Spring cold stress at high altitudes in southeastern Xizang activates CsABF2 to regulate chlorophyll degradation and phenolic biosynthesis in tea plants
Yipeng Huang, Didi Jin, Tianming Jiao, Zhenhong Wang, Ting Jiang, Lei Zhao, Xiaolan Jiang, Haiyan Wang, Yajun Liu, Yunsheng Wang, Liping Gao, Tao Xia

TL;DR
Tea plants in high-altitude regions experience cold stress, which affects their growth and quality through changes in chlorophyll and phenolic compounds.
Contribution
The study identifies CsABF2 as a key transcription factor regulating cold stress responses in tea plants at high altitudes.
Findings
Cold stress at high altitudes delays budburst and causes leaf yellowing in tea plants.
CsABF2 activates genes involved in chlorophyll degradation and phenolic biosynthesis under cold stress.
Transcriptome analysis reveals altered pathways in response to fluctuating low temperatures.
Abstract
The tea plant (Camellia sinensis), native to warm and humid low-latitude regions of southwestern China, has expanded to higher altitudes, including southeastern Xizang, where cultivation above 2500 m poses challenges due to low accumulated temperatures. However, the impact of high-altitude climatic conditions, particularly temperature, on tea growth remains underexplored. To investigate, weather stations were deployed at three altitudes in southeastern Xizang to monitor spring temperature fluctuations: Medog (MD, 1200 m), Zayü (ZY, 1720 m), and Layue in Bayi District (BY, 2600 m). Field observations and meteorological data indicated that the milder spring temperatures in MD and ZY facilitated normal budburst and growth, whereas the lower temperatures in BY delayed budburst and resulted in leaf yellowing and browning. Comparative experiments revealed that seedlings exposed to fluctuating…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
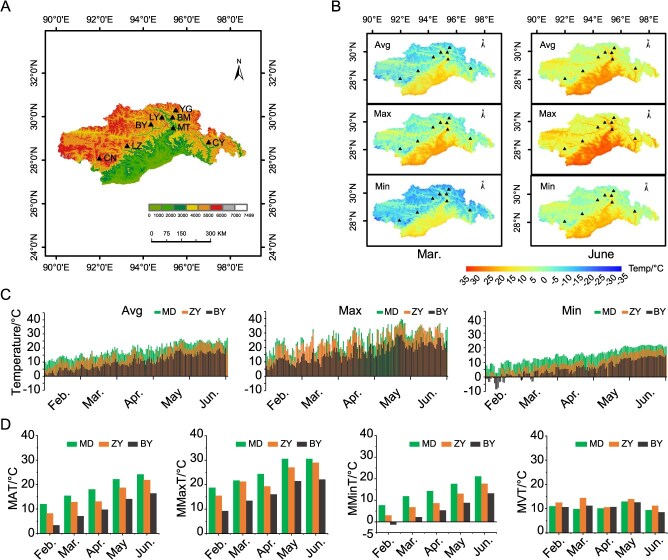 Figure 1
Figure 1 Figure 2
Figure 2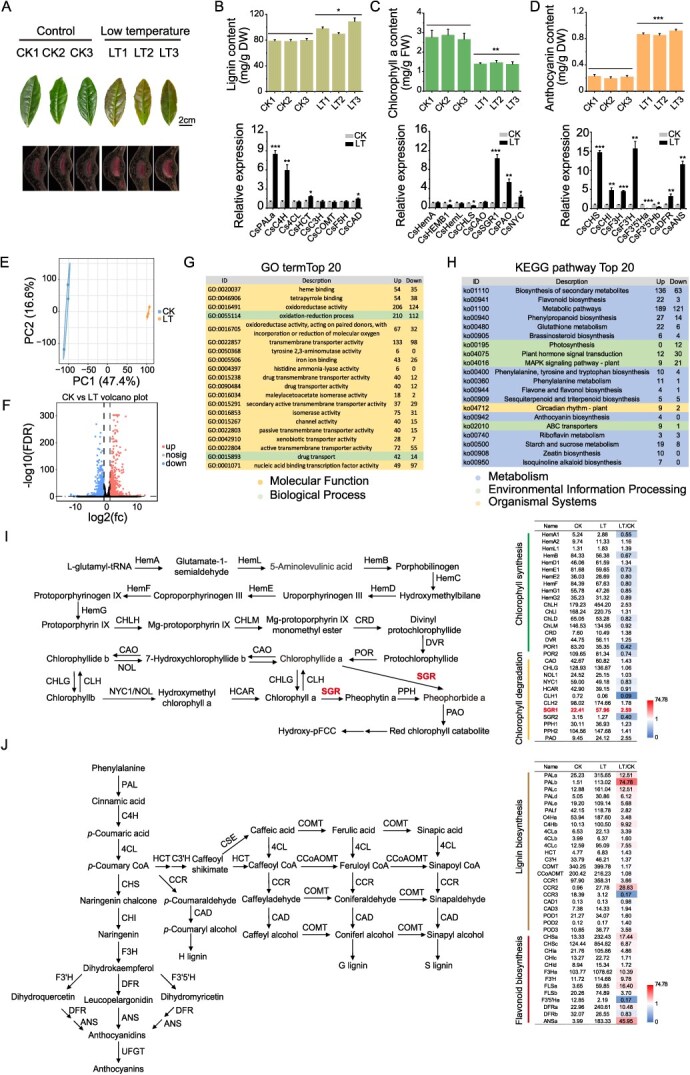 Figure 3
Figure 3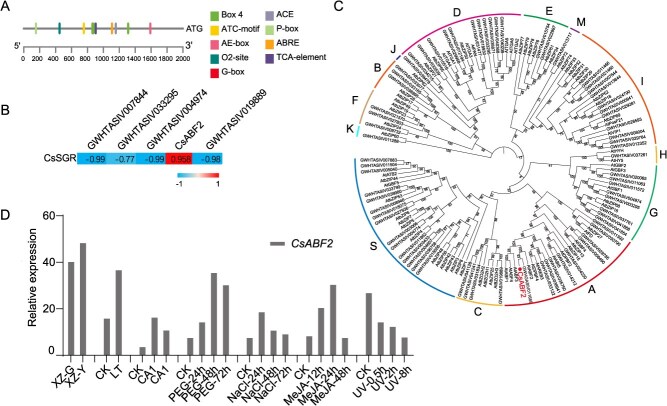 Figure 4
Figure 4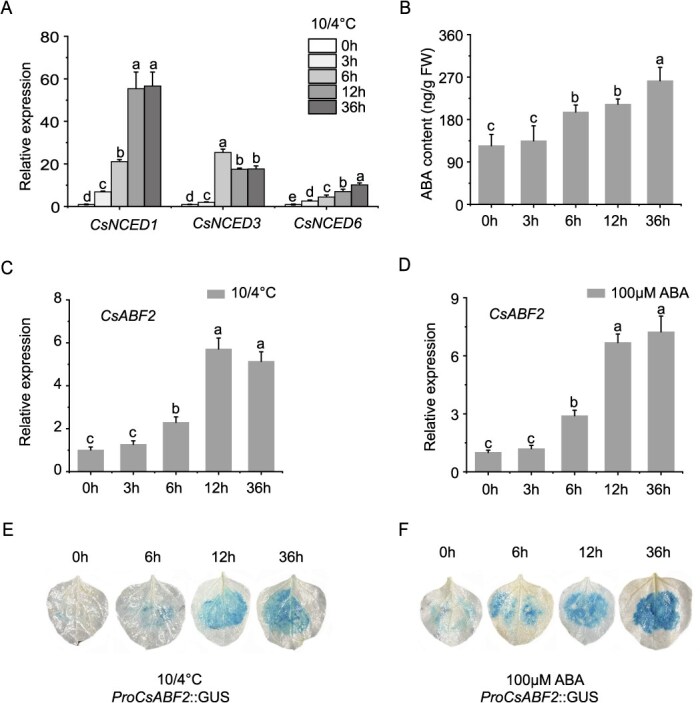 Figure 5
Figure 5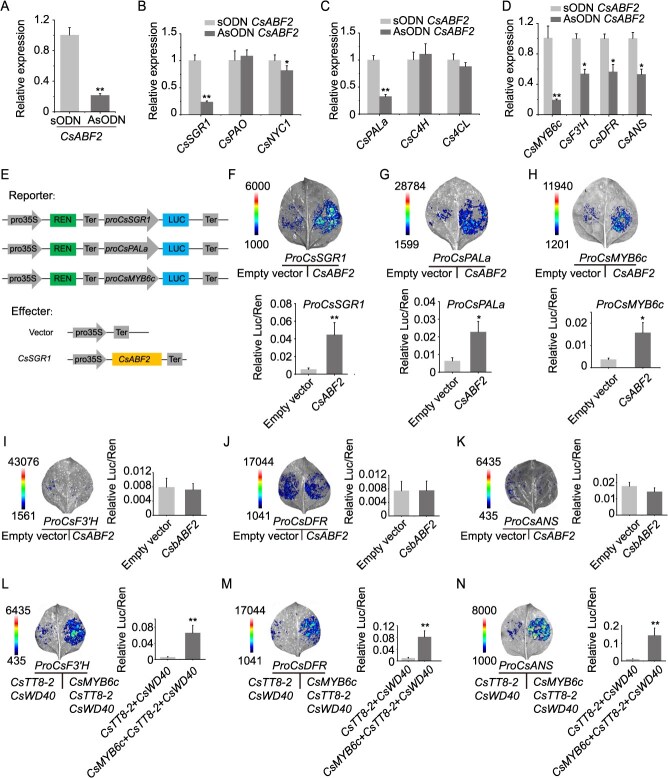 Figure 6
Figure 6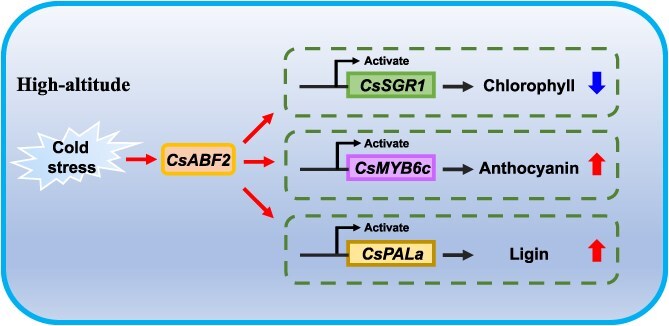 Figure 7
Figure 7- —National Key Research and Development Program of China10.13039/501100012166
- —Natural Science Foundation of China10.13039/501100001809
- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTea Polyphenols and Effects
Introduction
The tea plants, originally adapted to southwestern China, thrives in warm, humid, and shaded environment [1, 2]. Through cultivation expansion, tea has gradually migrated to higher latitudes and altitudes, undergoing adaptive changes in response to progressively colder climates [3]. Climatic factors, particularly temperature, play multifaceted roles in influencing the survival of tea plants and the accumulation of quality-related components [4–7]. From the perspective of germplasm, the primary concern is the impact of low temperatures in winter on plant survival. Tea cultivars vary significantly in cold tolerance and can endure temperatures as low as −10 to −15°C [8]. Also, cultivation has even extended as far north as 49°N in Ukraine [9]. Another important consideration is the effect of low temperatures in spring on bud burst timing and the development of quality traits.
