Balitora tiandengensis (Teleostei, Balitoridae), a new species of cave-dwelling loach from Guangxi, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
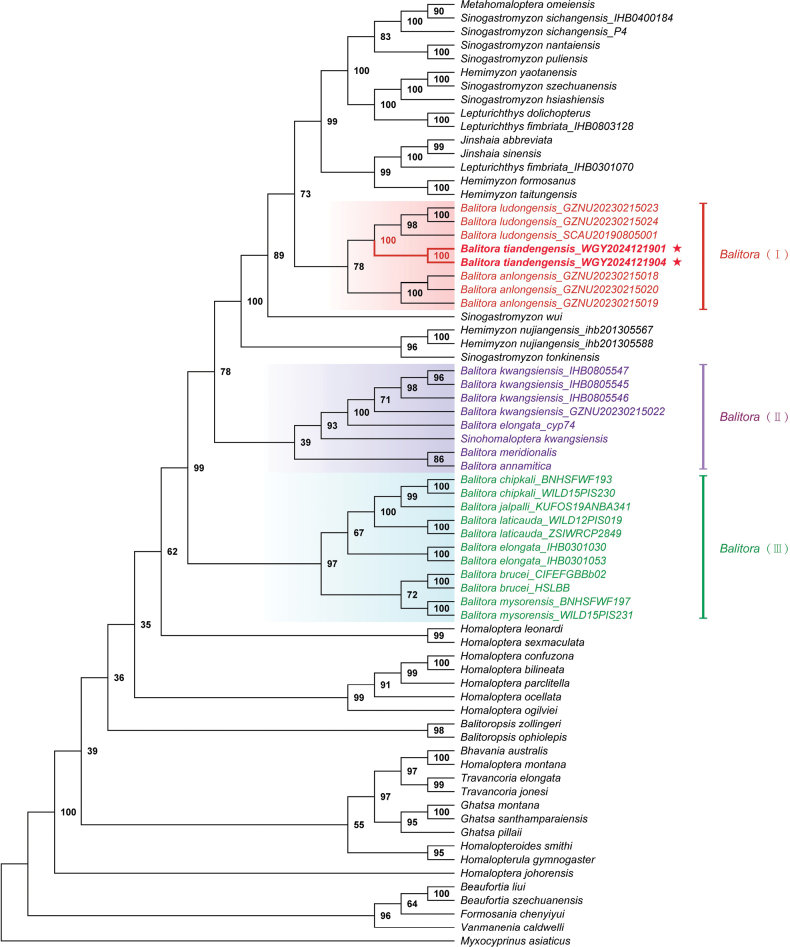 Figure 1
Figure 1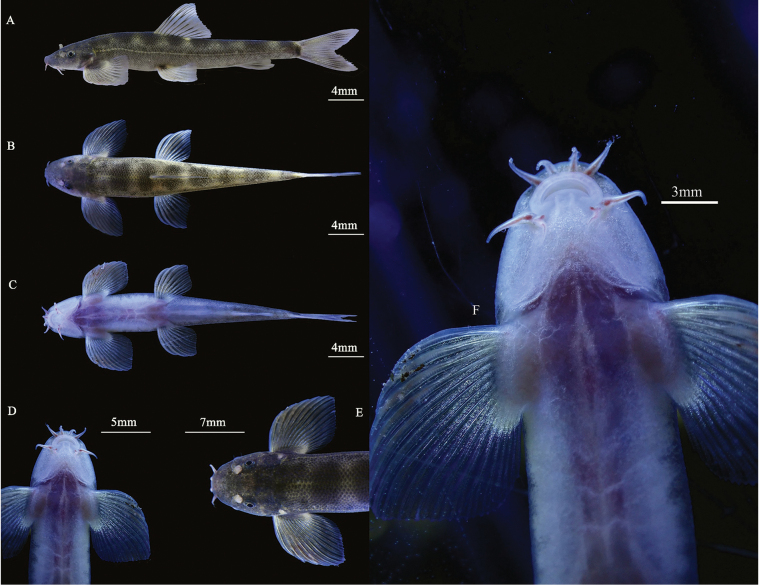 Figure 2
Figure 2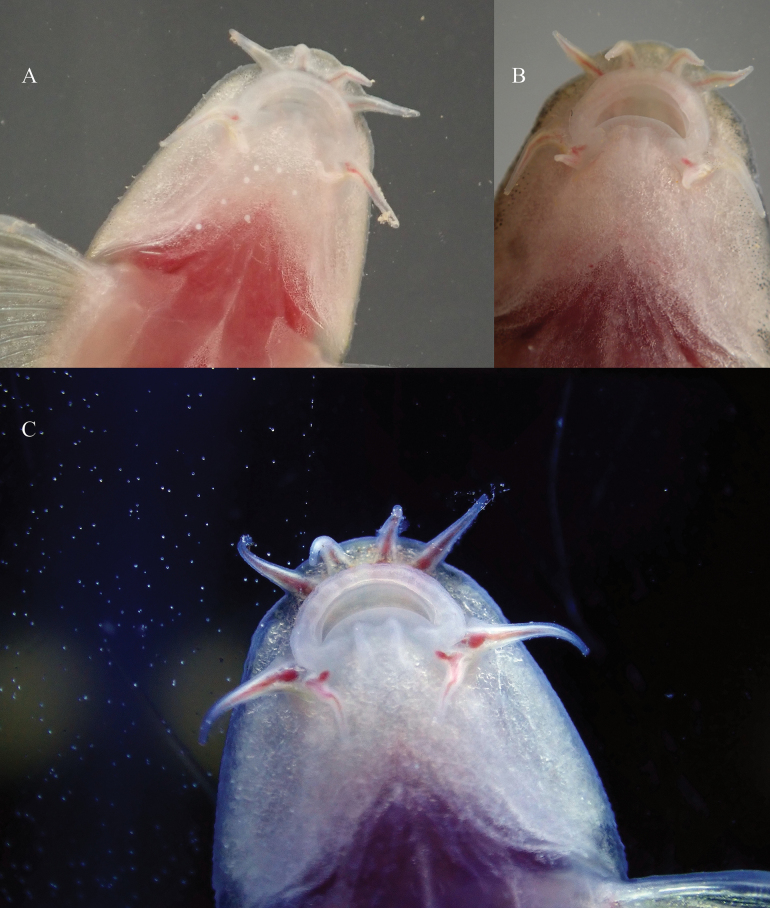 Figure 3
Figure 3 Figure 4
Figure 4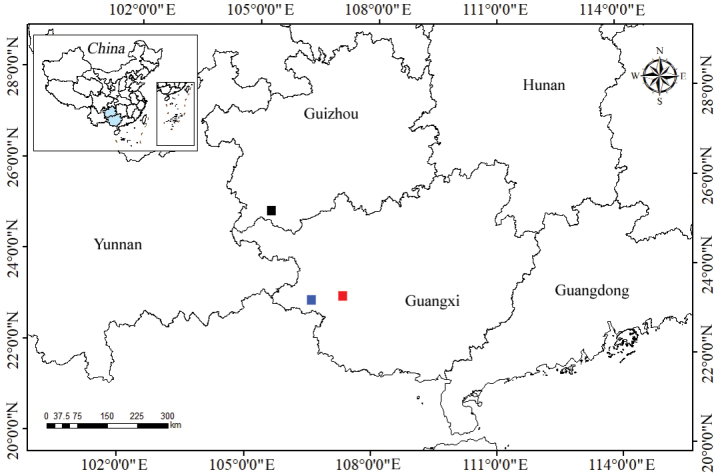 Figure 5
Figure 5| ID | Species | Locality (* type localities) | Voucher |
|
|
|
|
|
|---|---|---|---|---|---|---|---|---|
| 1 |
| Xinglong Town, Anlong County, Guzihou, China | GZNU20230215018 |
|
|
|
|
|
| 2 |
| Xinglong Town, Anlong County, Guzihou, China | GZNU20230215019 |
|
|
|
|
|
| 3 |
| Xinglong Town, Anlong County, Guzihou, China | GZNU20230215020 |
|
|
|
|
|
| 4 |
| – | – | – | – |
| – | – |
| 5 |
| – | CIFEFGB-Bb-02 |
|
| – | – | – |
| 6 |
| – | HSLBB | – |
| – | – | – |
| 7 |
| India, Karnataka, Ramnagar | BNHS FWF 193 |
|
| – | – | – |
| 8 |
| India, Karnataka, Kamra, Joida | WILD-15-PIS-230 |
|
| – | – | – |
| 9 |
| Menglun, Yunnan Province, China | IHB0301053 |
| – | – | – | – |
| 10 |
| Xishuangbanna, Yunnan Province, China | IHB0301030 |
| – | – | – | – |
| 11 |
| – | cyp74 | – | – |
|
| – |
| 12 |
| – | KUFOS-19-AN-BA-34.1 | – |
| – | – | – |
| 13 |
| Yuanjiang, Yunnan Province, China | IHB0805545 |
| – |
| – | – |
| 14 |
| – | GZNU20230215022 |
| – |
|
| – |
| 15 |
| Yuanjiang, Yunnan Province, China | IHB0805546 |
|
|
| – | – |
| 16 |
| Yuanjiang, Yunnan Province, China | IHB0805547 |
|
|
|
|
|
| 17 |
| / | / |
| – | – | – | – |
| 18 |
| India, Maharashtra, Venegaon | WILD-12-PIS-019 |
|
| – | – | – |
| 19 |
| India, Maharashtra, Venegaon | ZSI-WRC P/2849 |
|
| – | – | – |
| 20 |
| Jingxi City, Guangxi Province, China | GZNU20230215023 |
| – | – | – | – |
| 21 |
| Jingxi City, Guangxi Province, China | GZNU20230215024 |
| – | – | – | – |
| 22 |
| Jingxi City, Guangxi Province, China | SCAU-20190805001 |
|
| – | – | – |
| 23 |
| – | – | – | – |
| – | – |
| 24 |
| India, Karnataka, Hattihole | WILD-15-PIS-231 |
|
| – | – | – |
| 25 |
| India, Karnataka, Hattihole | BNHS FWF 197 |
|
| – | – | – |
| 26 | Dukang Town, Tiandeng County, Guangxi, China* | WGY2024121901 |
|
|
|
|
| |
| 27 | Dukang Town, Tiandeng County, Guangxi, China* | WGY2024121904 |
|
|
|
|
| |
| 28 |
| – | Vial 2006-0588 | – |
|
| – | – |
| 29 |
| – | Vial 2005-0948 | – |
|
| – | – |
| 30 |
| Panzhihua, Sichuan Province, China | IHB1004172 |
|
| – | – | – |
| 31 |
| Zhaotong, Yunna Province, China | IHB0709025 |
|
|
|
|
|
| 32 |
| – | KUFOS.2019.12.54 |
|
|
| – | – |
| 33 |
| Changting, Fujia Province, China | IHB0301051 |
|
| – | – | – |
| 34 |
| – | KUFOS-19-RR-GA-39.1 | – |
| – | – | – |
| 35 |
| – | KUFOS-19-AN-GA-40.1 | – |
| – | – | – |
| 36 |
| – | KUFOS-19-AN-GA-42.1 | – |
| – | – | – |
| 37 |
| – | cyp903 |
|
| – |
| – |
| 38 |
| Nujiang, Yunnan Province, China | / | – |
| – | – | – |
| 39 |
| Nujiang, Yunnan Province, China | ihb201305588 | – |
| – | – | – |
| 40 |
| Taiwan Province, China | – |
