A new lineage of looper moths discovered in the South African Cape Floristic Region (Lepidoptera, Geometridae)

Abstract
Click any figure to enlarge with its caption.
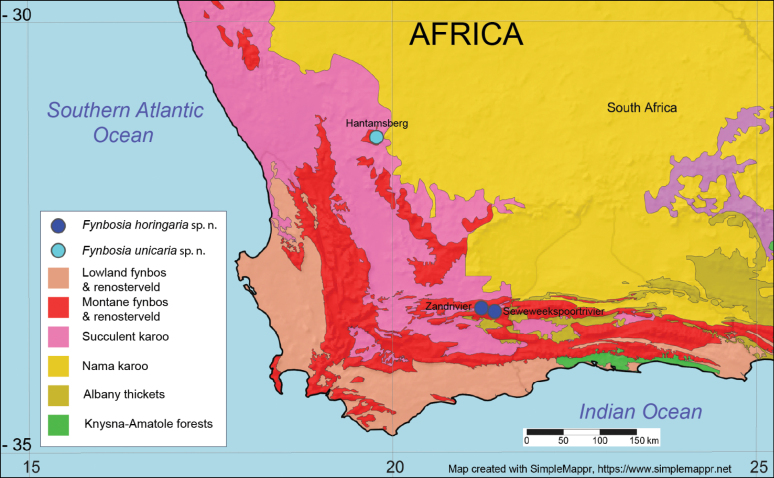 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3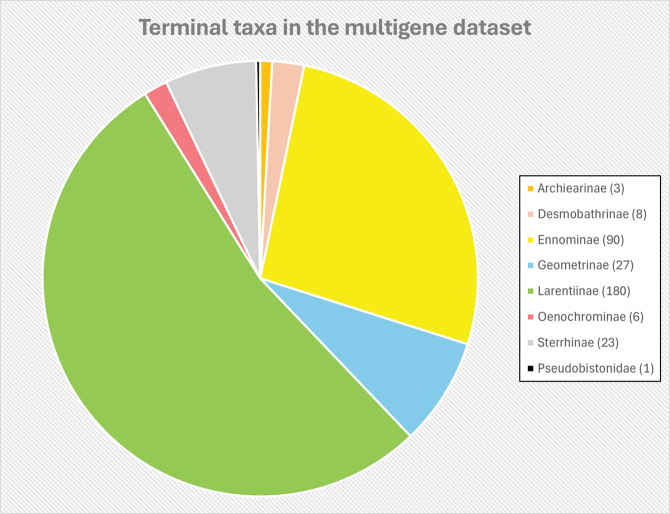 Figure 4
Figure 4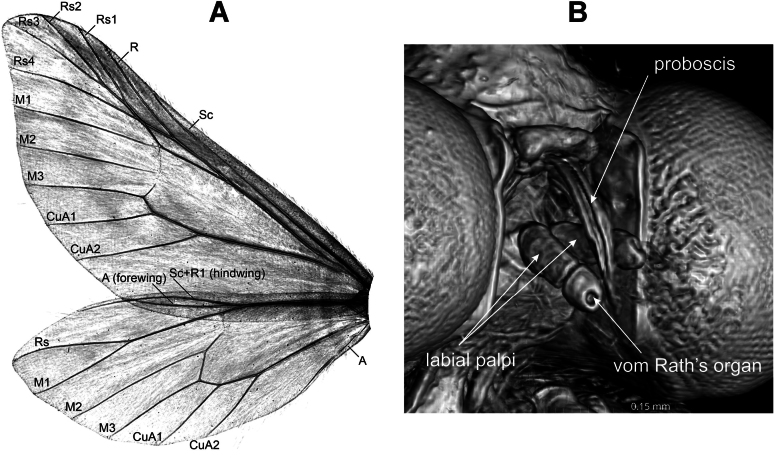 Figure 5
Figure 5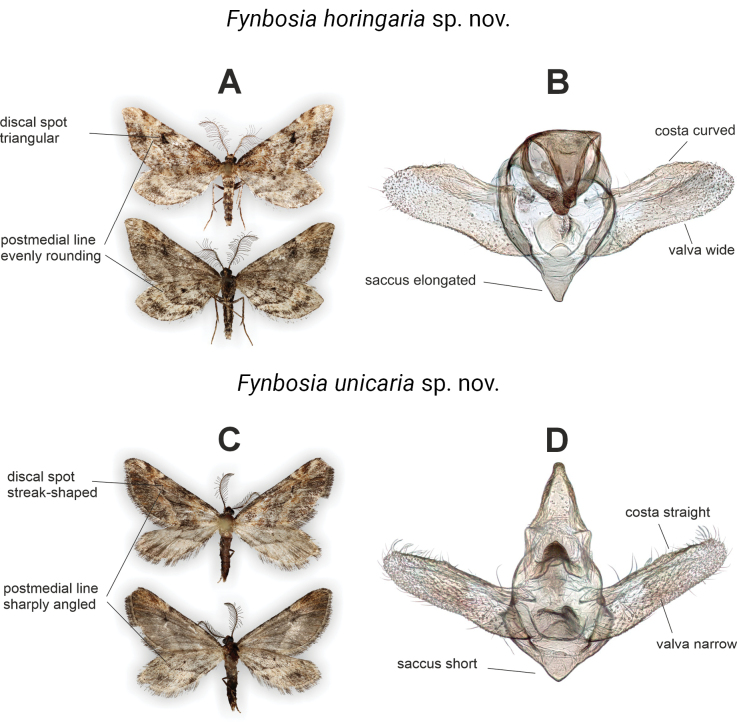 Figure 6
Figure 6 Figure 7
Figure 7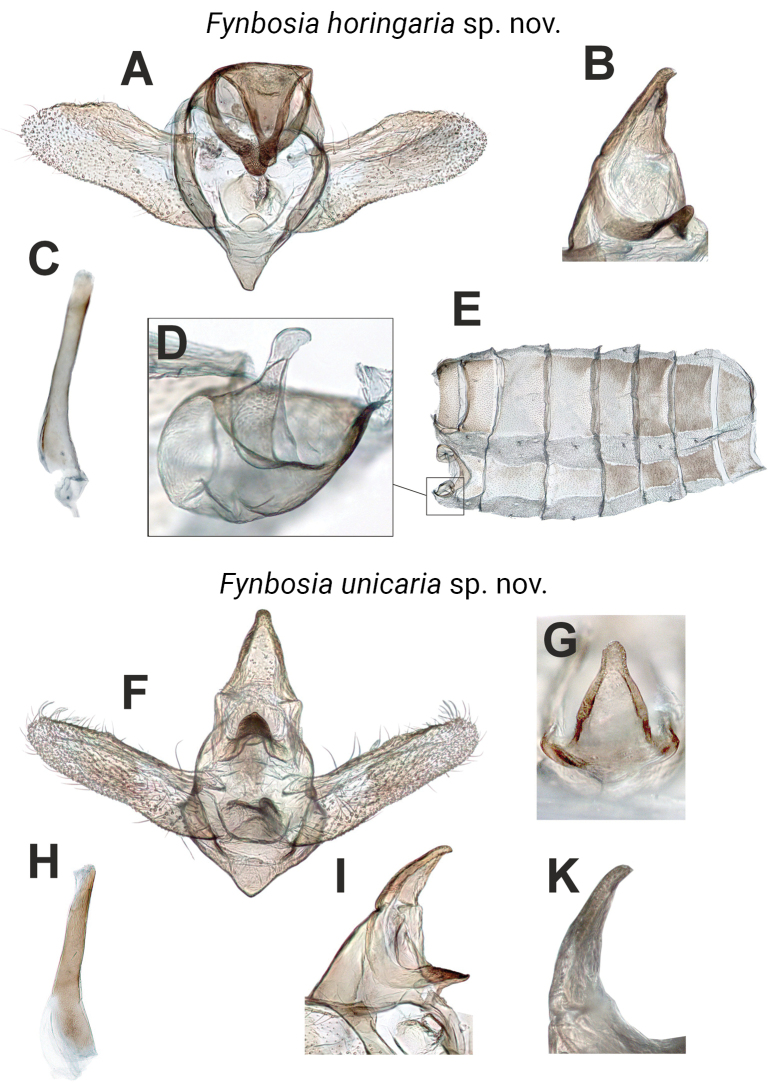 Figure 8
Figure 8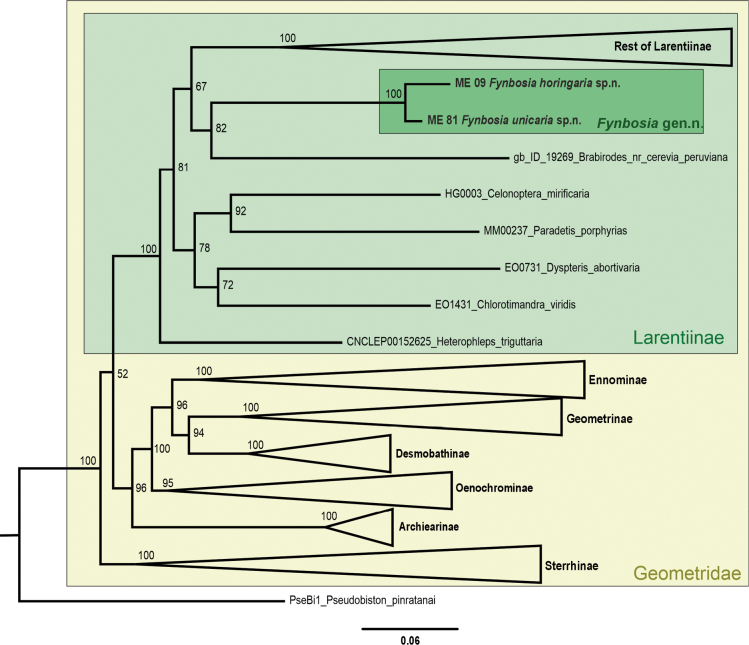 Figure 9
Figure 9| 1 | Postmedial lines evenly rounded, best visible on the wing underside; forewing discal spot triangular (Fig. | |
| – | Postmedial line sharply angled, forewing discal spot streak-shaped (Fig. |
| 1 | Valva wide, length of valva less than three times the height of the valva, costa curved, saccus elongated (Fig. | |
| – | Valva narrow, length of valva more than three times height of valva, costa straight, saccus short (Fig. |
- —Academy of Finland 501100002341 https://ror.org/05k73zm37 http://doi.org/10.13039/501100002341
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLepidoptera: Biology and Taxonomy · Biological Control of Invasive Species · Insect Pheromone Research and Control
Introduction
The Cape Floristic Region (CFR, Fig. 1) in the southwestern tip of Africa is recognized as one of the six floral kingdoms of the Earth with a high biodiversity and highly endemic flora and fauna (Born et al. 2007; Grobler and Cowling 2021; Samways et al. 2024a). Almost 11 500 species of plants exist in the CFR, of which more than three-quarters are endemic (Forest et al. 2018). The CFR can be divided into several biomes, each subdivided into several subtypes (e.g., Mucina and Rutherford 2006, Fig. 1). Due to the high floral endemism and stable geological history, the region also harbours a high but incompletely studied phytophagous insect diversity (Procheş and Cowling 2006; Kemp and Ellis 2017; Samways et al. 2024a). Several undescribed species of geometrid moths from the CFR are present in extant insect collections, and we discovered two unknown male moth specimens in the Western and Northern Cape provinces, some 300 kilometres apart, in March and April of 2022. These specimens turned out to belong to two closely related undescribed species representing an isolated lineage that we classify in the moth family Geometridae and subfamily Larentiinae.
The Cape Floral Region and the known locations of Fynbosia moths. Map base and vegetation type limits retrieved from https://www.simplemappr.net.
We used an integrative approach (Padial et al. 2010) for species delimitation. i.e., the validity of a species is more probable if supported by several unlinked character sets widely used in geometrid taxonomy (see e.g., Hausmann 2001; Sihvonen 2005; Müller et al. 2019) such as external and genitalia morphology, mitochondrial DNA barcode and distribution. Here, we present an integrative taxonomy of the new taxa and describe the genus Fynbosia (gen. nov.) and two species F. horingaria (sp. nov.) and F. unicaria (sp. nov.) as new to science.
Materials and methods