Early spring temperatures around 0°C can lead to cold damage or frost injury and pose serious challenges to tea growth, especially during cold snaps following premature warming [6, 10, 11]. While extensive research has been conducted on the physiological and molecular mechanisms of cold stress and low-temperature adaptation in tea plants [7, 12–14], factors such as the duration of low temperatures (above 0°C) in early spring, large diurnal temperature variations, and slow warming rates in high-latitude and high-altitude regions have received little attention, despite their significant impact on tea production.
Southeastern Xizang, influenced by warm, humid air currents from the Indian Ocean and characterized by its complex terrain of mountainous river valleys, exhibits a significant altitudinal gradient [15]. Variations in altitude bring about significant changes in multiple environmental factors, including temperature, humidity, oxygen availability, and light intensity [16, 17]. These environmental gradients impose considerable challenges on plant growth and survival. In high-altitude environments, plants have developed various mechanisms to cope with the adverse conditions present in such settings. Morphologically, most alpine plants exhibit thickened waxy cuticles on their leaves to mitigate damage caused by low temperatures, suppression of vegetative growth, increase in stomatal conductance, enhancement of photosynthetic efficiency, and dwarfing to minimize exposure to wind and cold [16]. Physiologically, alpine plants enhance the production of osmoregulatory substances to prevent damage from freezing or ice crystal formation [18]. High-altitude plants possess a stronger reactive oxygen species scavenging capacity, and the elevation-dependent increase in proline and soluble sugar contents suggests that these compounds play an important role in osmotic regulation [19].
To adapt to environmental stresses, plants enhance their secondary metabolism as a defense strategy [16]. Phenolic compounds, such as flavonoids and phenolic acids, increases with elevation [20, 21]. The mechanisms through which certain novel flavonoid biosynthesis gene clusters modulate flavonoid production and enhance adaptation to high-altitude environments [22]. Additionally, some alpine plants enhance the accumulation of B-ring trihydroxy anthocyanins, deepening leaf or flower coloration to prevent ultraviolet radiation damage [23]. Moreover, terpenoids, including monoterpenes, sesquiterpenes, diterpenes, and triterpenes and as well as volatile secondary metabolites, accumulate significantly under high-altitude conditions [24–26]. In summary, these integrated physiological and morphological strategies reflect long-term evolutionary adaptations to high-altitude environments and provide significant insights into the potential mechanisms of ecological resilience in plants. In summary, these integrated morphological strategies reflect long-term evolutionary adaptations to high-altitude environments.
Tea plants cultivated at higher altitudes in southeastern Xizang display adaptive traits, including a shortened growth period and reduced survival rates of cuttings in early spring. The tea plants cultivated in this region are subjected to diverse ecological conditions, providing valuable material for studying the effects of environmental differences on tea plant growth, development, and the accumulation of quality-related components. Furthermore, this serves as a basis for investigating how high-altitude environments affect the aroma and taste characteristics of tea plant [27].