| – | – | – | – |
| 41 |
| Mudong, Chongqing City, China | IHB0809019 |
| – |
| – | – |
| 42 |
| – | – | – | – |
| – | – |
| 43 |
| – | CBM: ZF 11705 |
|
|
| – | – |
| 44 |
| – | CBM: ZF 12286 |
|
| – | – | – |
| 45 |
| – | – |
|
|
|
|
|
| 46 |
| – | NBFGR:8118E | – |
| – | – | – |
| 47 |
| – | CBM:ZF 12287 |
|
|
| – | – |
| 48 |
| – | – |
|
| – | – | – |
| 49 |
| – | – |
|
|
|
|
|
| 50 |
| – | – | – |
|
| – | – |
| 51 |
| – | CBM: ZF 12281 |
|
|
| – | – |
| 52 |
| – | Vial SN25 | – | – |
| – | – |
| 53 |
| Guizhou Province, China | IHB0709424 |
|
|
|
|
|
| 54 |
| Chongqing City, China | IHB0301068 |
|
|
| – | – |
| 55 |
| Jianou, Fujian Province, China | IHB0706070 |
|
|
| – | – |
| 56 |
| Jinkou, Hubei Province, China | IHB0803128 |
|
|
|
|
|
| 57 |
| Mudong, Chongqing City, China | IHB0301070 |
|
|
|
| – |
| 58 |
| Mudong, Chongqing City, China | IHB0301071 |
|
|
| – | – |
| 59 |
| Mudong, Chongqing City, China | IHB0809033 |
| – |
| – |
|
| 60 |
| Taoyuan, Hunan Province, China | IHB1004171/IHB030105 |
|
|
| – | – |
| 61 |
| Taiwan Province, China | ASIZP0806662 | – |
| – | – | – |
| 62 |
| – | – |
|
| – | – | – |
| 63 |
| Cishui, Guizhou Province, China | IHB0400184 |
|
|
|
|
|
| 64 |
| – | P4 |
| – |
|
|
|
| 65 |
| Neijiang, Sichuan Province, China | 20170920BB03 |
|
| – | – | – |
| 66 |
| Yuanjiang, Yunnan Province, China | IHB0805543 |
|
|
|
|
|
| 67 |
| Zhaoping, Guangxi Province, China | IHB0400321 |
|
| – | – | – |
| 68 |
| – | – | – |
| – | – | – |
| 69 |
| – | KUFOS-19-AN-TR-49.1 | – |
| – | – | – |
| 70 |
| Wuyishan, Fujian Province, China | IHB0706028 |
|
|
|
|
|
| Characters | ||||||||
|---|---|---|---|---|---|---|---|---|
| WGY2024121901 (holotype) | WGY2024121902 | WGY2024121903 | WGY2024121904 | WGY2024121905 | Mean ± SD | |||
| Dorsal-fin rays | iii,9 | iii,9 | iii,9 | iii,9 | iii,9 | - | iii, 8½ | iii,8 |
| Pectoral-fin rays | v,12 | v,12 | v,12 | v,12 | v,12 | - | viii, 11 | vii,12 |
| Pelvic-fin rays | ii,7 | ii,7 | ii,7 | ii,7 | ii,7 | - | ii, 9 | ii,7 |
| Anal-fin rays | i,6 | i,6 | i,6 | i,6 | i,6 | - | iii, 5½ | ii,5 |
| Caudal-fin rays | ii,19 | ii,19 | ii,19 | ii,19 | ii,19 | - | 17 | - |
| Lateral-line pores/scales | 69 | 72 | 70 | 69 | 75 | 71.5 ± 2.6 | 68 | 70 |
| Total length | 47.3 | 45.6 | 55.3 | 48.9 | 43.6 | 48.4 ± 5.1 | 60.4 | - |
| Standard length | 38.8 | 36.1 | 47.1 | 39.6 | 33.9 | 39.2 ± 5.8 | 47.9 | 63.2 |
| Body depth | 6.0 | 5.9 | 5.9 | 5.8 | 5.7 | 5.8 ± 0.1 | 5.2 | 12 |
| Body width | 6.5 | 6.3 | 6.1 | 6.2 | 6.0 | 6.2 ± 0.1 | 6.5 | 12.4 |
| Head length | 7.0 | 7.1 | 6.8 | 6.9 | 6.8 | 6.9 ± 0.1 | 10.3 | 12.6 |
| Head depth | 4.5 | 4.6 | 4.5 | 4.3 | 4.4 | 4.5 ± 0.1 | 4.9 | 8.4 |
| Head width | 6.9 | 7 | 7.1 | 6.8 | 7.0 | 7 ± 0.1 | 8.7 | 10.7 |
| Pre-anterior distance | 3.1 | 3.0 | 3.0 | 2.9 | 3.0 | 3 ± 0.1 | 3.5 | - |
| Distance between anterior nostrils | 2.5 | 2.3 | 2.4 | 2.5 | 2.4 | 2.4 ± 0.1 | 2.8 | - |
| Distance between posterior nostrils | 4.0 | 4.0 | 4.0 | 3.8 | 3.9 | 3.9 ± 0.1 | 3.8 | - |