During an expedition to the South African Swartberg Mountain Range in late April 2022, we found a small, rather worn geometrid moth individual attracted to the dim light of our bathroom window in the farm Zandrivier near Calitzdorp (Figs 1, 2A, 3A, C). Since none of the authors (ME, HS, PS) could recognize the specimen on site, we took it with us for further examination to Helsinki, Finland.
Habitats of Fynbosia moths: Fynbosia horingaria sp. nov., habitats from Zandrivier (A) and Seweweekspoortrivier (B) near Calitzdorp, South Africa; habitat of Fynbosia unicaria sp. nov., on Hantamsberg near Calvinia, South Africa (C); live Fynbosia male specimen (D). Photo (B) retrieved from https://www.facebook.com/7weekspoort/.
Adult male specimens of Fynbosia. Fynbosia horingaria sp. nov., holotype (A, C, E, F): A. Dorsal view; C. Ventral view; E, F. Paratypes dorsal view. Fynbosia unicaria sp. nov., holotype (B, D): B. Dorsal view; D. Ventral view.
Later, Hermann Staude recognised that he had collected a rather similar appearing geometrid male moth from the Hantamsberg near Calvinia, Northern Cape, South Africa (Figs 1, 2C, 3B, D) some four weeks earlier. This specimen turned out to belong to a related, but different species.
In mid-April of 2023, Axel Hofmann and Jörg-Uwe Meinecke collected three more specimens, two males and a female of the first-mentioned species from Staude’s light trap set up in Seweweekspoortrivier, near Calitzdorp (Figs 1, 2B), some four kilometres southeast of the Zandrivier farm, where the first specimen was found. Unfortunately, the female specimen has not been located for closer scrutiny. We discovered a further two males (Fig. 3E, F) in 2025 from material collected in 2022 at Zandrivier Farm by the authors.