This study, based on monitoring data provided by the National Tibetan Plateau/Third Pole Environment Data Center, as well as observations from miniature weather stations and records from tea cultivation, examines how temperature variations in spring across different altitudes in southeastern Xizang affect the growth of tea seedlings. Additionally, by simulating low-temperature conditions during early spring in a controlled climate chamber, we employed transcriptomic and molecular biology techniques to investigate the causes of cold-induced leaf damage and the functional role of the cold-responsive gene CsABF2. In summary, the CsABF2 transcription factor induced by low temperature not only promotes the degradation of chlorophyll in tea leaves but also facilitates the biosynthesis of lignin and anthocyanins. This study enhances our understanding of the effects of low temperatures at high altitudes on the yellowing of tea leaves and the accumulation of secondary metabolites.
Results
Climatic characteristics of southeastern Xizang and cold injury in spring for tea plants
This study first downloads the GeoJSON format data corresponding to the required administrative divisions from the official Tianditu website (https://www.tianditu.gov.cn, Map Approval Number: GS (2024)0650), and the map with the approval number is shown in Fig. S1. Subsequently, the JSON format data is converted into ESRI Shapefile (.shp) using Mapshaper (https://mapshaper.org). After completion of the conversion, the shapefile is imported into ArcMap 10.8.1 for further processing. Finally, digital elevation models (DEMs) are constructed for Nyingchi city and Shannan city in Xizang, enabling effective visualization of the locations of tea plantations situated in southeastern Xizang (Fig. 1A). The map reveals that tea cultivation in southern Xizang is primarily distributed between 28°N–31°N latitude and 92°E–97°E longitude. Among these regions, the tea gardens in Medog (MD) are mostly located at the lowest altitudes (average altitude of ~1200 m) and are strongly influenced by warm, humid air masses from the Indian Ocean [28, 29]. Consequently, this area features a low-mountain subtropical humid climate. In contrast, the tea-cultivating region of Zayü (ZY) lies at the confluence of the Himalayas and the Hengduan Mountains, with altitudes ~1700 m. This area experiences a subtropical mountain humid monsoon climate characterized by mild winters, hot summers, and consistently humid, rainy conditions throughout the year. The other tea-cultivating regions are situated at average altitudes above 2000 m and are subtropical mountainous areas characterized by a semi-humid, warm temperate monsoon climate. Among them, Layue in Bayi District (BY) is located at the highest altitude, exceeding 2600 m.
Climatic conditions suitable for tea growth in southeastern Xizang. (A) DEM of tea cultivation regions in southeastern Xizang. The map was obtained from the Tianditu website (https://www.tianditu.gov.cn, Map Approval Number: GS (2024)0650). Tea cultivating locations in southeastern Xizang include Medog county (MD), Bomi county (BM), Zayü county (ZY), Bayi district (BY), Cuona county (CN), Longzi county (LZ), Layue (LY), and Yigong tea garden (YG); (B) spatial distribution of temperature in March and June (2014–2023) across tea-cultivating areas in southeastern Xizang, temperature includes average temperature (Avg), maximum temperature (Max), and minimum temperature (Max); (C) diurnal variation of average air temperature in spring 2024 at the three tea gardens in southeastern Xizang; and (D) monthly variation of average air temperature in spring 2024 at the three tea gardens, monthly average temperature (MAT), monthly maximum temperature (MMaxT), monthly minimum temperature (MMinT), and monthly variation temperature (MVT).
Monthly average temperature, minimum temperature, and maximum temperature datasets with a spatial resolution of 1 km were obtained from the National Tibetan Plateau/Third Pole Environment Data Center (https://data.tpdc.ac.cn) [30–33]. With ArcMap 10.8.1, spatial distribution maps of average, minimum, and maximum temperatures for March and June over the past decade (2014–2023) in southeastern Xizang were generated (Fig. 1B). The results indicate that the low-altitude regions of MD and ZY exhibit higher monthly average, minimum, and maximum temperatures compared to the high-altitude regions in both March and June. Additionally, the data reveal that some valley areas within the high-altitude regions experience higher temperatures than the surrounding mountainous terrain.
In most tea-cultivating regions of the Northern Hemisphere, tea buds typically begin to sprout between March and April each year. An average spring temperature of 10°C is generally considered the threshold for budburst. During this critical period, low monthly minimum temperature (MMinT) or sudden extreme cold events can lead to chilling or freezing injury. In order to closely examine variations in springtime temperatures during the budding season across tea gardens at different altitudes, compact weather stations were installed in tea gardens at MD, ZY, and BY tea. These stations recorded daily fluctuations in air temperature from February to June 2024 (Fig. 1C; Table S1), along with variations in monthly average temperature (Fig. 1D). The results showed a clear trend of decreasing monthly mean temperature (MAT), monthly maximum temperature (MMaxT), and MMinT with increasing altitude. In March, the low-altitude MD garden recorded a monthly minimum average temperature of 11.89°C, a maximum of 21.72°C, and an average temperature of 15.36°C. The ZY garden recorded a minimum of 6.80°C, a maximum of 21.24°C, and an average of 12.78°C. In contrast, the BY Garden at high altitude had MMinT only 2.23°C, MMaxT was 13.44°C, and MAT was 7.12°C. These findings indicate that budburst occurs earliest in the MD garden and latest in the BY garden. Based on temperature patterns, it can be inferred that budburst in the high-altitude garden at BY likely occurs after May and is highly susceptible to freezing injury.
In order to observe the effect of high-altitude climate on the growth and development of tea plants, in 2022, we planted 20 one-year-old tea plant varieties from Anhui, Zhejiang, and Fujian at three distinct altitudes in Xizang: low-altitude region ZY (1720 m), and high-altitude Yigong (YG, 2200 m), and BY (2600 m). Through comparative cultivation experiments, this study aimed to investigate whether the high-altitude climate induces frost damage in tea plants and to identify tea plant varieties that are suitable for cultivation in these conditions. The growth of the seedlings was assessed in mid-May two years later, and images of seven cultivars were selected for presentation (Fig. S2A). Compared with the vigorous growth observed in ZY, seedlings at the higher altitude sites (YG and BY) exhibited symptoms across all cultivars. Specifically, the young apical leaves showed uneven yellowing, while the older basal leaves developed browning. These symptoms gradually subsided as temperatures increased (Fig. S2B). In summary, these observations suggest that the slow rise in spring temperatures (April–May) at high altitudes is likely a major environmental factor limiting the growth of tea plants in this region.
According to the variation in diurnal temperature (Fig. 1C), the minimum temperature in the high-altitude region of BY often falls below 10°C, and the average difference between daily maximum and minimum temperatures reaches ~10°C. Based on this, we hypothesize that large fluctuations in diurnal temperatures may be a primary cause of cold injury in tea plants grown at high altitudes. To test whether low temperatures (4–10°C) contribute to cold injury in tea plants, we designed three controlled day/night temperature treatments: 4°C/4°C (constant low temperature), 10°C/4°C (diurnal fluctuation), and 10°C/10°C (constant moderate temperature). As shown in Fig. S2C, no symptoms of cold injury were observed in the 10°C/10°C group. In contrast, both the 4°C/4°C and 10°C/4°C treatments resulted in water-soaked lesions characteristic of cold injury. In addition, leaf browning was more significant in the 10°C/4°C treatment than in the 4°C/4°C treatment.