| Distance between anterior and posterior nostrils | 2.2 | 2.0 | 2.1 | 2.1 | 2.0 | 2.1 ± 0.1 | 2 | - |
| Snout length | 3.0 | 2.9 | 2.8 | 3.0 | 2.8 | 2.9 ± 0.1 | 5.9 | 6.9 |
| Upper jaw length | 2.0 | 1.9 | 2.1 | 2.0 | 2.0 | 2 ± 0.1 | 2.8 | - |
| Lower jaw length | 1.1 | 1.0 | 0.9 | 1.0 | 1.0 | 1 ± 0.1 | 2 | - |
| Mouth width | 3.1 | 2.8 | 2.7 | 2.9 | 2.8 | 2.8 ± 0.1 | 3.6 | - |
| Eye diameter | 1.1 | 0.9 | 1.1 | 0.9 | 1.2 | 1 ± 0.2 | 1.3 | 1.9 |
| Interorbital distance | 4.1 | 3.8 | 3.9 | 4.2 | 4.1 | 4 ± 0.2 | 4.7 | 5.1 |
| Predorsal length | 16.4 | 17.3 | 16.2 | 17.1 | 15.8 | 16.6 ± 0.7 | 22.3 | 28.8 |
| Dorsal-fin base length | 6.2 | 5.8 | 5.6 | 6.1 | 5.9 | 5.9 ± 0.2 | 7.4 | - |
| Dorsal-fin length | 7.1 | 7.2 | 6.9 | 7.1 | 6.4 | 6.9 ± 0.4 | 10.9 | - |
| Pectoral-fin length | 8.1 | 7.8 | 8.3 | 7.5 | 7.2 | 7.7 ± 0.5 | 11.7 | 15.4 |
| Pectoral-fin base length | 4.4 | 3.5 | 3.2 | 4.1 | 3.2 | 3.5 ± 0.4 | 4.9 | - |
| Pre-pectoral length | 6.1 | 5.8 | 5.5 | 6.1 | 5.2 | 5.7 ± 0.4 | 9.3 | - |
| Pelvic-fin length | 6.2 | 5.8 | 5.9 | 5.8 | 5.8 | 5.8 ± 0.1 | 10 | - |
| Pelvic-fin base length | 2.8 | 2.5 | 2.6 | 2.7 | 2.6 | 2.6 ± 0.1 | 3.4 | - |
| Pre-pelvic length | 16.4 | 15.3 | 16.1 | 15.6 | 15.1 | 15.5 ± 0.4 | 22.6 | - |
| Anal-fin length | 6.4 | 5.8 | 5.9 | 6.2 | 5.9 | 6 ± 0.2 | 8.7 | - |
| Anal-fin base length | 3.6 | 3.4 | 3.5 | 3.5 | 3.5 | 3.5 ± 0.1 | 3.9 | - |
| Pre-anal length | 30.2 | 28.1 | 29.4 | 28.3 | 29.8 | 28.9 ± 0.8 | 34.8 | 47.3 |
| Distance between origin of pectoral fin and origin of ventral fin | 12.1 | 11.8 | 11.3 | 12.5 | 11.7 | 11.8 ± 0.5 | 10 | - |
| Distance between origin of ventral fin and origin of anal fin | 12.2 | 12.3 | 11.6 | 12.4 | 11.4 | 11.9 ± 0.5 | 10.6 | - |
| Distance between end of Anal fin and anus | 9.2 | 8.4 | 8.3 | 9.1 | 8.2 | 8.5 ± 0.4 | 1.8 | - |
| Caudal peduncle length | 8.5 | 8.1 | 8.2 | 8.2 | 8.1 | 8.2 ± 0.1 | 8.6 | - |
| Caudal peduncle depth | 8.5 | 9.2 | 8.3 | 9.1 | 8.6 | 8.8 ± 0.4 | 3.1 | - |
| Inner maxillary barbel length | 2.3 | 2.5 | 2.3 | 2.2 | 2.3 | 2.3 ± 0.1 | 1.4 | - |
| Outer maxillary barbel length | 2.1 | 2.2 | 2.1 | 2.1 | 2.1 | 2.1 ± 0.1 | 1.1 | 3.6 |
| Inner rostral barbel length | 1.2 | 1.1 | 1.3 | 1.2 | 1.1 | 1.2 ± 0.1 | 1.4 | - |
| Outer rostral barbel length | 2.3 | 2.2 | 2.1 | 2.2 | 2.1 | 2.2 ± 0.1 | 1.9 | - |
| ID | Species | Maxillary barbels | Dorsal fin rays | Anal fin rays | Pectoral fin rays | Pelvic fin rays | Lateral line scales | Dorsal black spot |
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | iii, 9 | i, 6 | v, 12 | ii, 7 | 69–75 | 6–7 | |
| 2 | 2 | iii, 8½ | iii, 5½ | viii, 11 | iii, 9 | 66–68 | 6–7 | |
| 3 |
| 1 | ?, 12 | ?, 8 | viii–x, 8 | - | 61–62 | - |
| 4 |
| 1 | iii, 8 | i, 5 | ix, 11 | ii, 9 | 65–69 | - |
| 5 | 1 | iii, 8 | i, 5 | ix, 11 | ii, 8 | 70–72 | - | |
| 6 |
| 1 | iii, 8 | iii, 5 | iii–ix, 11–12 | ii, 9 | 66–68 | 7 |
| 7 |
| 1 | iii, 9 | iii, 5–7 | vi, 10–12 | ii, 8–9 | 66–67 | Without |
| 8 |
| 1 | iii, 8 | ii, 5 | x, 10–12 | iii, 8 | 67 | 7 |
| 9 |
| 1 | iii, 8 | ii, 5 | ix, 10–11 | ii, 8–9 | 64–66 | 9 |