Authors (HS, ME, PS), Jannik Wagner (JW) of the State Museum of Natural History Stuttgart (SMNS), and Axel Hausmann (AH) of the Bavarian State Collection of Zoology in Munich (ZSM), examined the Lepidoptera collections of Ditsong Natural History Museum, Pretoria, South Africa (DMNH; HS, ME, JW), Natural History Museum in London (NHMUK; PS), and ZSM (AH, PS) for additional specimens similar to those we had discovered but found none. Consequently, we are aware of just seven specimens belonging to this undescribed moth lineage, and we had four male specimens (Fig. 3) from 2022 available for closer examination and description. All specimens were collected under the relevant permits held by Hermann Staude.
All specimens were attracted to artificial light at night: the F. horingaria sp. nov. holotype to a standard room light and the rest to battery-operated UV bulbs placed inside a custom-made light trap (Fig. 2C).
The holotype specimens were photographed using a Canon digital camera (model: EOS 5D) with an EF 100 mm macro lens attached to a StackShot automated macro rail, and the resulting multifocal images stacked in Zerene Stacker software (v. 1.04) (Figs 3, 6). The stacked image files were edited in Adobe Photoshop (v. 25.4.0), and the final plates were arranged using CorelDraw (v. 25.1.0) and Adobe Illustrator (v. 28.1).
The type specimens were scanned using a Nikon micro-CT scanner (model: XT H 225) at the Finnish Museum of Natural History (Luomus, University of Helsinki) and rendered images along with the videoclips (Fig. 5, Suppl. materials 1, 2) following a non-destructive protocol we have previously described in detail (Englund et al. 2024).
After detaching an antenna from the paratype of Fynbosia horingaria sp. nov. (Fig. 3F), it was cut into two pieces. The proximal part was attached dorsal side up, and the distal part ventral side up with two-sided tape on a metal pod for scanning electron microscopy (SEM) imaging. The preparates were platinum-coated and scanned with Zeiss Gemini device (model: 460 FEG) in the University of Helsinki Institute of Biotechnology Electron Microscopy Unit (Fig. 7).
Following imaging, we removed the abdomens from the holotypes, dissected the genitalia using established preparation protocols (e.g., Hardwick 1950; Robinson 1976; Sihvonen 2001, 2005) stained the preparates with Chlorazol Black, and after photography, mounted them in Euparal. The structural characters of the male genitalia were photographed during dissection through a Leica stereo microscope (model: DM1000 LED) and an attached Leica camera (model: MC170 HD) for scrutiny and illustration of the target structures from a desired angle. We stacked and edited the multi-plane microscope photos using the same software as for the macro photos (Figs 6, 8).
Genomic DNA was extracted from leg tissues of the dry holotype specimens using the Qiagen DNeasy Blood and Tissue Kit following the manufacturer’s protocol at the Luomus DNA laboratory. DNA amplification and sequencing were carried out following the protocols we have described earlier (Englund et al. 2024). One mitochondrial gene (COI) and six protein-coding nuclear gene regions, Arginine Kinase (ArgK), sarco/endoplasmic reticulum calcium ATPase (Ca-ATPase), elongation factor 1 alpha (EF-1alpha), sorting nexin-9-like (Nex9), ribosomal protein (RpS5) and wingless (wgl) were successfully sequenced from the holotypes of F. horingaria sp. nov. (ME09) and F. unicaria sp. nov. (ME81). The sequences for each gene were aligned using the Muscle algorithm, and pairwise genetic distances were calculated using the Kimura 2-parameter model (Kimura 1980) for the COI gene (Suppl. material 3) in MEGA11 (Tamura et al. 2021). The aligned DNA sequences were then uploaded to VoSeq (Peña and Malm 2012). These sequences were combined with a multigene dataset published by Murillo-Ramos et al. (Murillo-Ramos et al. 2019) and pruned to a final dataset of 338 terminal taxa (Fig. 4), representing all subfamilies of Geometridae and an outgroup taxon (Pseudobiston pinratanai Inoue, 1994). The total length of the final dataset was 4980 bp, including gaps. GenBank accession numbers for the newly generated sequences are as follows: PV988405–PV988416.
Breakdown of terminal taxa represented in the pruned and final phylogenetic tree, including the two new Fynbosia species.
The aligned gene sequences from VoSeq were downloaded and used for a maximum likelihood phylogenetic tree construction in IQ-TREE web server (v. 1.6.12) (Nguyen et al. 2015). We applied ModelFinder with ‘-m MFP + MERGE’ option for a best-fitting substitution model (Kalyaanamoorthy et al. 2017). The phylogenetic analyses were carried out using the edge-proportional ‘-spp’ option, which allowed each partition to have its own evolutionary rate. We applied ultrafast bootstrap (UFBoot2) approximations (Guindon et al. 2010; Hoang et al. 2018) using ‘-B 1000 -alrt 1000’ option to evaluate the node supports. To limit the risk of overestimating branch supports in ultrafast bootstrap approximation, we used the ‘-bnni’ option, which optimises each bootstrap tree using a hill-climbing nearest-neighbour-interchange search. The resulting trees we visualised and rooted in FigTree (v. 1.4.4) (Rambaut 2018) and modified the tree in CorelDraw (v. 25.1.0).