All three groups of treated tea seedlings were maintained at room temperature to observe bud development and assess the impact of cold injury on the yield. As shown in Figs S2D and E, compared with seedlings subjected to the 10°C/10°C treatment, those exposed to 4°C/4°C and 10°C/4°C exhibited significantly reduced weights of fresh shoots (one bud with two leaves) at both the first (15 days at room temperature) and the second (30 days at room temperature) harvests. Notably, the damage from the 10°C/4°C treatment persisted into the second sampling. These findings suggest that although the temperature difference (6°C) was relatively small, diurnal fluctuations between 4 and 10°C caused more severe damage to tea plants than other treatments. This further supports the idea that in high-altitude regions such as BY, day–night temperature fluctuation during the spring (April–May) has a significant impact on the growth of tea plants.
Molecular basis of spring regional yellowing leaf in high-altitude tea plants
In early spring, tea plants in the high-altitude regions of southeastern Xizang exhibit reddening of mature leaves and uneven region-specific yellowing young leaves. To explore the molecular mechanism underlying the regional yellowing tender shoots, this study employed transcriptome sequencing to compare the gene expression profiles between XZ-Y and XZ-G areas of the third leaf from the apical shoot (Fig. 2A).
Transcriptomic insights into leaf yellowing in tea plants grown at high altitudes in Xizang: (A) XZ-Y and XZ-G indicate different leaf areas, respectively; (B) principal component analysis (PCA) of transcriptome profiles; (C) number of differentially expressed genes (DEGs) identified between yellow and green leaf regions; (D) Gene Ontology (GO) enrichment analysis of DEGs; (E) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of DEGs; (F) DEGs in pathways of chlorophyll biosynthesis and degradation; and (G) DEGs in pathways of phenylpropanoid and lignin biosynthesis.
Principal component analysis (PCA) of the transcriptome data showed clear clustering within groups and distinct separation between groups, indicating good intra-group reproducibility and significant inter-group differences (Fig. 2B). Based on screening criteria of FDR < 0.05 and |log₂ fold change| > 1, a total of 2590 genes were identified as upregulated and 5854 as downregulated in the green area relative to the yellow area (Fig. 2C). GO enrichment analysis revealed that the differentially expressed genes (DEGs) were primarily enriched in categories such as plastid (GO:0009536; GO:0009507), plastid envelope (GO:0009526), small molecule metabolic processes (GO:0044281), oxoacid metabolism (GO:0043436), and organic acid metabolism (GO:0006082) (Fig. 2D; Table S2). KEGG pathway analysis showed significant enrichment in secondary metabolism (ko01110) and general metabolic pathways (ko01100), particularly those related to photosynthesis (ko00195), ubiquinone and terpenoid–quinone biosynthesis (ko00130), and phenylalanine metabolism (ko00940) (Fig. 2E; Table S3).
Further analysis of gene expression in the pathways related to chlorophyll metabolism revealed that several genes involved in chlorophyll degradation, Non-Yellow Coloring 1 (NYC1), Chlorophyllase (CLH), Pheophorbide a oxygenase (PAO), and Stay-Green 1 (SGR1), were significantly upregulated in the yellow-spotted areas compared to the green regions, with CsSGR1 exhibiting a 10.3-fold increase (Fig. 2F). According to previous studies, CsSGR1 is crucial in chlorophyll degradation, as plants lacking SGR1 exhibit a stay-green phenotype [34]. Additionally, the study examined gene expression in the phenylpropanoid and lignin biosynthesis pathways (ko00940). The expression of phenylalanine ammonia lyase (PAL), cinnamate 4-hydroxylase (C4H), 4-coumaroyl-CoA ligase (4CL), and shikimate/quinate hydroxy cinnamoyl transferase (HCT) was significantly higher in the region-specific yellowing leaves, especially in the upstream segment of the phenylalanine metabolic pathway (Fig. 2G).
Transcriptomic and metabolic responses of tea plants to indoor low-temperature treatment
To investigate the molecular basis of cold-induced color changes in tea seedlings under low diurnal temperatures, this study employed transcriptome sequencing to analyze the impact of normal diurnal temperatures (25°C/20°C) versus low-temperature stress (10°C/4°C) on gene expression profiles in tea seedlings (Fig. 3A). Quantitative analysis of lignin, chlorophyll, and anthocyanins in seedling leaves revealed that low-temperature treatment significantly altered the metabolic characteristics of tea plants. With phloroglucinol-HCl staining to detect lignin distribution, the low-temperature group exhibited a more intense red coloration, indicating a significantly higher lignin accumulation compared to the control group (Fig. 3A). Further quantitative analysis confirmed that low-temperature treatment not only increased the total content of lignin but also significantly upregulated the expression of key genes in the phenylpropanoid-lignin pathway, i.e., CsPALa and CsC4H (Fig. 3B). In terms of pigment metabolism, low-temperature treatment led to a significant reduction in the content of chlorophyll a in tea leaves, accompanied by significant upregulation of CsSGR1 and CsPAO, genes involved in chlorophyll degradation (Fig. 3C). Also, genes associated with anthocyanin biosynthesis, chalcone synthase (CHS), chalcone isomerase (CHI), flavanone 3-hydroxylase (F3H), dihydroflavonol 4-reductase (DFR), and anthocyanidin synthase (ANS), were all significantly upregulated in the low-temperature group, resulting in a significant increase in anthocyanin content compared to the control group (Fig. 3D).
*Effects of indoor low-temperature treatment on gene expression and metabolism in tea seedlings. (A) Phenotypic variation in the third leaf of tea seedlings after three weeks of low-temperature treatment, including differences in lignin content. Lignin content was visualized by phloroglucinol staining of leaf cross-sections; (B) quantification of lignin content and expression levels of lignin biosynthesis-related genes; (C) chlorophyll content and expression levels of chlorophyll-related genes; (D) anthocyanin content and expression levels of anthocyanin biosynthesis genes; (E) PCA of transcriptome profiles under different temperature conditions; (F) volcano plotting of DEGs between control groups and low-temperature; (G) GO enrichment analysis of DEGs; (H) KEGG enrichment analysis of DEGs; (I) differential expression of genes in pathways of chlorophyll biosynthesis and degradation; and (J) differential expression of genes in pathways of phenylpropanoid and lignin biosynthesis. All data are presented as the mean ± standard deviation (SD) of three biological replicates (n = 3). Statistical significance was determined by the Student’s t-test, with *P < 0.05, **P < 0.01, and **P < 0.001.
