe clinical outcomes requiring medical care. The peak of case numbers observed in August and September is most likely due to increased travel during the summer holiday period and the impact of warmer climatic conditions in the northern hemisphere. Most notifications were from cases reporting travel to India or Pakistan, reflecting both the high incidence of V. cholerae associated with the Indian subcontinent and the proportion of UK nationals travelling to this region each year, compared to other endemic regions.
It is universally acknowledged that the key to prevention of cholera is proper investment in water, sanitation and hygiene (WASH); however, surveillance is necessary to assess the impact of WASH initiatives [29]. Accurate diagnostics and early case detection, integrated into strong surveillance systems, are essential tools for the management and control of cholera epidemics. Local investment in genomic surveillance is crucial to determine the origins of cholera outbreaks, monitor the spread of the disease and inform public health approaches to the control of the disease, particularly with regard to cross-border and international collaboration, to prevent ongoing transmission [29]. The WHO recommends that surveillance of V. cholerae should be part of an integrated disease surveillance system that includes feedback at the local level and information sharing at the global level [46]. To support these initiatives, it is UKHSA policy to make genome sequencing data linked to V. cholerae surveillance generated in the UK publicly available in real-time [47].
Closer to home, there is an increased risk of domestic acquisition of V. cholerae. Due to climate change, UK waters have become progressively warmer over the past 100 years, with average winter temperatures in particular increasing over the past 20 years [48]. We recommend widespread molecular testing of shellfish and more comprehensive testing of faecal specimens from non-travellers with gastrointestinal symptoms to monitor the emergence of V. cholerae in UK waters and ensure safety in the food chain.
Supplementary material
10.1099/jmm.0.002121Uncited Table S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Harris JB La Rocque RC Qadri F Ryan ET Calderwood SB Cholera Lancet 20123792466247610.1016/S 0140-6736(12)60436-X 22748592 PMC 3761070 · doi ↗ · pubmed ↗
- 2Kupferschmidt K A time of cholera Science 20233811391–139110.1126/science.adl 041437769103 · doi ↗ · pubmed ↗
- 3Mutreja A Dougan G Molecular epidemiology and intercontinental spread of cholera Vaccine 202038 Suppl 1A 46A 5110.1016/j.vaccine.2019.07.03831345641 · doi ↗ · pubmed ↗
- 4Bote L Taylor-Brown A Maes M Ingle DJ Valcanis M et al Surveillance of travel-associated isolates elucidates the diversity of non-pandemic Vibrio cholerae Microb Genom 2024103941287110.1099/mgen.0.001307 PMC 1190082839412871 · doi ↗ · pubmed ↗
- 5Engel MF Muijsken MA Mooi-Kokenberg E Kuijper EJ van Westerloo DJ Vibrio cholerae non-O 1 bacteraemia: description of three cases in the Netherlands and a literature review Euro Surveill 2016212710423710.2807/1560-7917.ES.2016.21.15.3019727104237 · doi ↗ · pubmed ↗
- 6Rehm C Kolm C Pleininger S Heger F Indra A et al Vibrio cholerae-an emerging pathogen in Austrian bathing waters?Wien Klin Wochenschr 202313559760810.1007/s 00508-023-02241-037530997 PMC 10651712 · doi ↗ · pubmed ↗
- 7Baker-Austin C Oliver JD Alam M Ali A Waldor MK et al Vibrio spp. infections Nat Rev Dis Primers 20184810.1038/s 41572-018-0005-830002421 · doi ↗ · pubmed ↗
- 8Sousa FBM Nolêto I Chaves LS Pacheco G Oliveira AP et al A comprehensive review of therapeutic approaches available for the treatment of cholera J Pharm Pharmacol 2020721715173110.1111/jphp.1334432737883 · doi ↗ · pubmed ↗