Results
The wing venation of F. horingaria sp. nov. (Fig. 5A) exhibits multiple features synapomorphic to the subfamily Larentiinae. Hindwing veins Sc + R1 and Rs are fused for a long-distance, forewing radial veins Rs1– Rs4 arise from the areole and are stalked, M1 proceeds in line with the anterior margin of the discal cell, and the sub-costal accessory cell between veins Sc and R is missing (Murillo-Ramos et al. 2021 and references therein).
Micro-CT images of structural characters of Fynbosia horingaria. A. Wing venation; B. Head, ventral view.
The proboscis and the labial palpi, which comprise just two palpomeres (Fig. 5B), are vestigial, differing from those of most genera in Larentiinae. The cavity of vom Rath’s organ is present at the terminal tip of the labial palpus (Fig. 5B). A micro-CT 3D footage of the entire holotype of F. horingaria sp. nov. is shown in Suppl. material 1, and a close-up from the head exhibiting the proboscis and labial palpi in Suppl. material 2 from multiple view angles.
The general appearance of the two Fynbosia species, wing shape, small size (Figs 3, 6) and resting position (Fig. 2D) would suggest an affiliation to the tribus Eupitheciini in Larentiinae (Viidalepp 2011), but the extremely long rami of the male antennae, the vestigial proboscis and labial palpi, and the male genitalia indicate otherwise. The male genitalia of Fynbosia are generally weakly sclerotised and contain few diagnostic features. The main diagnostic characters of the wing ornamentation and the male genitalia of F. horingaria sp. nov. and F. unicaria sp. nov. are highlighted in Fig. 6 and the keys below.
Key diagnostic characters of Fynbosia species. A, C. Wing ornamentation, dorsal view above, ventral view below; B, D. Male genitalia, caudal view.
Key 1 to species of Fynbosia gen. nov. based on male wing pattern
**: **
Key 2 to species of Fynbosia gen. nov. based on male genitalia morphology
**: **
The male antennae of Fynbosia are disproportionately large and complex compared to those of most other genera in Larentiinae (Fig. 7). The extensive branching and cuticular folding (Fig. 7E, F) of the rami imply a highly sensitive male chemoreception system.
Scanning electron microscope (SEM) images of the antenna of Fynbosia horingaria sp. nov. A. Proximal half of the antenna, dorsal view; B. Distal half of the antenna, ventral view; C. Scale-covered flagella, dorsal view; D. Rami branching pairwise from the ventral side of flagellomere; E. Crumpled cutis of the ramus; F. Surface structure of the cilium. Magnification 250–3000×.
The male genitalia of Fynbosia are generally weakly sclerotised, exhibiting few diagnostic details (Fig. 8), yet the two known species can be distinguished by the shapes of the valva and saccus. We also attempted to render images of the male genitalia from the micro-CT scans, but in the absence of contrast-enhancing sample preparation methods applicable only to fresh specimens, we found the resulting images of insufficient quality to be presented here. The tympanal organs found on the ventral side of the abdominal segments 1+2 are rather small (Fig. 8E) with a slightly swollen tip of the ansa (Fig. 8D).
Dissected Fynbosia abdomens. A. Fynbosia horingaria sp. nov. (holotype) male genitalia, caudal view; B. Uncus, ventro-lateral view; C. Aedeagus; D. Tympanal organ; E. Abdominal skin; F. Fynbosia unicaria sp. nov. (holotype) male genitalia; G. Tip of uncus, dorsal view; H. Aedeagus; I. Uncus, ventro-lateral view; K. Tip of uncus, lateral view.
The multigene molecular phylogeny (Fig. 9) supported our assessment based on morphological characters that Fynbosia moths in the family Geometridae and subfamily Larentiinae constitute an isolated lineage without close affiliation to any described tribus.
Multigene maximum likelihood phylogeny of the Fynbosia gen. nov., showing its placement within Geometridae.
The COI genetic distance between the two Fynbosia species was 5%, and to their nearest neighbours (Rheumaptera hastata (Linnaeus, 1758) and Xanthorhoe abrasaria (Herrich-Schäffer, 1855)) in BOLD was more than 9%. The intraspecific genetic variance in Fynbosia horingaria sp. nov. (N = 3) is less than 1% (Suppl. material 3).
Description of new taxa
Fynbosia
Taxon classificationAnimaliaLepidopteraGeometridae
Englund, Staude & Sihvonen gen. nov.
133DC6CD-3E41-5CE9-B9BB-56A878C73619
https://zoobank.org/4D24E751-A39F-4DA7-9116-B86EB49C52BC
Type species.