Transcriptome sequencing was performed on two groups of samples, and PCA was used to assess the distribution patterns of the data. The analysis revealed that samples within each group were tightly clustered, while those between groups were clearly separated, indicating high intra-group consistency and significant differences between treatments (Fig. 3E). Based on the thresholds of FDR < 0.05 and |log₂FC| > 1, the volcano plot analysis identified 1220 genes as significantly upregulated and 1275 genes as downregulated under low-temperature treatment (Fig. 3F). GO enrichment analysis indicated that among the top 20 enriched terms, 18 were classified under ‘molecular function’ and 2 under ‘biological process.’ Notably, ‘oxidoreductase activity’ (GO:0016491), and ‘oxidation–reduction process’ (GO:0055114) were significantly enriched (Fig. 3G; Table S4). KEGG pathway analysis further revealed that DEGs were predominantly enriched in pathways related to secondary metabolism (ko01110) and primary metabolism (ko01100), specifically involving flavonoid biosynthesis (ko00941), glutathione metabolism (ko00480), phenylalanine metabolism (ko00940), photosynthesis (ko00195), and plant hormone signal transduction (ko04075) (Fig. 3H and Table S5). These results suggest that low-temperature stress may influence photosynthetic efficiency in tea plants and modulate the biosynthetic pathways of lignin and flavonoids.
This study further explored the expression characteristics of genes involved in chlorophyll biosynthesis, phenylpropanoid metabolism, and lignin biosynthesis. Among the genes associated with chlorophyll degradation, CsNYC1, CsCLH1, CsPAO, and CsSGR1 exhibited significantly elevated transcript levels in the yellow-spotted region compared to the green region (control), with CsSGR1 exhibiting the most significant upregulation, 2.59 times higher than the control (Fig. 3I). Similarly, key genes involved in the pathways of phenylpropanoid and lignin biosynthesis, CsPALb, CsC4Hb, Cs4CLc, and CsCCR2, were significantly upregulated, particularly in the early stages of the phenylalanine metabolism, with CsPALb exhibiting the most substantial increase in expression (Fig. 3J). Notably, the change trend of expression levels of some genes related to chlorophyll synthesis and degradation, and lignin synthesis was consistent between samples from cold-stressed tea plants in Xizang plantations and laboratory-grown seedlings subjected to low-temperature treatment.
Functional characterization of the cold-induced gene CsABF2
Identification and expression analysis of CsABF2 as a potential regulator of CsSGR1 in tea plants
As demonstrated above, the region-specific yellowing observed in tea leaves during spring in high-altitude regions of Xizang is primarily caused by low-temperature conditions. To investigate the molecular basis of this phenomenon, this study focused on identifying transcription factors that regulate chlorophyll content in tea leaves. An analysis of the functional regulatory elements in the promoter region of CsSGR1 identified several key elements associated with light signaling and plant hormone responses (Fig. 4A). Specifically, Box 4, AE-box, and ATC-motif are involved in light responsiveness; G-box and ACE also function in light signaling; ABRE is associated with abscisic acid (ABA) signaling; TCA-element is linked to salicylic acid signaling; and P-box is related to gibberellin response. Previous research has shown that ABF transcription factors, which belong to subgroup A of the bZIP family, are crucial in regulating ABA-mediated abiotic stress responses in plants [35, 36]. In the model plant Arabidopsis thaliana, members such as ABF2, ABF3, and ABF4 significantly regulate ABA-induced chlorophyll degradation and leaf senescence [34]. Based on expression correlation analysis, the expression of bZIP transcription factors exhibited a strong positive correlation with CsSGR1 expression (Fig. 4B; Table S6). Phylogenetic analysis using MEGA7 revealed that the identified CsbZIP is homologous to AtABF2, an ABA-responsive element-binding factor in Arabidopsis, and belongs to the ABF subgroup of the bZIP family (Fig. 4C). Therefore, it was designated as CsABF2. Analysis of publicly available transcriptome datasets from the TPIA and NCBI database (Fig. 4D) showed that CsABF2 expression was significantly altered under various stress conditions. Cold treatment, PEG-induced drought, salt stress, and MeJA application all significantly upregulated CsABF2 expression, whereas UV radiation suppressed it. These results demonstrate that CsABF2 is a stress-responsive gene.
Identification and expression profiling of CsABF2 in tea plants. (A) Cis-regulatory element analysis of the CsSGR1 promoter; (B) correlation analysis between the expression of bZIP transcription factors and CsSGR1; (C) phylogenetic analysis of CsABF2 and its homologs in other plant species; (D) expression of CsABF2 in response to low temperature, PEG-induced drought, salt stress, MeJA treatment, and UV exposure based on publicly available transcriptome datasets (Tea Plant Information Archive (TPIA)).
CsABF2 expression is upregulated by low-temperature and ABA treatments
As a key regulatory enzyme in ABA biosynthesis, 9-cis-Epoxycarotenoid Dioxygenase (NCED) is crucial [37, 38]. In plants, ABA levels are induced in response to various stress signals, primarily through the upregulation of genes encoding enzymes involved in ABA biosynthesis [39]. Among these, NCED is considered to catalyze the rate-limiting step in ABA production [40]. Previous studies have reported that the expression of NCED and ABA2 is upregulated when tea plants are exposed to cold stress [39]. To investigate the involvement of CsABF2 in ABA signaling under cold stress, we analyzed the expression of CsNCEDs gene and ABA accumulation. The results indicated that cold treatment led to an upregulation of CsNCED1, CsNCED3, and CsNCED6, all of which are closely associated with ABA biosynthesis, accompanied by a continuous increase in ABA levels (Fig. 5A and B). Further molecular analysis revealed that both cold stress and exogenous ABA treatment significantly increased the transcriptional level of CsABF2 (Fig. 5C and D). Promoter activity assays in Nicotiana benthamiana further confirmed this response, where GUS staining showed that CsABF2 promoter activity increased in a time-dependent manner following cold and ABA treatments (Fig. 5E and F). The mechanism of CsABF2 gene expression stimulated by low temperature and ABA requires further investigation.
Induction of CsABF2 expression and promoter activity by low temperature and ABA. (A) Low-temperature treatment upregulates the expression of CsNCED1, CsNCED3, and CsNCED6, key genes involved in ABA biosynthesis; (B) low-temperature treatment promotes ABA accumulation; (C) low-temperature treatment enhances CsABF2 transcript levels; (D) exogenous application of 100 μM ABA promotes CsABF2 expression; (E) low temperature activates CsABF2 promoter activity, as shown by GUS staining in N. benthamiana; (F) ABA treatment also induces CsABF2 promoter activity in N. benthamiana. All data are presented as the mean ± standard deviation (SD) of three biological replicates (n = 3). Different letters indicate statistically significant differences at P < 0.05, based on Tukey’s honestly significant difference (HSD) test.
Functional validation of CsABF2 target genes via antisense oligodeoxynucleotide-mediated gene silencing
Antisense oligodeoxynucleotide (AsODN) technology is an effective tool for suppressing gene expression and investigating gene function [41, 42]. In this study, AsODN-mediated knockdown of CsABF2 was conducted in tea leaves, resulting in a significant reduction in CsABF2 expression (Fig. 6A). qPCR analysis showed that suppression of CsABF2 led to a significant decrease in the transcript levels of CsSGR1, which is the key gene related to chlorophyll degradation, and CsNYC1, while the expression of CsPAO remained largely unchanged (Fig. 6B), suggesting that CsSGR1 may be directly regulated by CsABF2. In the pathway of phenylpropanoid biosynthesis, the CsPALa gene also exhibited CsABF2-dependent expression (Fig. 6C). Moreover, the expression of genes involved in anthocyanin biosynthesis, including CsMYB6c, CsF3′H, CsDFR, and CsANS, was significantly regulated (Fig. 6D).