| 10 | 2 | iii, 8 | ii, 5 | vi–viii, 10–13 | ii, 8 | 61–65 | 6–8 | |
| 11 |
| 1 | iii, 8 | ii, 5 | viii, 10–12 | ii, 8–9 | 68–70 | 7–8 |
| 12 |
| 1 | iii, 8 | iii, 5 | viii–ix, 10–11 | ii, 8–9 | 66–68 | 10 |
| 13 | 2 | iii, 8 | ii, 5 | viii–x, 11–14 | ii, 9–11 | 74–76 | 8–9 | |
| 14 | 2 | iii, 8 | ii, 5 | vi–vii, 11–12 | ii, 6–7 | 69–74 | 6–9 | |
| 15 |
| 1 | ?, 11 | ?, 8 | ?, 9–10 | - | - | - |
| 16 |
| 1 | iii, 8–9 | ii, 5 | viii-ix 10–12 | ii, 8–9 | 68–69 | - |
| 17 | 1 | iii, 8 | ii, 5 | viii–x, 9–12 | ii, 9 | 59–64 | - | |
| 18 |
| 1 | iii, 8 | ii, 5 | x, 10–12 | iii, 8 | 67 | 7 |
| 19 |
| 1 | iii, 7 | ii, 5 | xii, 13 | v, 13 | 74 | 8 |
| 20 | 1 | iii, 7 | iii, 5½ | ix, 14 | ii, 9 | 63–67 | - |
|
|
|
|
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|---|---|---|---|
|
| ||||||||||
|
| 0.95 | |||||||||
|
| 0.58 | 0.33 | ||||||||
|
| 1.89 | 0.77 | 0.52 | |||||||
|
| 1.26 | 0.58 | 0.01 | 1.1 | ||||||
|
| 0.58 | 0.3 | 0.55 | 0.06 | 0 | |||||
|
| 1.39 | 0.7 | - | 1.35 | 1.2 | - | ||||
|
| 1.88 | 0.8 | 0.49 | 0.56 | 1.11 | 0.11 | 1.28 | |||
|
| 1.98 | 0.84 | 0.64 | 1.86 | 1.11 | 0.67 | 1.1 | 1.88 | ||
|
| 1.78 | 0.83 | 0.4 | 1.67 | 1.12 | 0.49 | 1.26 | 1.54 | 1.72 | |
|
| 2.51 | 0.89 | 0.63 | 2.23 | 1.57 | 0.6 | 1.62 | 2.25 | 1.18 | 2.24 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies · Ichthyology and Marine Biology · Fish biology, ecology, and behavior
Introduction
The genus Balitora, Gray, 1830 was established with Balitora brucei Gray, 1930 as the type species, originally placed in Cobitidae but it is currently the type genus for the family Balitoridae (Kottelat and Chu 1988; Fricke et al. 2025). The genus Balitora has long been the subject of taxonomic controversy, with different taxonomic schemes proposed based on morphological differences (Zheng et al. 1982; Zheng and Zhang 1983; Li and Chen 1985). At first, it was recognized by one pair of maxillary barbels, a character that distinguishes Balitora from other genera (Chen 1978). Kottelat (1988) and Kottelat and Chu (1988) reviewed this genus, revealed the unreliability of maxillary-barbel number in defining the three Balitorid genera, Balitora, Hemimyzon, and Sinohomaloptera, and proposed that Sinohomaloptera be a junior subjective synonym of Balitora. Based on three or more unbranched pelvic fin rays, B. elongata Chen & Li, 1985, B. nujiangensis Zhang & Zheng, 1983, B. pengi Huang, 1982, and B. tchangi Zheng, 1982 were placed in Hemimyzon Regan 1911, while species with two branched pelvic fin rays were placed in Balitora, namely B. kwangsiensis (Fang, 1930), B. lancangjiangensis (Zheng, 1980), and B. longibarbata (Chen, 1982) (Kottelat and Chu 1988). However, Chinese scholars did not embrace the proposal by Kottelat and Chu (1988), and continued to classify species within the genus Balitora based on the count of maxillary barbels as having one or two pairs of maxillary barbels at each corner of mouth. So far, no in-depth phylogenetic studies of Balitoridae species have been conducted.