Fynbosia horingaria Englund, Staude & Sihvonen, sp. nov.
Diagnosis.
Southern African genus of subfamily Larentiinae (Lepidoptera: Geometridae) present at Cape Floristic Region not attributable or closely related to any described tribus of Larentiinae. Nocturnal small-sized moths with conspicuous male antenna with long ciliated rami, female antenna filiform. Vestigial, defunct proboscis and minuscule two-segmented labial palpi. Adult resting position, small size and elongated fore wing shape resembling those present in genera of Eupitheciini, but the structures of male antenna, proboscis, labial palpi, genitalia, and molecular phylogeny not compatible to any genera of that tribus.
Barcode (COI) 658 bp sequence minimum K2P distance to closest species (Rheumaptera hastata and Xanthorhoe abrasaria) > 9%. Closest sister lineage Brabirodes (Brabirodini) based on multigene 4980 bp maximum likelihood phylogenetic estimation.
Etymology.
Fynbos (> Afrikaans = fine bush) refers to the biome of Cape Floristic Region, where the type taxa reside.
Fynbosia
horingaria
Taxon classificationAnimaliaLepidopteraGeometridae
Englund, Staude & Sihvonen sp. nov.
8C6C69E9-E369-591B-BEE0-C3BC82D0A2F1
https://zoobank.org/03D251B8-9E65-4F92-B168-71F803D317B5
Type material.
Holotype • ♂ (Figs 3A, 3C, 5 A, 6 A), South Africa: Calitzdorp, Swartberg, farm Zandrivier, 33.373°S, 21.330°E, 1200 m, 28 Apr. 2022, Mikael Englund leg., DNA sample ME09, gen. prep. PS 2872, micro-CT scan MS 2, coll. Ditsong Museum of Natural History, Pretoria, South Africa. Paratypes (5♂♂) • ♂ (Fig. 3E), South Africa: Western Cape, 40 km SE of Calitzdorp, farm Zandrivier, dry renosterveld – nama karoo 33.351°S, 21.317°E, 1200 m, 28–29 Apr. 2022, Mikael Englund, Pasi Sihvonen, Hermann Staude leg., DNA sample PS 1641, photo PMS_31002, coll. Englund, Finland; • ♂ (Fig. 3F), South Africa: Western Cape, 40 km SE of Calitzdorp, farm Zandrivier, dry renosterveld – nama karoo 33.351°S, 21.317°E, 1200 m, 28–29 Apr. 2022, Mikael Englund, Pasi Sihvonen, Hermann Staude leg., DNA sample PS 1659, photo PMS_31003, micro-CT scan MS 184, coll. Sihvonen, Finland; • ♂ South Africa: Western Cape, Calitzdorp, Seweweekspoort, Aristata, riverine fynbos, 33.4057°S, 21.4017°E, 12 Apr. 2023, A. Hofmann & J. Meineke, coll. Staude, South Africa; • ♂ South Africa: Western Cape, Calitzdorp, Seweweekspoort, Aristata, riverine fynbos, 33.4057°S, 21.4017°E, 12 Apr. 2023, A. Hofmann & J. Meineke, coll. State Museum of Natural History Stuttgart, Germany.
Diagnosis.
Adult male F. horingaria sp. nov. distinguishable from F. unicaria sp. nov. based on the wing patterns and genitalia. Both fore- and hindwing medial lines evenly rounded, not sharply angled near costa as in F. unicaria sp. nov., best visible on the wing undersides. Discal spots prominent, blackish, triangular in forewings (Figs 3, 6; Key 1). Male genitalia weakly sclerotized, but distinguishable from F. unicaria sp. nov. by the broader valva and elongated saccus (Figs 6, 8; Key 2). Average mtCOI genetic distance to F. unicaria sp. nov. is 5%.
Description.
Adults.
Head: Frons and vertex covered with light brown scales, mixed with some scattered dark brown scales, a string of hair-like scales protruding at the posterior margin of head, compound eye naked. Labial palpi short, with just two segments (Fig. 5B), length less than 1/5 of eye diameter, oriented ventrally, covered with yellowish ochreous scales; vom Rath’s organ cavity on the tip of the terminal palpomere. Proboscis vestigial, hardly visible between labial palpi, apparently defunct. Male antenna (Figs 3, 6, 7) bipectinate, flagella dorsally covered with ochraceous cells, flagellomeres light brown, except for the blackish swollen end of each flagellomere, ramus black, conspicuously long, length up to twice the diameter of compound eye, abundant long cilia covering the ramus. Female antenna filiform.
Thorax: Covered with mixed light to dark brown scales. Spur pattern on legs 0–0–2.