*CsABF2 activates genes involved in chlorophyll degradation, the phenylpropanoid pathway, and anthocyanin biosynthesis. (A–D) AsODN-mediated knockdown of CsABF2 in tea leaves: (A) relative expression level of CsABF2 following AsODN treatment; (B) transcript levels of chlorophyll degradation–related genes (CsSGR1, CsNYC1, and CsPAO) after CsABF2 silencing; (C) expression changes in phenylpropanoid pathway–related gene CsPALa, CsC4H, and Cs4CL; (D) expression of flavonoid/anthocyanin biosynthetic genes (CsMYB6c, CsF3′H, CsDFR, and CsANS) after CsABF2 suppression. (E–H) Dual-luciferase reporter assays for transcriptional activation by CsABF2: (E) schematic diagram of effector and reporter construct design; (F–H) activation of CsSGR1, CsPALc, and CsMYB6c promoters by CsABF2, shown by increased Luc/Ren ratios in N. benthamiana. (I–N) Regulatory cascade involving CsABF2, CsMYB6c, and CsTT8–2; (I–K) dual-luciferase assays showing activation of CsMYB6c and CsTT8–2 by CsABF2; (L–N) promoter activation of CsF3′H, CsDFR, and CsANS by CsMYB6c, CsTT8-2, and CsWD40, as indicated by Luc/Ren activity. All data are presented as the mean ± SD of three biological replicates (n = 3). Statistical significance was determined by the Student’s t-test, with *P < 0.05 and *P < 0.01.
To elucidate the specific regulatory role of CsABF2 in chlorophyll degradation, phenylpropanoid metabolism, and anthocyanin biosynthesis, the promoter regions of CsSGR1, CsPALa, CsMYB6c, CsF3′H, CsDFR, and CsANS were cloned and inserted into the pGreenII 0800 vector for dual-luciferase reporter assays. The results demonstrated that CsABF2 significantly enhanced the promoter activities of CsSGR1, CsPALa, and CsMYB6c, whereas it had no significant effect on the transcriptional activation of CsF3′H, CsDFR, or CsANS (Fig. 6E–H). These findings suggest that CsABF2 plays a key upstream role in regulating chlorophyll degradation and anthocyanin metabolism, thereby contributing to leaf coloration and metabolic reprogramming under low-temperature conditions. In the flavonoid biosynthetic pathway, R2R3-MYB, bHLH, and WD40 form a MYB-bHLH-WD40 (MBW) complex that serves as a core regulatory module [43, 44]. Co-expression assays involving CsMYB6c, the bHLH transcription factor CsTT8-2, and WD40 showed that the promoter activities of CsF3′H, CsDFR, and CsANS were significantly upregulated by 11.59-fold, 8.43-fold, and 13.16-fold, respectively (Fig. 6I–N). These results indicate that CsABF2 regulates the expression of downstream structural genes CsF3'H, CsDFR, and CsANS by activating CsMYB6, thereby promoting anthocyanin biosynthesis.
Discussion
In early spring, low temperatures in high-altitude regions inhibit chlorophyll biosynthesis while promoting anthocyanin accumulation in tea plants. This results in newly sprouted tender leaves displaying tip yellowing or a uniform yellow or reddish hue across the entire leaf. These symptoms gradually fade as the leaves mature. Green tea made from such fresh leaves typically features a refreshing floral fragrance.
The yellowing observed on young, tender leaves of tea seedlings from each cultivar at the high-altitude BY and BM in Xizang (including the YG and LY tea gardens) differs from the spring leaf symptoms of tea plants grown at high altitudes in the mountainous areas of Anhui. In addition to yellowing at the leaf tips and margins, yellow spots appear on green leaves (Fig. 1E). The upper leaves exhibit uneven yellowing, while the lower leaves exhibit signs of browning. Observations indicate that these yellowing and browning symptoms gradually subside as temperatures rise (Fig. 1F). It is speculated that although the daytime maximum temperatures in these regions exceed the critical threshold for tea bud sprouting (10°C), the nighttime minimum temperatures remain in the low-temperature range above 0°C. This condition renders the newly sprouted tea leaves more vulnerable to cold stress.
To verify this hypothesis, tea plant samples were subjected to controlled temperature treatments: constant 4°C, constant 10°C, and a day/night temperature variation of 10°C / 4°C. Data analysis revealed that the day/night temperature fluctuation had a significant impact on the extent of cold injury and the progression of bud development, with lasting effects on the subsequent bud growth (Figs 1G–I). These findings further confirm that tea plants are particularly sensitive to cold stress during the sprouting stage [13, 45, 46]. Based on this, it is recommended that when introducing tea cultivars to high-latitude or high-altitude regions, priority should be given to those with cold tolerance or a later sprouting period.
This study utilized transcriptome sequencing to investigate the DEGs associated with the response of tea plants to cold stress. The analysis revealed that DEGs from both transcriptome datasets were predominantly enriched in pathways related to chlorophyll metabolism and the biosynthesis of phenylpropanoids and phenolic compounds (Figs 2 and 3). It is worth noting that a cold-induced bZIP transcription factor, CsABF2, was identified as a key candidate potentially involved in regulating leaf yellowing, lignin biosynthesis, and the production of phenolic compounds (Figs 4 and 5).
As a highly conserved protein family in eukaryotes, bZIP transcription factors are composed of a basic DNA-binding domain and a leucine zipper dimerization domain [47]. In the model plant A. thaliana, 75 AtbZIP transcription factors have been classified into 13 subfamilies, designated A to I, as well as S, M, K, and J. Among them, ABF transcription factors belong to subfamily A of the bZIP family [47]. Studies have shown that these transcription factors are crucial in the ABA signaling and are especially important in plant responses to abiotic stresses, including in tea plants [35, 36, 48]. Molecular experiments have demonstrated that ABF2, ABF3, and ABF4 in A. thaliana can directly bind to and activate the promoters of NYE1, PAO, and NYC1, thereby participating in ABA-mediated chlorophyll degradation [34]. In AsODN interference experiments in tea plants, suppression of CsABF2 expression led to a significant decrease in the expression of chlorophyll degradation-related genes CsSGR1 and CsNYC1 (Fig. 6A and B), suggesting that CsSGR1 is likely transcriptionally regulated by CsABF2. Promoter activity assays further indicated that the CsSGR1 promoter is positively regulated by CsABF2 (Fig. 6F). These findings collectively confirm that CsABF2 plays a regulatory role in chlorophyll degradation under low-temperature conditions.