Nevertheless, previous morphology-based studies have not resolved the phylogenetic relationships between Balitora, Hemimyzon, and Sinohomaloptera, with various studies concluding that the phylogenetic position of the genus Balitora is unclear and may not be monophyletic (Šlechtová et al. 2007; Liu et al. 2012a; Kumkar et al. 2016; Tao et al. 2019; Shao et al. 2020). The genus Balitora includes both surface-dwelling and cave-adapted species (Liu et al. 2012b; Luo et al. 2023). To date, the classification of the genus remains controversial, although Luo et al. (2023) showed clearly in their phylogenetic analysis that Balitora is not a monophyletic and contains three distinct clades with significant genetic divergence among species, but also suggested that more evidence was needed to further clarify the taxonomic composition of the genus Balitora.
In this study, we followed the latest taxonomic scheme for the genus Balitora (Balitoridae, Cypriniformes) as proposed by Luo et al. (2023), which integrated molecular phylogenetic analyses based on two mitochondrial (COI and Cyt b) and three nuclear genes (RAG1, IRBP, and EGR2B) with geometric morphometrics to resolve longstanding taxonomic controversies. According to the Catalog of Fishes (Fricke et al. 2025), the genus currently comprises 22 recognized species, of which twelve species are distributed in China, namely B. anlongensis Luo, Chen, Zhao, Yu, Lan & Zhou, 2023, B. brucei, B. dehouensis Lei, Pu & Yang, 2025, B. elongata, B. kwangsiensis, B. lancangjiangensis, B. longibarbata, B. ludongensis Liu & Chen, 2012, B. nantingensis Chen, Cui & Yang, 2005, B. scyphus Endruweit, 2025, B. tchangi, and Balitora yingjiangensis (Chen, 2006), which was previously mistakenly classified as Hemimyzon, and was re-described through examination of type specimens and collection of topotypic specimens from the Jieyanghe River, a tributary of the Irrawaddy River in Nabang Town, Yingjiang County, Yunnan Province, China (Liu et al. 2012b; Luo et al. 2023; Endruweit 2025; Lei et al. 2025a, 2025b).
During our field surveys for plant resources in Tiandeng County, Guangxi in December 2024, we unexpectedly collected five specimens of the genus Balitora in a karst cave. After consulting relevant literature (Kottelat and Chu 1988; Kottelat 1988; Chen et al. 2005; Nguyen 2005; Conway and Mayden 2010; Liu et al. 2012b; Luo et al. 2023; Lei et al. 2025a, 2025b) and comparing with relevant specimens, we confirm that this unusual fish represents a new species, which is described below. In addition, all five specimens were collected from the same karst cave water system, and this species has not been found in open surface waters outside the cave. Their eye diameter is small and light-sensitive, so we define it as a new species of cave-dwelling loach according to the study of Culver and Pipan (2019).
Material and methods
Sampling and morphological measurement
Five specimens representing a new species, were collected from Dukang Town, Tiandeng County, Guangxi Zhuang Autonomous Region, China for morphological comparison and two of them were sampled for genetic analysis (vouchers WGY2024121901 and WGY2024121904). All specimens were then fixed in 10% buffered formalin and later transferred to 70% ethanol for preservation. Muscle samples used for molecular analysis were preserved in 95% alcohol and stored at −20 °C. All specimens are deposited at Guangxi Institute of Chinese Medicine & Pharmaceutical Science (GXMI), Nanning City, Guangxi Zhuang Autonomous Region, China under the holotype number WGY2024121901 and paratypes numbers, WGY2024121902–WGY2024121905.
The new species was described, based on field observations that were made in December 2024 and examination of specimens at GXMI. Other related Balitora species examined during this study are based on data from the National Specimen Information Infrastructure (http://nsii.org.cn/2017/), FishBase (https://fishbase.mnhn.fr/), and National Animal Specimen Resource Center (http://museum.ioz.ac.cn/index.html). The morphological data of 19 species of the genus Balitora comes from relevant literature (Liu et al. 2012b; Luo et al. 2023; Lei et al. 2025a, 2025b). All measurements of the new species were taken on the left side of the fish specimens, point to point with digital calipers at 0.1 mm accuracy. Counts and proportional measurements follow Tang et al. (2012).