Abdomen: Segments covered with brown scales of varying tone, mottled with blackish scales. Some long hair-like scales extending distally from the proximal segments of the male abdomen. Bowl-shaped paired tympanal organs laterally in the segments 1+2, tip of ansa slightly swollen.
Wings (Figs 3A, 3C, 5A, 6A): Forewing triangular, elongated, termen slightly convex from apex until midwing (M3), tornus round, without distinct transition from termen to dorsum. Wingspan 18–21 mm. Forewing ground colour ranging from light to medium brown with several dark wavy transverse lines most pronounced at costa. Proximal parts of medial and subcostal areas lighter brown than rest of forewing. Basal area with hazel brown tone. Discal spot distinct, triangular, blackish. Three dark brown blotches in terminal area, largest situated medially, smaller ones towards apex and tornus. Ante- and postmedial lines dark, evenly rounding inwards near costa in both fore- and hindwings. Hind wing apex rather sharply angled, discal spot dark. Terminal area lighter with darker suffuse plots forming discontinuous wavy line. Markings on hindwings more distinct on underside.
Genitalia: Male (Figs 6B, 8A–E): Genitalia generally weakly sclerotised with few diagnostic ornaments, except for uncus and gnathos. Valva wide, costa evenly rounded. Uncus rather well sclerotised, prominent with evenly downward hooking tip, juxta sclerotised with saddle-form upward hooking tip. Saccus elongated, aedeagus lacking cornuti.
Female genitalia unknown.
Immature stages unknown.
Genetic data.
GenBank accession numbers PV988405–PV988410.
Etymology.
Horing (> Afrikaans = horn); pertaining to the large and conspicuous, plume-like antennae of the male.
Fynbosia
unicaria
Taxon classificationAnimaliaLepidopteraGeometridae
Englund, Staude & Sihvonen sp. nov.
D3AD5F36-BA32-5667-AFD9-145402272C63
https://zoobank.org/C8FD0470-84AE-4161-B1CC-E46208F9D241
Type material.
Holotype • ♂ (Figs 3B, 3D, 6C), South Africa: Northern Cape, Calvinia, Hantamsberg, Van Rhynshoek, 33.377°S, 19.785°E, 1577 m, Renosterveld gonnabos, 31 Mar. 2022, leg. H.S. Staude, DNA sample ME81, gen. prep. PS 2919, micro-CT scan MS 125, coll. Ditsong Museum of Natural History, Pretoria, South Africa.
Diagnosis.
Adult male F. unicaria sp. nov. distinguishable from F. horingaria sp. nov. based on the wing patterns and genitalia. Both fore- and hindwing medial lines sharply angled near costa, not evenly rounded as in F. horingaria sp. nov., best visible on the wing undersides. Discal spots suffuse, dark, streak-like in forewings (Figs 3, 6; Key 1). Male genitalia weakly sclerotized, but distinguishable from F. unicaria sp. nov. by the narrower valva and short, blunt saccus (Figs 6, 8; Key 2). Average mtCOI genetic distance to F. horingaria sp. nov. is 5%.
Description.
Adults (male).
Head: Frons and vertex covered with light brown scales, string of hair-like scales protruding at posterior margin of head, compound eye naked. Labial palpi extremely short, length less than 1/5 of eye diameter, oriented ventrally, hardly visible between eyes, proboscis vestigial, defunct. Male antenna (Figs 3B, 3D, 6C) bipectinate, proximal third of flagella dorsally covered with dark brown scales, two distal thirds covered with ochraceous cells scattered with darker brown scales, ramus black, conspicuously long, length up to 1.5 times diameter of compound eye, abundant long cilia covering ramus.
Thorax: Covered with brown scales of varying tone. Spur pattern on legs 0–0–2.
Abdomen: Segments covered with brown scales of varying tone, mottled with blackish scales. Some long hair-like scales extending distally from proximal segments of male abdomen.
Wings (Figs 3B, 3D, 6C): Forewing triangular, elongated, narrow, costa and termen almost straight, tornus round, without distinct transition from termen to dorsum, wingspan 20 mm. Forewing ground colour grey-brown, medial and terminal areas darker than rest of forewing, postmedial line almost straight, bordered distally with broad, suffuse hazel brown margin, turning inwards at sharp angle before touching costa. Discal spot stripe-like, black. Hindwings echoing forewing ornamentation pattern in less pronounced manner; postmedial line sharply angled, discal spot small, round, black. Dark, suffuse blotches in terminal area.
Genitalia: Male (Figs 6D, 8F–K): Genitalia generally weakly sclerotised and simple, except for the tip of gnathos. Few diagnostic features. Valva narrow, costa straight. Saccus short, borders almost straight, forming roughly right angle at tip. Aedeagus without well-sclerotised cornuti.
Female and immature stages unknown.
Genetic data.
GenBank accession numbers PV988411–PV988416.
Etymology.
Unicus (> Latin = sole, only); pertaining to the rarity and solitary known specimen of the species at the time of description.
Discussion
There are close to 24000 described species of geometrid moths worldwide, and according to recent estimates, at least the same number of species remain undescribed (Rajaei et al. 2022). Within southern Africa, more than 2000 species of geometrid moths are present in extant collections, about one-third of which remain undescribed (Staude et al. 2025).
Against this background, the discovery of two new species from the poorly investigated fynbos is not particularly surprising. However, more surprising is that the new taxa do not fit well within the current classification system and apparently represent a phylogenetically isolated lineage. We consider our data adequate to support classifying Fynbosia in a monogenetic tribe within Larentiinae, but refrain from doing so because the taxon sampling underlying the current multigene molecular phylogenies is rather limited, particularly from the tropical areas. We expect the addition of new taxa will provide more robust support for the phylogenetic position of Fynbosia within the basal Larentiinae in the Geometridae tree of life.
We also examined the morphology of Brabirodes peruviana, the terminal taxon in our phylogeny (Fig. 9) as the closest relative to Fynbosia and found several morphological differences in B. peruviana compared to both Fynbosia species. For instance, the wing venation, male genitalia and tympanal organs in these species exhibit clearly distinct morphological features, as described in greater detail in a forthcoming publication (Sihvonen et al. 2025).
The stable geological and climatological conditions in the Cape region of South Africa have provided a unique setting for the floral and animal endemism in the Cape (Samways et al. 2024b). The elevated mountain slopes often support a flora distinct from the surrounding arid Karoo, rendering these biomes effectively isolated from one another. Especially phytophagous insect populations with likely limited dispersal, residing at higher elevations, such as Fynbosia moths, may be effectively allopatric.
Conclusions
We experimented with the rendering of non-destructive diagnostic images from inner organs and structures from micro-CT scans (Englund et al. 2024) and found that we could produce adequate-quality images of wing venation, proboscis, and labial palpi, saving the rare specimens from further destruction. However, we assessed the micro-CT images of the tympanal organs and genitalia as insufficient for publication here due to weak natural sclerotization, and also to avoid destructive sample preparation methods.
While the life history, ecology and deeper phylogenetic relationships of Fynbosia remain largely unstudied at present, the new genus has some interesting traits, such as a vestigial proboscis, truncated labial palpi, and an apparently highly efficient male chemoreception apparatus, present in few other genera of the subfamily. The life history and tribal-level phylogeny for Fynbosia, as well as a few neighbouring genera, merit further study.
Supplementary Material
XML Treatment for Fynbosia
XML Treatment for Fynbosia horingaria
XML Treatment for Fynbosia unicaria
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Born J, Linder HP, Desmet P (2007) The Greater Cape Floristic Region. Journal of Biogeography 34(1): 147–162. 10.1111/j.1365-2699.2006.01595.x · doi ↗
- 2Englund M, Lee KM, Staude H, Duplouy A, Hausmann A, Laiho E, Söderholm M, Sihvonen P (2024) 130 years from discovery to description: micro-CT scanning applied to construct the integrative taxonomy of a forgotten moth from Southern Africa (Lepidoptera: Geometridae). Systematic Entomology 49(3): 507–525. 10.1111/syen.12627 · doi ↗
- 3Forest F, Colville JF, Cowling RM (2018) Evolutionary Diversity Patterns in the Cape Flora of South Africa. In: Scherson R, Faith D (Eds) Phylogenetic Diversity. Springer, Cham, 167–187. 10.1007/978-3-319-93145-6 · doi ↗
- 4Grobler BA, Cowling RM (2021) The composition, geography, biology and assembly of the coastal flora of the Cape Floristic Region. Peer J 9(e 11916): 1–52. 10.7717/peerj.11916 PMC 836432634447627 · doi ↗ · pubmed ↗
- 5Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of Phy ML 3.0. Systematic Biology 59(3): 307–321. 10.1093/sysbio/syq 01020525638 · doi ↗ · pubmed ↗
- 6Hardwick DF (1950) Preparation of Slide Mounts of Lepidopterous Genitalia. Canadian Entomologist 82(11): 231–235. 10.4039/Ent 82231-11 · doi ↗
- 7Hoang DT, Chernomor O, Von Haeseler A, Minh BQ, Vinh LS (2018) UF Boot 2: Improving the ultrafast bootstrap approximation. Molecular Biology and Evolution 35(2): 518–522. 10.1093/molbev/msx 281PMC 585022229077904 · doi ↗ · pubmed ↗
- 8Kalyaanamoorthy S, Minh BQ, Wong TKF, Von Haeseler A, Jermiin LS (2017) Model Finder: Fast model selection for accurate phylogenetic estimates. Nature Methods 14(6): 587–589. 10.1038/nmeth.4285 PMC 545324528481363 · doi ↗ · pubmed ↗