There is increasing evidence that ABFs are involved in regulating the biosynthesis of anthocyanins and other phenolic compounds. In Litchi chinensis Sonn., members of the bZIP family have been shown to play dual roles in pigment metabolism during fruit ripening, LcABF1/2 promote chlorophyll degradation by activating chlorophyll catabolic genes such as PAO and SGR, while LcABF3 influences anthocyanin accumulation by modulating the expression of the anthocyanin-related transcription factor LcMYB1 [49]. In addition, the expression of VvABF2 has been positively correlated with the biosynthesis of stilbenes, secondary metabolites that are crucial for plant defense [50]. AsODN experiments in tea plants further confirmed that CsABF2 not only upregulates the expression of the key lignin biosynthetic gene CsPALa, but also activates the promoter activity of CsMYB6c, a transcription factor regulating anthocyanin biosynthesis (Fig. 6G and H). In summary, these findings demonstrate that CsABF2, which is induced by low temperature, not only facilitates chlorophyll degradation but also promotes lignin and anthocyanin biosynthesis in tea plants. This integrated regulatory mechanism is summarized in a proposed model (Fig. 7).
Schematic diagram of CsABF2 regulation under low-temperature stress.
Materials and methods
Low-temperature treatment of tea seedlings
To assess how climatic conditions at different altitudes affect the growth of tea plants, twenty commercially available one-year-old tea cultivars were planted in the spring of 2022 at three locations in southeastern Xizang: Medog (MD, 1200 m), Zayü (ZY, 1720 m), and the high-altitude Layue in Bayi District (BY, 2600 m) and Yigong (YG,2200 m). After two years of cultivation, the growth and budburst of the seedlings were monitored in the following spring. Leaves exhibiting symptoms of cold damage were collected for transcriptome sequencing.
To simulate the impact of low temperatures in early spring on the growth of tea plants, three day/night temperature regimes were established in an artificial climate chamber: 4°C/4°C, 10°C/4°C, and 10°C/10°C. For each treatment, 90 one-year-old cold-resistant tea plants (Camellia sinensis cv. Zhongbai 1) were used. The seedlings were subjected to the low-temperature treatments for 15 days, followed by 30 days of cultivation under normal conditions at 25°C/20°C. Seedling growth and budburst were recorded after both the low-temperature treatment and the subsequent period at normal temperature.
To investigate the effects of low temperature on gene expression in tea plants, seedlings were exposed to two temperature regimes in the climate chamber: 25°C/20°C and 10°C/4°C (day/night). After 15 days of treatment, fresh leaf samples were collected for transcriptome sequencing and other analyses. Nicotiana benthamiana used in genetic transformation were grown in an artificial climate chamber under a 16-hour light/8-hour dark photoperiod at a constant temperature of 22°C.
Meteorological data in southeastern Xizang
The data (1901–2023) for monthly mean temperature, monthly maximum temperature, and monthly minimum temperature, each at a 1-km spatial resolution, were obtained from the National Tibetan Plateau/Third Pole Environment Data Center (http://data.tpdc.ac.cn) and visualized using ArcMAP10.8.1. To monitor local temperature fluctuations, compact weather stations (model NHQXZ601, Wuhan Zhongke Nenghui Technology Development Co., Ltd.) were installed in tea gardens at MD, ZY, and BY, recording temperature changes from February to June 2024.
Transcriptome and qRT-PCR analysis
Fresh tea leaf samples were submitted to Guangzhou Genedenovo Biotechnology Co., Ltd. for transcriptome sequencing, with three biological replicates per sample. Sequencing was conducted using the Illumina NovaSeq 6000 platform. The resulting paired-end reads were aligned to the reference genome of tea plants (http://tpia.teaplants.cn/) using HISAT2 with default settings. Gene expression was quantified using RSEM and expressed as fragments per kilobase of transcript per million mapped reads. DEGs were identified using the DESeq2 package, with genes meeting the criteria of FDR < 0.05 and |log₂FC| > 1 considered significantly differentially expressed.
NCBI database and Primer 5.0 were used to design qPCR primers for the target gene (Table S7). Total RNA was extracted from tea leaves using the MiniBEST Plant RNA Extraction Kit, following the manufacturer’s instructions. The concentration and quality of the extracted RNA were assessed using a microvolume spectrophotometer and agarose gel electrophoresis, respectively. Reverse transcription was conducted using the PrimeScript™ RT Reagent Kit to synthesize cDNA from the RNA samples. The resulting cDNA was diluted with RNase-free water to a final concentration of 250 ng/μl for use in qRT-PCR analysis. The CsGAPDH gene was used as an internal reference, and the relative expression of the target genes was calculated using the 2^−ΔΔCt^ method.
Gene cloning and promoter cis-acting element prediction
Using cDNA from tea plant samples as the template, the target gene was amplified by PCR with a high-fidelity polymerase. The PCR product was then ligated into a cloning vector, and the inserted sequence was verified by DNA sequencing. The cloned target gene is used for subsequent experiments. To clone and analyze promoter sequences of the genes related to tea plants, chromosomal location for CsABF2, CsF3′H, CsDFR, CsANS, CsSGR1, CsPALa, and CsMYB6c was first retrieved from the published genomes of tea plants. Primers were designed based on the sequence within 2000 bp upstream of the start codon. Genomic DNA was extracted from tea leaves with a plant genomic DNA kit and used as the template for promoter cloning. The amplified promoter fragments were then verified by DNA sequencing. Primers were designed and synthesized based on the full-length coding sequence (Table S7).
The PlantCare website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) was utilized to predict the cis-acting elements of promoters [51]. Subsequently, cis-acting elements associated with transcription factors were screened, and these cis-acting elements were visualized and analyzed using the TBtools software [52].
Phylogenetic tree
A phylogenetic tree of the bZIP family in tea plants was constructed using the neighbor-joining (NJ) method in MEGA7 software [53], with bootstrap analysis performed using 1000 replicates. Gene names were assigned following the nomenclature convention of A. thaliana.
Transient gene silencing in tea plants using antisense oligodeoxynucleotides
AsODN sequences targeting candidate genes were designed using Soligo [54] (https://sfold.wadsworth.org/cgi-bin/index.pl), while sense oligodeoxynucleotide (sODN) sequences were used as controls. The young shoots of tea plants were employed for AsODN treatment. A 100-μM solution of AsODN or sODN was injected into the abaxial side of the third leaf using a syringe. The treated seedlings were then incubated in an artificial climate chamber, and samples were collected after 48 hours for the analysis of gene expression. All primer sequences are listed in Table S7.
Histochemical GUS staining
The promoters of the candidate genes were cloned into the pCAMBIA1304 vector to replace the CaMV35S promoter. The primers used are listed in Table S7. Nicotiana benthamiana leaves were incubated at 37°C for 12 hours, followed by decolorization in 75% ethanol. The ethanol was changed every 3 hours and replaced three times in total. After decolorization, the leaves were collected and photographed. For each treatment, at least three independent transgenic seedlings were analyzed, and representative results are shown.