Molecular phylogenetic analysis
The genomic DNA was extracted from muscle tissues by standard phenol chloroform methods (Sambrook and Russell 2001). Then two tissue samples used for molecular analysis were amplified and sequenced for two mitochondrial genes and three protein-coding genes: mitochondrial gene cytochrome b (Cyt b), cytochrome oxidase subunit 1 (COI), recombination activating gene 1 (RAG1), retinoid binding protein (IRBP), and early growth response protein 2B (EGR2B) as in Luo et al. (2023). Primer sequences are shown in Suppl. material 1. Both the amplification and sequencing were completed in Beijing Ruijie Gene Technology Co., Ltd (Beijing, China). All sequences were aligned using MAFFT v. 7.490 (Katoh and Standley 2013). The sequences after alignment were connected in series using the “Concatenate” tool in Genious Prime v. 2021.1.1 (Kearse et al. 2012). The aligned matrices were automatically trimmed using trimAL v. 1.4 (Capella-Gutiérrez et al. 2009) to remove non-conservative sequence parts. Maximum likelihood phylogenies were inferred using IQ-TREE v. 2.2.0 (Nguyen et al. 2014) under the GTR+I+G4+F model for 1000 ultrafast bootstraps (Minh et al. 2013). Vouchers of specimens and GenBank accession numbers for this study are presented in Table 1.
Results
Molecular phylogenetic analysis
The ML tree based on two mitochondrial genes and three nuclear genes (Fig. 1) shows that Balitora is not a monophyletic and contains three distant clades. Clade I, including, B. anlongensis, B. ludongensis, and B. tiandengensis; Clade II, including, B. annamitica Kottelat, 1988, B. elongata Chen & Li, 1985, B. kwangsiensis, and B. meridionalis Kottelat, 1988; and Clade III, including B. brucei Gray, 1830, B. chipkali Kumar, Katwate, Raghavan & Dahanukar, 2016, B. elongata Chen & Li, 1985, B. jalpalli Raghavan, Tharian, Ali, Jadhav & Dahanukar, 2013, B. laticauda Bhoite, Jadhav & Dahanukar, 2012, and B. mysorensis Hora, 1941. Balitora tiandengensis sp. nov. formed sister groups with B. ludongensis. The smallest and biggest uncorrected p-distance (%) between Balitora tiandengensis sp. nov. and other species of the genus Balitora were 0.6% with B. jalpalli and 2.51% with B. anlongensis (Table 4). It can be distinguished by the combination of the following morphological characters: dorsal fin rays iii, 9, anal fin rays i, 6, pectoral fin rays v, 12, pelvic fin rays ii, 7, lateral line scales 69–75. The combination of distinct morphological characteristics and phylogenetic evidence supports the recognition of this previously unidentified population as a novel taxonomic entity. Thus, the population at this locality represents an independently evolved lineage and is described as a new species, Balitora tiandengensis sp. nov.
Maximum likelihood phylogenetic reconstruction of Balitora tiandengensis sp. nov. and its related species based on combined dataset COI+Cyt b+RAG1+IRBP+EGR2B. The bootstrap supports from maximum likelihood (ML) analyses are indicated alongside the corresponding nodes.
Taxonomy account
Balitora
tiandengensis
Taxon classificationAnimaliaCypriniformesBalitoridae
Nong & Wei sp. nov.
0651CEAB-4DE3-5001-9EA6-B5618F8BBF7A
https://zoobank.org/A088AD1B-5C8B-476F-8E8C-D38D5BDB6AEE
Type material.
Holotype • WGY2024121901 (Fig. 2), 47.3 mm total length (TL), 38.8 mm standard length (SL), collected by You Nong & Gui-Yuan Wei on 19 December 2024 in a karst cave nearby Duoru Village, Dukang Town, Tiandeng County, Guangxi Zhuang Autonomous Region, China (23°03'45.32"N, 106°59'44.81"E, 545 m; Fig. 5). Paratypes • Four specimens, WGY2024121902–WGY2024121905, 43.6–55.3 mm TL, 33.9–47.1 mm SL, collected together with the holotype.
Morphological characters of holotype (WGY2024121901) of Balitora tiandengensis sp. nov. A. Lateral view; B. Dorsal view; C. Ventral view; D. Ventral side view of head; E. Dorsal side view of head; F. Close up photo of the mouth.
Oral morphology of Balitora tiandengensis sp. nov. A. Holotype (WGY2024121901); B. Paratype (WGY2024121902); C. Paratype (WGY2024121904).
Habitat and photographs of Balitora tiandengensis sp. nov. in life.
The distribution of similar species of Balitora, B. tiandengensis sp. nov. (red square), B. ludongensis (blue square), and B. anlongensis (black square).
Diagnosis.
Balitora tiandengensis sp. nov. can be distinguished from other congeners by the following combination of characters: (1) two pairs of maxillary barbels; (2) dorsal fin rays iii, 9; (3) pectoral fin rays v, 12; (4) pelvic fin rays ii, 7; (5) anal fin rays i, 6; (6) lateral-line scales 69–75; and (7) 6–7 indistinctly separated transversely oval blotches on the dorsal side.
Description.
Morphological data of the five specimens of the Balitora tiandengensis sp. nov. are provided in Table 2. Body elongated and sub-cylindrical, posterior portion gradually compressed from dorsal fin to caudal-fin base, with deepest body depth anterior to dorsal-fin origin, deepest body depth 15.4–16.3% of SL. Dorsal profile slightly convex from snout to dorsal-fin insertion, then straight from posterior portion of dorsal-fin origin to caudal-fin base. Pelvic profile flat. Head blunt and depressed, head length (HL) 18.7–19.4% of SL and greater than head width, head width greater than depth (head width/head depth = 1.4). Snout short, oblique, and blunt, length 40.9–43.5% HL. Interorbital space wide and flat. Mouth inferior, small and curved, mouth corner situated below anterior nostril, upper and lower lips smooth and fleshy. Relatively shallow preoral groove present between rostral cap and upper lip, extending across corners of mouth. Mouth width 40.0–43.5% of head width. Rostral cap around upper lip divided into three lobes, median one largest, slightly curved. Four pairs of barbels: two pairs of rostral barbels, short, outer rostral barbel longer than inner one; two pairs of maxillary barbels, short, situated at corner of mouth: outer maxillary barbels longer than inner ones. Mouth inferior, small and curved, mouth corner situated below anterior nostril, lips without complex folding or thickening, maintaining a relatively thin and simple structure, upper and lower lips smooth and fleshy. Upper and lower lips connected at corner of mouth, and upper lip with a row of inconspicuous papillae, and lower lip thin. Lower jaw with radiate ridges on its surface. Two longitudinal fleshy ridges on mid-chin. Anterior and posterior nostrils closely set, anterior nostril without any elongated barbel-like tip. Eyes small, 12.7–17.1% HL. Gill opening small, gill rakers not developed.
Table 2.: Morphological characters and measurement data (mm) of the new species Balitora tiandengensis sp. nov. and B. anlongensis (data from Luo et al. 2023) and B. ludongensis (data from Liu et al. 2012b).
Dorsal fin iii, 9; anal fin i, 6; pectoral fin v, 12; pelvic fin ii, 7; and 19 branched caudal-fin rays. Dorsal fin long, 16.7–19.4% of SL, nearly equal to head length, distal margin truncated, origin anterior to pelvic fin insertion, situated slightly anterior to midpoint between snout tip and the caudal-fin base, first branched ray longest, shorter than HL, tip of the dorsal fin extending to the vertical of the anus. Pectoral fin elongated and developed, distal margin rounded, pectoral fin length approximately equal than HL, 18.4–20.5% of SL, tip of the pectoral fin extends backward beyond 54.0%–60.0% of the distance between the origin of the pectoral fin and the origin of the pelvic fin, without reaching to the pelvic fin-origin. Pelvic fin moderately developed, distal margin rounded, pelvic fin length slightly shorter than HL, 15.1–16.7% of SL, vertically aligned with the fourth branched ray of the dorsal fin, pelvic fin origin closer to the snout tip than the caudal-fin base and closer to the anal fin origin than the snout tip, tips of the pelvic fin reaching to the anus. Anus 72.5%-73.3% distance from posterior end of the pelvic fin base to the anal fin origin. Anal fin short, 15.0–16.7% of SL, distal margin truncated, origin close to the anus and far from the caudal-fin base, spacing ~ 2.5 mm, tips of the anal fin extending backwards and not reaching caudal-fin base, distance between the end of the anal fin and the anus 7.5–9 × the eye diameter. Caudal fin deeply forked, upper lobe equal in length to the lower one, tips pointed, caudal peduncle length 8.1 mm, caudal peduncle depth 2.8 mm, without adipose crests along both dorsal and ventral sides.
Body smooth, covered with thin scales all over except for on the ventral side, head, and fins. Lateral line complete and straight, with 69–75 lateral line scales, exceeding the tip of the pectoral fin and reaching the base of the caudal fin.
Sexual dimorphism.
No sexual dimorphism was observed based on the present specimens of Balitora tiandengensis sp. nov.
Coloration in life.
Dorsal and lateral side of body brown, while ventral side white with slightly pinkish. Dorsal side of head blackish-brown, usually with black irregular blotches. Dorsal side of body with six or seven transverse black round blotches encircled by yellow interspace, usually two or three blotches in front of dorsal-fin origin, one or two on dorsal-fin base and three or four behind. Dorsal fin almost hyaline with discontinuous black spots. Pectoral and pelvic fin bases pale gray from dorsal view; dorsal side of pectoral and pelvic fin rays blackish spots, membrane lighter; distal tip of pectoral and pelvic fin hyaline and colorless. Anal fin hyaline with a pale blackish bar in the middle. Caudal fin almost hyaline with blackish spots. Lateral line of the body from the posterior to the eye to the base of the caudal fin pale grayish-yellow. After being moved from inside the cave to outside, the body pigmentation lightened in ~ 6 hours while it was alive.
Coloration in alcohol.
After being fixed in 10% formalin and stored in 70% ethanol, the body pigmentation deepened. Other parts of the body similar to the living condition.
Distribution.
Balitora tiandengensis sp. nov. is only known from the type locality, a karst cave near Duoru Village, Dukang Town, Tiandeng County, Guangxi, China at an elevation of 545 m (Fig. 5).
Ecology.
Within this cave, Balitora tiandengensis sp. nov. co-occurred with catfish (Silurus sp). Around the cave, the arable land was farmed to produce tea (Ilex latifolia Thunb.).
Etymology.
The specific epithet tiandengensis is in reference to the type locality of the new species: Duoru Village, Dukang Town, Tiandeng County, Guangxi Zhuang Autonomous Region, China. We propose the common English name “Tiandeng cave loach” and the Chinese name “tiān děng pá qiū (天等爬鳅)”.
Discussion
The new species is assigned to the genus Balitora based on the combination of the following diagnostic characters (Kottelat 1988; Kottelat and Chu 1988): (1) body strongly depressed; head and abdomen ventrally flattened; (2) mouth inferior, arched, with both jaws covered by a horny sheath; (3) rostral flap divided into three lobes, the lips exhibit no complex folding or thickening, maintaining a relatively thin and simple structure; upper lip with a row of inconspicuous papillae; (4) two pairs of maxillary barbels; (5) gill-openings extending on the ventral surface of head; (6) two unbranched pelvic rays, eight branched pelvic rays; (7) five unbranched pectoral rays, ten branched pectoral rays; and (8) adhesive pads present on ventral surface of the 8–11 anterior most pectoral rays and three or four anteriormost pelvic rays.
Our molecular phylogenetic analysis, based on COI+Cyt b+RAG1+IRBP+EGR2B genes, confirms Balitora tiandengensis as a distinct lineage within Balitora, supported by high bootstrap values (100%) and genetic divergence metrics (uncorrected p-distance ≥ 0.6% from congeners). This aligns with prior studies demonstrating that Balitora is non-monophyletic (Luo et al. 2023; Lei et al. 2025a), with species clustering into three clades: one containing the type species B. brucei, another comprising B. kwangsiensis (formerly Sinohomaloptera), and a third including B. tiandengensis, B. anlongensis, and B. ludongensis.
Morphologically, Balitora tiandengensis is distinguished from congeners by its unique combination of traits: (1) two pairs of maxillary barbels; (2) dorsal fin rays iii, 9; (3) pectoral fin rays v, 12; (4) pelvic fin rays ii, 7; (5) anal fin rays i, 6; (6) lateral-line scales 69–75; and (7) six or seven indistinctly separated transversely oval blotches on the dorsal side. These features contrast with B. ludongensis, with which it shares a similar distribution but differs in having more dorsal fin rays, anal fin rays, pelvic fin rays, and lateral line scales (Liu et al. 2012b). Such morphological plasticity within the genus underscores the need for integrative taxonomic approaches, as reliance on single traits (e.g., barbel number) has historically led to misclassifications (e.g., B. pengi and B. tchangi were initially placed in Hemimyzon due to overlapping morphological characters; Kottelat 1988).
Balitora tiandengensis sp. nov. differs from B. annamiticaKottelat 1988, B. brucei, B. burmanica, B. chipkali, B. eddsi Conway & Mayden, 2010, B. jalpalli, B. lancangjiangensis, B. laticauda, B. meridionalis, B. mysorensis, B. nantingensis, B. elongata, B. tchangi, and B. yingjiangensis based on the presence of two maxillary barbels at each corner of the mouth (vs 1). Balitora tiandengensis sp. nov. differs from B. kwangsiensis, B. longibarbata, and B. ludongensis based on the dorsal fin rays (iii, 9 vs iii, 8). Balitora tiandengensis sp. nov. can be further distinguished from B. brucei, B. burmanica, B. chipkali, B. jalpalli, B. kwangsiensis, B. lancangjiangensis, B. laticauda, B. longibarbata, B. ludongensis, B. meridionalis, B. nantingensis, and B. elongata based on dorsal fin rays (iii, 9 vs iii, 8). Balitora tiandengensis sp. nov. can be further distinguished from B. brucei, B. burmanica, B. chipkali, B. jalpalli, B. kwangsiensis, B. lancangjiangensis, B. laticauda, B. longibarbata, B. ludongensis, B. meridionalis, B. nantingensis, and B. elongata based on dorsal fin rays (iii, 9 vs iii, 8) (Table 3).
Balitora tiandengensis sp. nov. resembles B. ludongensis but can be further distinguished based on the dorsal fin rays (iii, 9 vs iii, 8), anal fin rays (i, 6 vs ii, 5), pectoral fin rays (v, 12 vs vi–vii, 11–12), lateral line scales (69–75 vs 66–68). Its percentage of body depth/SL (15.5 vs 18.9), body width/SL (16.8 vs 19.6), head length/SL (18.0 vs 20.0), predorsal length/SL (42.3 vs 45.6), preanal length/SL (77.8 vs 74.9), head depth/head length (64.3 vs 66.3), head width/head length (98.6.0 vs 85.1), eye diameter/head length (15.7 vs 14.8) (Table 3).
Balitora tiandengensis sp. nov. is similar to B. anlongensis but it can be distinguished from the latter by the combination of the following morphological characters: dorsal fin rays (iii, 9 vs iii, 8), anal fin rays (i, 6 vs iii, 5), pectoral fin rays (v, 12 vs viii, 11), pelvic fin rays (ii, 7 vs iii, 9), lateral line scales (69–75 vs 66–68). Its standard length (33.9–47.1 mm vs 39.8–48.2 mm), head length (6.8–7.1 mm vs 8.4–10.6 mm), snout length (2.8–3.0 mm vs 4.9–6.1 mm), outer maxillary barbel length (2.1–2.2 mm vs 0.9–1.5 mm) (Table 3).
The discovery of Balitora tiandengensis sp. nov. significantly advances our understanding of the systematics and biogeography of the genus Balitora, a group of rheophilic fishes adapted to high-gradient stream environments across south and southeast Asia (Kottelat and Chu 1988; Kottelat 1988; Nguyen 2005; Conway and Mayden 2010; Liu et al. 2012b; Luo et al. 2023; Lei et al. 2025a, 2025b).
The distribution of B. tiandengensis extends the known range of Balitora eastwards. This finding aligns with recent discoveries of cave-adapted Balitora in Guizhou (Luo et al. 2023), suggesting that the genus has undergone multiple dispersal events across mountain ranges.
Balitora tiandengensis inhabits in cave with fast-flowing, oxygen-rich waters, where it likely plays a role in controlling algal biofilms and invertebrate populations. However, similar to other Balitora species, B. tiandengensis may exhibit low dispersal capacity due to its specialized morphology, increasing its susceptibility to local extinctions. Future research should assess population genetics and habitat requirements to inform management strategies.
Supplementary Material
XML Treatment for Balitora tiandengensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics (Oxford, England) 25(15): 1972–1973. 10.1093/bioinformatics/btp 348PMC 271234419505945 · doi ↗ · pubmed ↗
- 2Chen YY (1978) Systematic studies on the fishes of the family Homalopteridae of China I. classification of the fishes of the subfamily Homalopterinae. Shui Sheng Sheng Wu Hsueh Bao 6(3): 331–348. 10.3724/issn 1000-3207-1976-3-331-o [In Chinese] · doi ↗
- 3Chen XY, Cui GH, Yang JX (2005) Balitora nantingensis (Teleostei: Balitoridae), a new hillstream loach from Salween drainage in Yunnan, southwestern China. The Raffles Bulletin of Zoology (Suppl. no. 13): 21–26.
- 4Conway KW, Mayden RL (2010) Balitora eddsi, a new species of hillstream loach (Ostariophysi: Balitoridae) from Nepal. Journal of Fish Biology 76(6): 1466–1473. 10.1111/j.1095-8649.2010.02575.x 20537025 · doi ↗ · pubmed ↗
- 5Culver DC, Pipan T (2019) The Biology of Caves and Other Subterranean Habitats, 2nd ed., Oxford, Academic Publishers, Oxford. 10.1093/oso/9780198820765.001.0001 [accessed 8 July 2025] · doi ↗
- 6Endruweit M (2025) Fishes of the Red River. Cypriniformes, Siluriformes. Isernhagen, Germany, 295 pp.
- 7Fricke R, Eschmeyer WN, Van der Laan R [Eds] (2025) Eschmeyer’s catalog of fishes: genera, species, references. https://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp [accessed 20 December 2025]
- 8Gray JE (1830) Illustrations of Indian Zoology: Chiefly Selected from the Collection of Major-General Hardwicke (Vol. 1). Treuttel, Wurtz, Treuttel, Jun. and Richter, London, 200 pp. 10.5962/bhl.title.95127 · doi ↗