Dual-luciferase reporter assay
The promoter activation assay was conducted according to the method described by Jiao [44] with a minor modification. Promoter sequences of candidate genes were cloned into the pGreenII 0800 vector, and transcription factor sequences were cloned into the pGreenII 62-SK vector using a one-step rapid cloning kit. After sequence verification, the plasmids were introduced into the Agrobacterium tumefaciens strain GV3101. Equal volumes of Agrobacterium cultures carrying the promoter and transcription factor constructs were mixed and co-infiltrated into N. benthamiana leaves. Three days after infiltration, the leaves were sprayed evenly with D-luciferin sodium salt solution (50 μg/ml) and incubated in the dark for 5 min. Luminescence signals were then detected using a chemiluminescence imaging system. Based on the luminescence results, two leaf discs (10 mm in diameter) were punched from the infected areas and placed in 2-ml centrifuge tubes preloaded with zirconia beads. Samples were flash-frozen in liquid nitrogen and ground at 23 Hz for 60 s. After adding 100 μl of lysis buffer, samples were thawed on ice and lysed for 5 min. The lysates were centrifuged at 13 680 xg for 5 min at 4°C, and the supernatants were collected. Firefly and Renilla luciferase activities were measured according to the manufacturer’s protocol for the dual-luciferase assay kit.
Chlorophyll content determination
The determination of chlorophyll content was conducted according to the previously reported method [55] with modifications. A total of 0.10 g of fresh tea leaves was finely chopped and homogenized, and then 10 ml of 95% ethanol extraction solution was added. The mixture was incubated in darkness at room temperature for 48 h. After extraction, the solution was centrifuged at 13 680 xg for 10 min and the supernatant was collected. The absorbance at 665, 649, and 470 nm was measured using a spectrophotometer, and the contents of chlorophyll a, chlorophyll b, and carotenoids were measured accordingly.
Anthocyanin content determination
According to the previously published method [56, 57] with a minor modification. A total of 0.05 g of freeze-dried powder derived from fresh tea leaves was extracted with 1 ml of 80% methanol, which contained 0.5% hydrochloric acid, utilizing ultrasonic treatment. The extract was centrifuged at 13 680 xg for 10 min, after which the supernatant was collected. The residue underwent four additional extraction cycles, with all supernatants subsequently combined. The absorbance at 530 nm was measured using a spectrophotometer, and the anthocyanin content was calculated accordingly.
Lignin content determination
Tea leaves were embedded and fixed using OCT compound (SAKURA, Japan), then sectioned transversely through the veins into 24-μm slices using a cryostat microtome (Leica CM3050S, Germany). Intact vein cross-sections were selected for staining, slide preparation, and microscopic observation. Each experiment was repeated three times. The transverse sections from both treatment and control groups were stained with phloroglucinol reagent and imaged using a stereomicroscope (ZEISS Axio Zoom.V16, Germany) to observe lignification in the vascular tissues.
The method for determining total lignin content was referenced from previous studies with slight modifications [58]. A total of 0.10 g of fresh tea leaf powder was weighed and extracted with 2 ml of 50% methanol in a water bath at 80°C for 5 h. The extract was centrifuged at 13 680 xg for 2 min, after which the supernatant was discarded and the pellet was collected and freeze-dried. A total of 0.004 g of the dried sample was transferred into a 10-ml stoppered test tube, followed by adding 2 ml of 25% acetyl bromide–glacial acetic acid solution (v/v) and 80 μL of 70% perchloric acid. The mixture was incubated in a 70°C water bath for 30 min. After incubation, 2 ml of 2 mol/L NaOH was added and mixed thoroughly, and the solution was brought to a final volume of 10 ml with glacial acetic acid. The absorbance of the extract was measured at 280 nm using a UV spectrophotometer, with glacial acetic acid as the blank control. Lignin content was quantified using a standard curve generated with commercially available alkali lignin.
ABA extraction and content determination
The content of ABA was determined using a method adapted from existing literature [59]. Briefly, 100 mg of fresh leaf samples were ground into a powder in liquid nitrogen. Subsequently, 1 ml of ethyl acetate was added, and the mixture was ball-milled for 10 min, followed by centrifugation at 4°C for 10 min at 13 680 xg. The supernatant was then transferred to a 1.5-ml centrifuge tube, concentrated into a powder using a vacuum centrifuge, and completely dissolved in 50% methanol in water. The resulting solution was centrifuged again at 13 680 xg for 10 min and determination of ABA content in the supernatant using UPLC-QqQ-MS/MS (Agilent Technologies, Palo Alto, CA, USA). The mobile phases consisted of 0.1% (v/v) formic acid in water (solvent A) and 100% (v/v) methanol (solvent B). The elution gradient was set as follows: 0%–20% B for 6.5 min, followed by a balance period from 6.5 to 8.0 min, 95%–20% B from 8.0 to 8.1 min, and a final balance maintained from 8.1 to 10.0 min. The parent ion and their fragments of ABA was analyzed in negative mode by multiple reaction monitoring.
Statistical analysis
Statistical analyses were conducted using one-way analysis of variance (ANOVA) in IBM SPSS Statistics for Windows, version 22.0 (IBMCorp., Armonk, N.Y., USA). Student’s t-test and one-way ANOVA was used to analyze the significance of differences between the two treatments. Graphs were generated using Origin (Pro), version 2024 (OriginLab Corporation, Northampton, MA, USA) and finalized using Adobe Illustrator 2021 for figure layout and export.
Supplementary Material
Web_Material_uhaf279
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xia E, Tong W, Hou Y. et al. The reference genome of tea plant and resequencing of 81 diverse accessions provide insights into its genome evolution and adaptation. Mol Plant. 2020;13:1013–2632353625 10.1016/j.molp.2020.04.010 · doi ↗ · pubmed ↗
- 2Kong W, Kong X, Xia Z. et al. Genomic analysis of 1,325 Camellia accessions sheds light on agronomic and metabolic traits for tea plant improvement. Nat Genet. 2025;57:997–100740097782 10.1038/s 41588-025-02135-z PMC 11985346 · doi ↗ · pubmed ↗
- 3Wang XC, Feng H, Chang YX. et al. Population sequencing enhances understanding of tea plant evolution. Nat Commun. 2020;11:10.1038/s 41467-020-18228-8PMC 747758332895382 · doi ↗ · pubmed ↗
- 4Sun YJ, Zhou J, Guo JS. Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis. Front Biosci (Landmark ed). 2021;26:1714–2234994184 10.52586/5063 · doi ↗ · pubmed ↗
- 5Wang M, Yang J, Li J. et al. Effects of temperature and light on quality-related metabolites in tea Camellia sinensis (L.) Kuntze leaves. Food Res Int. 2022;161:11188236192914 10.1016/j.foodres.2022.111882 · doi ↗ · pubmed ↗
- 6Hao X, Wang B, Wang L. et al. Comprehensive transcriptome analysis reveals common and specific genes and pathways involved in cold acclimation and cold stress in tea plant leaves. Sci Hortic. 2018;240:354–68
- 7Wang P, Jin S, Chen X. et al. Chromatin accessibility and translational landscapes of tea plants under chilling stress. Hortic Res. 2021;8:10.1038/s 41438-021-00529-8PMC 808771633931606 · doi ↗ · pubmed ↗
- 8Chen Y, Fu B, Xie Y. et al. Integrating extreme temperature metrics into tea land suitability models: a GIS-based assessment in the ecological and cultural tourism circle in Western Hubei, China. Theor Appl Climatol. 2025;156: