Antiviral and anti-inflammatory evaluation of herbal extracts: Implications for the management of calf diarrheal diseases
Xi-Rui Xiang, Eun-Seo Lee, Junho Lee, Su Min Kyung, Han Sang Yoo, Tofazzal Md Rakib, Tofazzal Md Rakib, Tofazzal Md Rakib, Tofazzal Md Rakib

TL;DR
This study explores how herbal extracts can fight viruses and reduce inflammation in calves, potentially offering new treatments for calf diarrhea.
Contribution
The study systematically evaluates antiviral and immunomodulatory effects of three herbal extracts in vitro.
Findings
All three herbal extracts inhibited bovine rotavirus and bovine coronavirus in vitro.
The extracts reduced inflammatory mediator expression in macrophage models.
Extract C modulated T helper cell responses and suppressed regulatory T-cell transcription in bovine PBMCs.
Abstract
Traditional herbal extracts are attracting attention in the context of animal disease control because of their low side effects, diverse bioactive compounds, and low antimicrobial resistance risk. However, the underlying mechanisms remain inadequately understood. To characterize the multifaceted biological activities that underlie their therapeutic potential, this study systematically evaluated the antiviral and complex immunomodulatory properties of three distinct herbal combinations (designated Extracts A, B, and C) in vitro. The antiviral activities of the extracts were tested against bovine rotavirus and bovine coronavirus, two major pathogens of neonatal calf diarrhea, and their regulation of inflammatory mediators was assessed in a murine macrophage model (RAW 264.7 cells) stimulated with lipopolysaccharide from Escherichia coli by measuring nitric oxide production and the gene…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
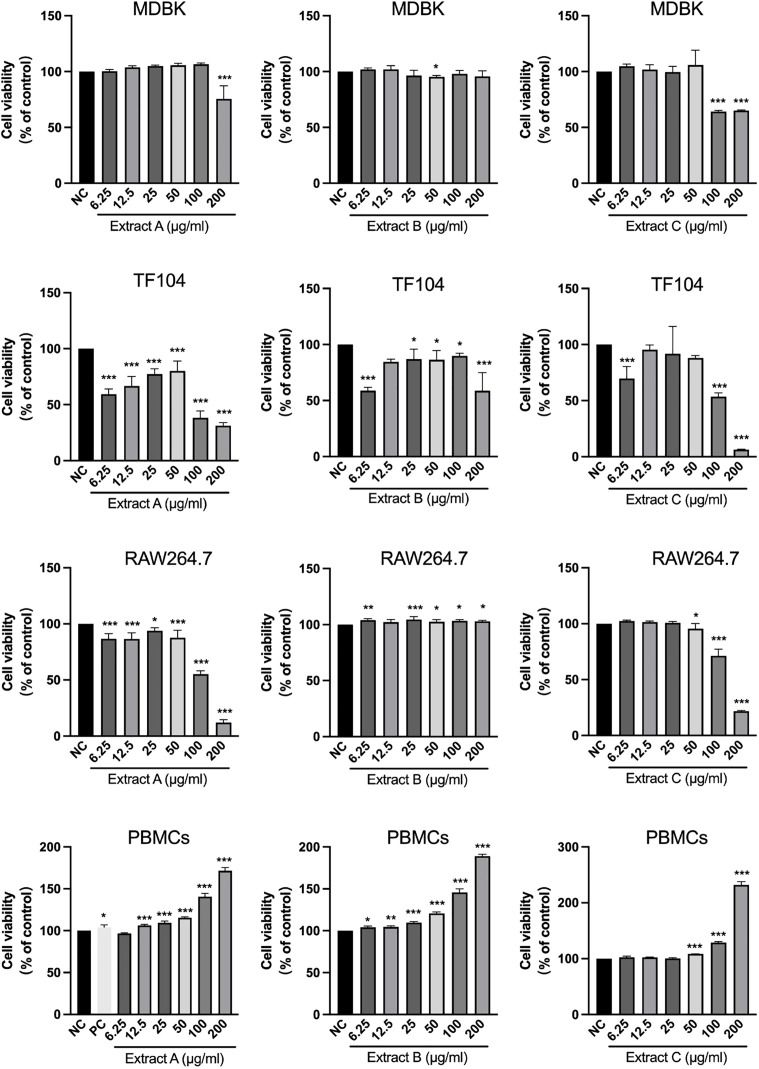 Fig 1
Fig 1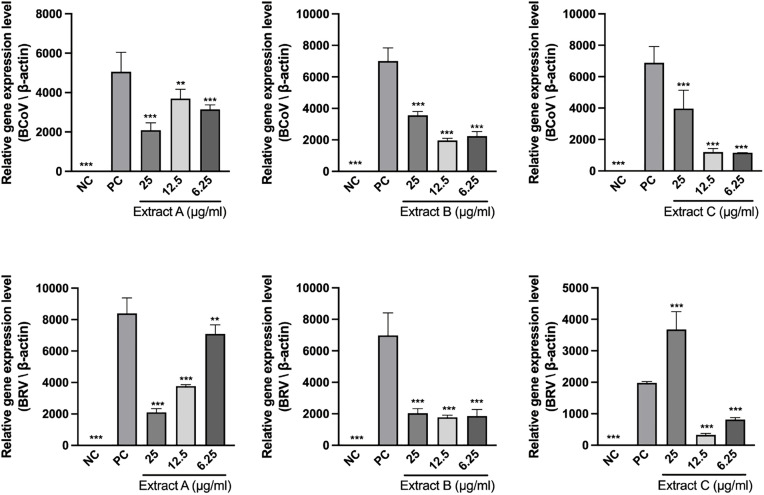 Fig 2
Fig 2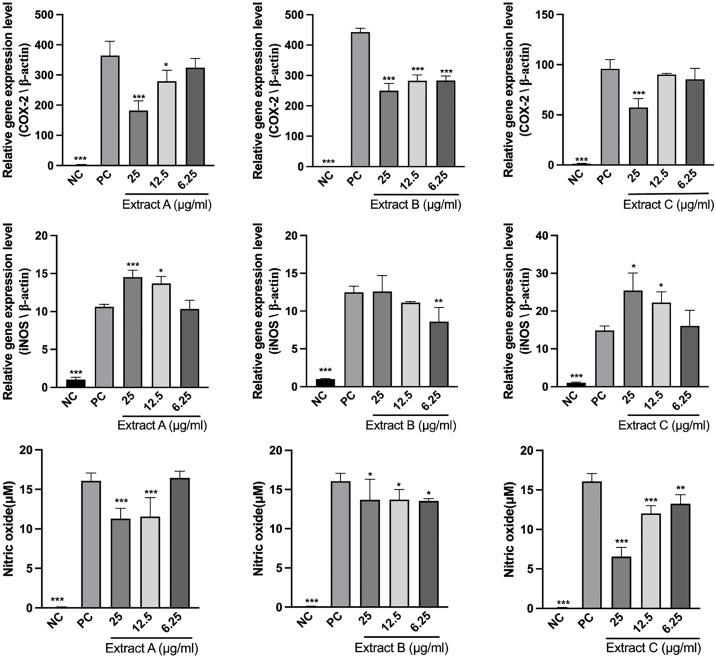 Fig 3
Fig 3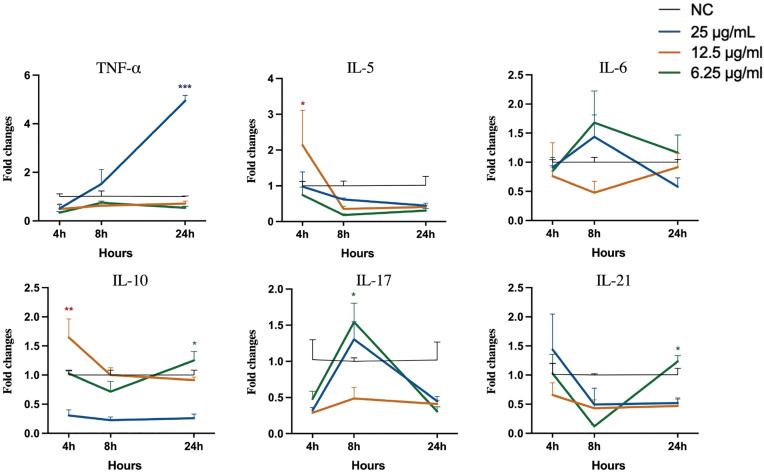 Fig 4
Fig 4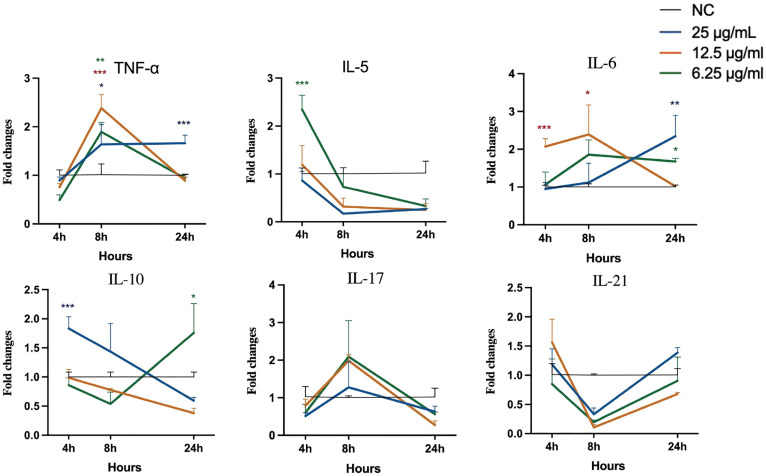 Fig 5
Fig 5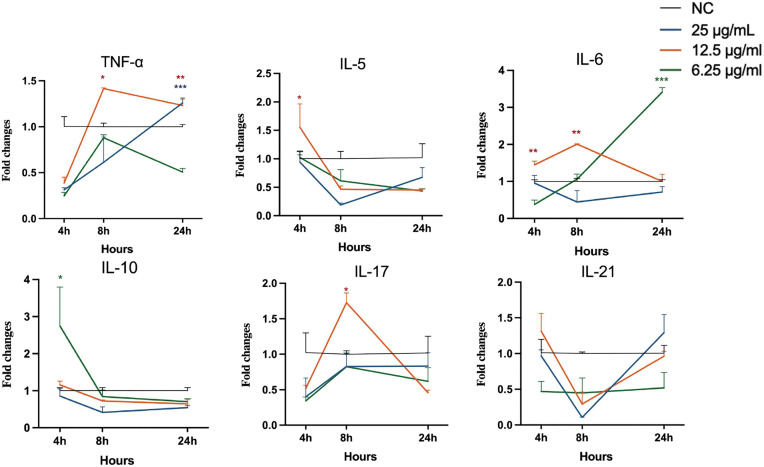 Fig 6
Fig 6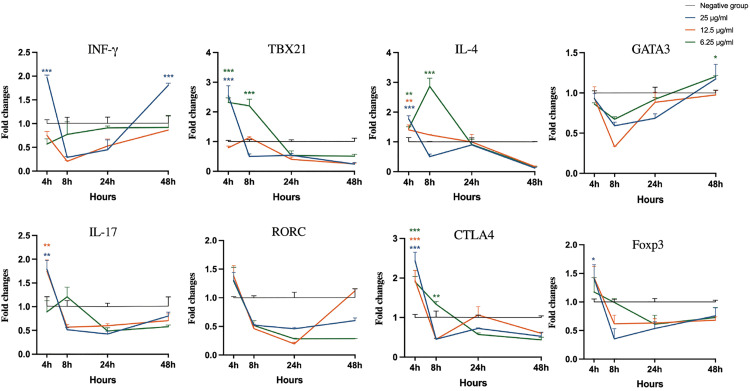 Fig 7
Fig 7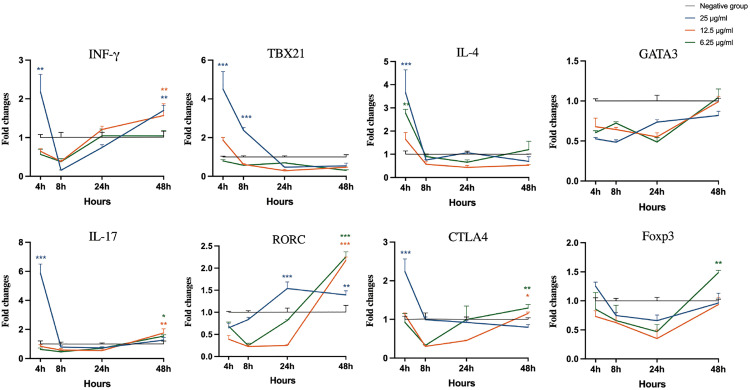 Fig 8
Fig 8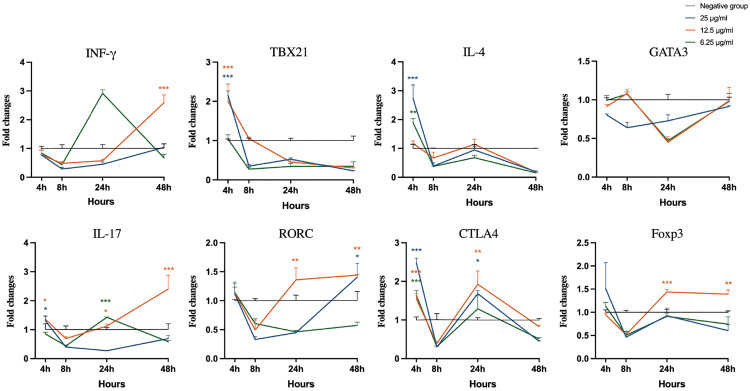 Fig 9
Fig 9- —SMTECH
- —BK21
- —http://dx.doi.org/10.13039/501100020696Research Institute for Veterinary Science, College of Veterinary Medicine, Seoul National University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal health and immunology · Polysaccharides and Plant Cell Walls · Viral gastroenteritis research and epidemiology
Introduction
Diarrhea is among the leading causes of morbidity and mortality in newborn calves, resulting in substantial economic losses [1,2] and posing a serious threat to the cattle industry across Asia and worldwide [3–5]. Among the diverse etiologies of the disease, including viruses, bacteria and parasites [6–8], bovine rotavirus (BRV) and bovine coronavirus (BCoV) are the predominant viral pathogens of enteritis [9,10]. Both viruses severely damage the integrity of the intestinal epithelium, leading to dehydration, electrolyte imbalance and even death [11–13].
At present, clinical treatment strategies for calf diarrhea rely mainly on supportive care [8], and often involve the use of antibiotics to prevent or control secondary bacterial infections [14,15]. However, the widespread use of antibiotics has exacerbated not only the global crisis of bacterial resistance [16], but also complicated subsequent treatments [5,17]. Importantly, the damage caused by viral infection extends beyond direct cytopathic injury to the gut epithelium, and frequently involves dysregulated inflammatory responses [18], such as excessive inflammatory reactions and disruption of immune homeostasis [19]. In antiviral host defense, the immune response driven by T cells plays a vital role [20,21]. CD4+ T helper (Th) cells represent a significant population [20], and include key components such as Th1, Th2, Th17, and regulatory T-cells (Treg) subsets [22]. The balance among these cells is critical, as each produces cytokines and transcription factors that initiate the immune response [23]. Th1 cells promote cellular immunity and antiviral responses through the secretion of cytokines such as interferon-gamma (IFN-γ), interleukin-2 (IL-2), and tumor necrosis factor-alpha (TNF-α), which are regulated by the transcription factor T-box 21 (TBX21, or T-bet) [24], whereas Th2 cells facilitate humoral immunity through cytokines, including IL-4, IL-5, IL-10 and IL-13, under the control of GATA binding protein 3 (GATA3) [25,26]. Th17 cells, characterized primarily by IL-17 and IL-22, are driven by the transcription factor RAR-related orphan receptor C (RORC), and are involved in mucosal defense and inflammatory pathology [27], whereas Treg cells produce cytokines such as IL-10 and transforming growth factor-beta (TGF-β) to suppress excessive immune activation, with high expression of cytotoxic T-lymphocyte antigen 4 (CTLA-4) and the transcription factor Forkhead Box P3 (Foxp3), to prevent tissue damage [28].
Because viral diarrhea involves both viral infection and dysregulated inflammatory responses, agents that can influence both processes are of particular interest. This concept provides the rationale for evaluating the antiviral and immunomodulatory activities of compound herbal extracts. Natural herbs have attracted increasing attention because of their properties involving multiple components and targets [29,30], allowing them to modulate both viral activity and host immunity. Moreover, herbal materials exert relatively low selective pressure for antimicrobial resistance, a feature consistent with One Health goals of mitigating drug resistance. For example, Scutellaria baicalensis [31–33], Commiphora myrrha [34,35] and Nypa fruticans [36,37] are known for their potent anti-inflammatory effects. Boswellia serrata [38–40] and Gardenia jasminoides [41]has been confirmed to have broad antiviral activity. Propolis, on the other hand, has significant immunomodulatory functions [42–44]. Based on these properties, three compound herbal extract formulations (Extracts A, B, and C) were prepared and standardized for evaluation.
To characterize their multifaceted biological activities, this study systematically evaluated the antiviral and immunomodulatory properties of these three herbal extracts in vitro using viral infection models and immune cell assays. The objective was to provide foundational evidence supporting further investigation of these formulations in the context of viral infections associated with calf diarrhea.
Materials and methods
All experiments were carried out in the laboratory facilities of the Research Institute for Veterinary Science, Seoul National University, and were performed by the authors of this study. All animal-related procedures were approved by the IACUC of Seoul National University (SNU-240219-4-1).
Herbal materials
Extract A, Extract B, and Extract C used in this study were provided by K Pharms Co., Ltd. (Suwon, Korea; formerly Yeskin Co., Ltd.). The compositions were as follows: Extract A was composed of B. serrata, C. myrrha and propolis, and Extract B was a single extract of N. fruticans. Extract C was a compound of B. serrata, C. myrrha, S. baicalensis, G. jasminoides, and propolis. These extracts were selected based on their demonstrated bioactivities in gastrointestinal or inflammatory conditions [31,36,37], and are formulated for veterinary health support. According to the information provided by the manufacturer, these extracts were produced using standard procedures for solvent extraction, although the detailed steps of the extraction process are proprietary and not publicly disclosed. All the extracts were dissolved in DMSO to prepare 200 mg/mL stock solutions, which were stored at −20 °C in the dark. Working concentrations of DMSO were freshly prepared immediately before experimentation. The final DMSO concentration did not exceed 0.1% (v/v), and vehicle controls were included accordingly.
Cells and virus culture
Cell lines and virus.
Madin-Darby Bovine Kidney (MDBK) cells, TF104 cells (a clonal derivative of MA-104 monkey kidney cells), and RAW 264.7 mouse macrophages, all of which were maintained as laboratory stocks, were used in this study. Cells were used at passages 5–10 after thawing. All cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco) and 100 µg/mL streptomycin and 100 U/mL penicillin (Gibco), and incubated at 37 °C and 5% CO₂. BCoV (strain number: VR1500040) was obtained from the Animal and Plant Quarantine Agency (Gimchen, Korea), and BRV was isolated from lyophilized commercial vaccines provided by Choong Ang Vaccine Laboratories Co., Ltd. (Daejeon, Korea). BCoV was propagated in MDBK cells, and BRV was propagated in TF104 cells. Infectious titers (50% tissue culture infectious dose, TCID₅₀) were determined by the Reed–Muench method [45], yielding titers of 1 × 10^5.62^ TCID₅₀/mL for BCoV and 1 × 10^2.86^ TCID₅₀/mL for BRV.
Isolation and culture of bovine PBMCs.
Heparinized jugular venous blood was collected from healthy cattle maintained at an experimental farm of Seoul National University. Bovine peripheral blood mononuclear cells (PBMCs) were isolated by centrifugation at 800 × g for 15 min through Leucosep tubes (Greiner Bio-One, Austria). The PBMC layer was harvested, washed with Dulbecco’s phosphate-buffered saline (DPBS; Gibco), and treated with red blood cell lysis buffer (Roche Diagnostics, Germany). The cells were re-suspended in RPMI 1640 medium (Gibco), supplemented with 10% heat-inactivated FBS and penicillin–streptomycin (100 U/mL; 100 μg/mL), and then cultivated at 37°C with 5% CO₂ until use.
Cytotoxicity assay
MDBK, TF104, RAW 264.7 cells and bovine PBMCs were seeded in 96-well plates at a density of 1–4 × 10^4^ cells/well, respectively. The plates were incubated overnight at 37°C to allow for cell attachment. For all experimental conditions, including the negative control (NC), positive control (PC), and extract-treated groups, assays were performed in quadruplicate wells. Subsequently, the culture medium was replaced with fresh medium containing each extract at concentrations of 6.25, 12.5, 25, 50, 100 and 200 μg/mL. The NC group was treated with 0.1% DMSO. For bovine PBMCs, 1 μg/mL lipopolysaccharide (LPS) from Escherichia coli (Sigma-Aldrich, USA) was added to the PC group, which was included as a positive reference for subsequent immunoactivity assays. In accordance with the cell growth characteristics, the viral host cells MDBK and TF104 were treated for 48 h and 96 h, respectively, to determine the optimal time points for evaluating the viral CPE. The immune cells, RAW 264.7 cells and bovine PBMCs, were treated for 24 h and 48 h, respectively, representing the optimal duration for assessing their immunomodulatory responses while maintaining good cell viability.
Adherent cells were processed with MTT reagent (5 mg/mL stock; Thermo Fisher Scientific, USA) for 4 h according to the manufacturer’s instructions. PBMCs received 10 μL of WST-1 (Abcam, UK) per well. The absorbance was measured on a microplate reader at 570 nm for MTT and 450 nm for WST-1.
RNA extraction and reverse transcription
Total RNA was extracted using TRIzol Reagent (Thermo Fisher Scientific). After chloroform phase separation, the RNA was precipitated with isopropanol, washed with 75% ethanol, and finally resuspended in DEPC-treated water. cDNA was synthesized from total RNA using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, USA) following the manufacturer’s protocol and then stored at −20 °C until use.
Antiviral effect on BCoV and BRV
MDBK (2 × 10^4^ cells/well) and TF104 cells (2 × 10^4^ cells/well) were seeded in 6-well plates overnight and inoculated with either BCoV (MOI = 0.1) or BRV (MOI = 0.01) for 1 h. A negative control (NC; vehicle-matched 0.1% DMSO) and a virus-only positive control (PC) were included. After adsorption, the inoculum was replaced with medium containing Extracts A–C at 6.25, 12.5 or 25 μg/mL. The cultures were continued for 48 h in BCoV or 96 h in BRV until CPE appeared. Afterward, the cells were harvested for RNA extraction and subsequent RT-qPCR analysis.
Relative viral gene expression was quantified by RT-qPCR targeting the BCoV M gene and the BRV VP1 gene on a Rotor-Gene Q (QIAGEN, Germany). Reactions contained 10 μL of EzAmp™ qPCR 2 × Master Mix (SYBR Green; ELPIS-BIOTECH, South Korea), 2 μL of cDNA template, and 10 pmol of each primer. Cycling: 95 °C for 3 min; 35 cycles of 95 °C for 15 s and 55–60°C for 55 s. The primers that were designed in this study were generated using Primer-BLAST (NCBI) with default parameters and targeting coding sequences obtained from GenBank. Primer specificity was confirmed by in silico BLAST analysis and by melting curves exhibiting a single peak in qPCR. And then synthesized by Macrogen Inc. (Seoul, Korea), and the sequences are shown in Table 1. The expression was normalized to that of β-actin and analyzed using the 2^−ΔΔCt^ method [46]. The antiviral experiments were independently repeated three times.
Table 1: Sequences of primers used for bovine coronavirus and bovine rotavirus viral RNA by RT-qPCR.
Regulation of Inflammatory Mediators in RAW264.7 Cells
RAW 264.7 cells (1 × 10^4^ cells/well; 96-well plates) were used to evaluate the regulation of inflammatory mediators. Prior to treatment, the cells were incubated overnight to allow for adherence. In an extract-only check, cells treated with Extracts A–C (6.25–25 μg/mL; vehicle 0.1% DMSO) for 24 h showed no increase in nitric oxide (NO) by the Griess reagent system (Promega, USA) at 540 nm (data not shown). For the main assay, cells were pretreated with different concentrations (6.25, 12.5 and 25 μg/mL) of the Extracts A–C for 4 h, and then coincubated with LPS-E.coli at 1 μg/mL for 24 h. A negative control (NC; vehicle only, no LPS) and a positive control (PC; LPS only) were also included. The cell supernatant was collected to quantify production of nitric oxide (NO) by the Griess Reagent System. The cell pellets were harvested for total RNA extraction. The mRNA expression levels of two representative proinflammatory genes, inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), were measured by RT-qPCR with β-actin as the housekeeping gene, according to the method described above. The primers used are listed in Table 2. The experiments were independently repeated three times.
Table 2: Sequences of primer for the detection of inflammation-related genes in RAW 264.7 cells by RT‒qPCR.
Immunomodulatory effects on RAW 264.7 cells and bovine PBMCs
RAW 264.7 macrophages and bovine PBMCs were seeded in 6-well plates at 5 × 10^5^ and 2–3 × 10^5^ cells/well, respectively, and incubated overnight to allow for cell attachment. The cells were treated with different concentrations (6.25, 12.5 and 25 μg/mL) of extracts, LPS-E. coli (1 μg/mL, positive control), or 0.1%DMSO (negative control) for 4, 8 and 24 h in RAW 264.7 cells or 4, 8, 24 and 48 h in bovine PBMCs. At each time point, total RNA was extracted, and RT‒qPCR was performed as previously described, while “relative” fold changes in cytokine expression levels were compared with those in the negative control group by the 2^−ΔΔCt^ method. The targets included immunomodulatory cytokines and transcription factors, as listed in Table 3. The experiments were independently repeated three times.
Table 3: Primer sequences used for the quantification of gene in RAW264.7 cells and bovine peripheral blood mononuclear cells by RT-qPCR.
Statistical analysis
All experiments were repeated a minimum of three times, and the data are presented as the mean ± standard deviation (SD). The information was analyzed using IBM SPSS Statistics 26, and figures were generated with GraphPad Prism 10.0.0. Comparisons among multiple groups were conducted using one-way analysis of variance (ANOVA), followed by Dunnett’s post-hoc test for pairwise comparisons between each extract-treated group and the corresponding control group. A p-value of less than 0.05 was considered to indicate statistical significance (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Cytotoxicity across four cell types
To systematically evaluate cellular safety, four cell types were treated with Extracts A–C, as shown in Fig 1, and their viability is expressed relative to that of the vehicle control (100%). In viral host MDBK cells, only Extract A at 200 μg/mL and Extract C at 100 and 200 μg/mL significantly reduced cell viability to 75.58%, 64.24% and 65.23%, respectively (p < 0.001), whereas the other concentrations did not show obvious cytotoxicity. Treatment of TF104 cells for 96 h resulted in a dose-dependent loss of viability, with 100–200 µg/mL reducing viability to ≤50%. In immune cells, 200 µg/mL Extracts A and C decreased the viability of RAW 264.7 cells to 12.05% and 21.72%, respectively (p < 0.001). In contrast, after treatment with bovine PBMCs for 48 h, the three extracts exhibited no cytotoxicity, and the high concentration treatment significantly increased cell growth. At 200 μg/mL, Extract C increased the cell viability to 231.97% of that of the control group (p < 0.001). Similarly, the PC group showed no significant cytotoxicity to bovine PBMCs. Therefore, based on these findings, noncytotoxic concentrations of 6.25, 12.5, and 25 µg/mL for Extracts A, B, and C, respectively, were selected for all subsequent experiments.
*The extracts have differential cytotoxic effects on different cell types.MDBK (48 h), TF104 (96 h), RAW264.7 (24 h), and bovine PBMCs (48 h) cells were treated with graded concentrations (6.25–200 μg/mL) of Extract A, B or C. The NC group was treated with vehicle (0.1% DMSO), and the PC group was treated with 1 μg/mL LPS. The data are presented as the mean ± SD (n = 4) and are expressed as a percentage of the NC group. Statistical significance relative to the NC group was determined by one-way ANOVA with Dunnett’s post-hoc test (*p < 0.05, **p < 0.01, **p < 0.001).
Antiviral effects on BCoV and BRV
As shown in Fig 2, three concentrations (6.25, 12.5, and 25 μg/mL) of all the extracts significantly inhibited BCoV. Extract A exhibited the strongest suppression at 25 µg/mL (p < 0.001), whereas Extracts B and C showed greater inhibition at 6.25–12.5 µg/mL than at 25 µg/mL. The extracts presented distinct patterns of activity against BRV. The effect of Extract A was strongly dose dependent, with greater inhibition observed at higher doses. At all tested concentrations, Extract B reduced viral replication. However, Extract C showed a biphasic pattern, with an inhibitory effect at the low concentration and a significant increase at the highest concentration of 25 μg/mL (p < 0.001).
*Antiviral effects of the extracts on BCoV and BRV.MDBK and TF104 cells were infected with BCoV (MOI = 0.1) and BRV (MOI = 0.01), respectively, and then treated with graded concentrations (6.25–25 μg/mL) of each extract for 48 h or 96 h. The relative mRNA expression of the BCoV M gene and the BRV VP1 gene was quantified by RT-qPCR. NC represents uninfected cells. Data are presented as the mean ± SD (n = 3), and statistical significance relative to the virus control (PC; treated with 0.1% DMSO) was determined by one-way ANOVA with Dunnett’s post-hoc test (**p < 0.01, **p < 0.001).
Herbal extracts reduce NO and COX-2 but differentially modulate iNOS
The regulation of inflammatory mediators by the extracts were evaluated in LPS-stimulated RAW 264.7 cells (Fig 3). At the transcript level, COX-2 mRNA expression was significantly suppressed by all the extracts. The inhibitory effects of Extracts A and C increased with increasing concentration, as Extract B significantly reduced COX-2 expression at all doses (p < 0.001). However, with respect to iNOS expression, the results revealed that compared with the positive control, Extract B significantly reduced iNOS expression only at 6.25 µg/mL (p < 0.01). Additionally, NO production decreased with all three extracts. It was reduced by Extract C at concentrations ranging from 6.25–25 µg/mL in a dose-dependent manner. At 12.5 and 25 µg/mL, Extract A reduced NO levels (p < 0.001), whereas Extract B did so at all the tested doses (p < 0.05).
*Regulation of inflammatory mediators by the extracts in LPS-stimulated RAW 264.7 macrophages.RAW 264.7 cells were treated with 6.25, 12.5 and 25 μg/mL of each extract for 4 h, followed by stimulation with 1 μg/mL LPS-E. coli for 24 h. The relative mRNA expression levels of COX-2 and iNOS were determined by RT‒qPCR and normalized to that of β-actin. The concentration of nitrite in the cell supernatant was measured using a Griess reagent system. The NC group was left untreated, while the PC group was stimulated with only LPS. The data are presented as the mean ± SD (n = 3). Statistical significance relative to the PC group was determined by one-way ANOVA with Dunnett’s post-hoc test (*p < 0.05, **p < 0.01, **p < 0.001).
Extracts induce differential expression of immunomodulatory genes in RAW 264.7 cells
All three extracts induced dynamic and distinct gene expression profiles over a 24 hours period. As shown in Fig 4, 25 µg/mL Extract A caused significant induction of TNF-α expression at 24 h (p < 0.001). This extract also induced peak expression of IL-6 and IL-17 at 8 h. For Extract B (Fig 5), treatment significantly increased TNF-α expression at 8 hours. In contrast, Extract C (Fig 6) at 6.25 µg/mL significantly induced IL-6 expression at 24 h. In a separate control experiment (S1 Fig), stimulation with LPS-E. coli caused a sustained increase in the expression of most proinflammatory genes, confirming the responsiveness of the assay system.
*Analysis of cytokine gene expression in RAW 264.7 cells stimulated with Extract A.Asterisks (*p < 0.05, **p < 0.01, **p < 0.001) indicate that extract-induced gene expression is significantly different from that in the negative control group at the same time points. Statistical significance was evaluated using one-way ANOVA with Dunnett’s post-hoc test.
*Analysis of cytokine gene expression in RAW 264.7 cells stimulated with Extract B.Asterisks (*p < 0.05, **p < 0.01, **p < 0.001) indicate that extract-induced gene expression is significantly different from that in the negative control group at the same time points. Statistical significance was evaluated using one-way ANOVA with Dunnett’s post-hoc test.
*Analysis of cytokine gene expression in RAW 264.7 cells stimulated with Extract C.Asterisks (*p < 0.05, **p < 0.01, **p < 0.001) indicate that extract-induced gene expression is significantly different from that in the negative control group at the same time points. Statistical significance was evaluated using one-way ANOVA with Dunnett’s post-hoc test.
Expression of T cell-associated genes in bovine PBMCs in response to extracts
As shown in Fig 7, all three concentrations of Extract A caused initial upregulation of CTLA4 at 4 h. The 6.25 µg/mL treatment subsequently increased the TBX21 level at 8 h (p < 0.001), and the 25 µg/mL treatment induced an increase in the IFN-γ level at the final 48-hour time point. Treatment with Extract B at 25 µg/mL resulted in a significant initial upregulation of IFN-γ, TBX21, IL-4, IL-17 and CTLA4 expression (Fig 8; p < 0.01). However, all concentrations of Extract B suppressed GATA3 expression throughout the entire time course. In cells treated with Extract C, 25 µg/mL led to significant increases in TBX21 and IL-4 expression at 4 h, whereas 12.5 µg/mL caused increases in IFN-γ and IL-17 expression at later stages (p < 0.001), with recovery of IL-4 and CTLA4 expression observed at 24 h across all concentrations (Fig 9). After LPS-E. coli stimulation, the mRNA expression of IFN-γ, TBX21, IL-17, and CTLA4 increased, peaking at 24 and 48 h, which established a benchmark for a proinflammatory response (S2 Fig).
*Analysis of cytokine and transcription factor gene expression in bovine peripheral blood mononuclear cells stimulated with Extract A.Statistical significance relative to the negative control at each time point was evaluated using one-way ANOVA with Dunnett’s post-hoc test (*p < 0.05, **p < 0.01, **p < 0.001).
*Analysis of cytokine and transcription factor gene expression in bovine peripheral blood mononuclear cells stimulated with Extract B.Statistical significance relative to the negative control at each time point was evaluated using one-way ANOVA with Dunnett’s post-hoc test (*p < 0.05, **p < 0.01, **p < 0.001).
*Analysis of cytokine and transcription factor gene expression in bovine peripheral blood mononuclear cells stimulated with Extract C.Statistical significance relative to the negative control at each time point was evaluated using one-way ANOVA with Dunnett’s post-hoc test (*p < 0.05, **p < 0.01, **p < 0.001).
Discussion
Neonatal calf diarrhea remains a persistent challenge to the global cattle industry owing to its high incidence rate and the limitations of available therapies [8,9,49]. This multifactorial disease involves both infectious and noninfectious factors, with a high probability of mixed infection by multiple pathogens [50,51]. As potential and valuable medicinal resources with intrinsic multitarget properties, natural herbs are gaining increasing attention in modern veterinary clinical research. [52,53]. Although previous studies have reported the efficacy of certain individual herbs [39,43], the combined antiviral and immunomodulatory effects of combinations of these herbs against BCoV and BRV have not been systematically studied.
In this study, all three herbal extracts clearly inhibited calf diarrhea viruses (BCoV and BRV) at concentrations that did not show detectable cytotoxicity in the cell models used. Notably, Extracts B and C showed strong antiviral effects even at relatively low concentration and the activity of Extract C is consistent with the reported broad-spectrum antiviral properties of its constituents B. serrata and S. baicalensis [38,54]. However, the antiviral effect reversed at higher concentrations, especially for Extract C. Possible mechanisms include a hormetic response [55], and the noncompetitive interactions of herbal compounds with host entry receptors at defined concentrations [56].
The distinct antiviral profiles of Extract A and Extract C (Fig 2) suggest a potential divergence in their mechanisms of action, likely driven by the differential herbal composition of the two formulations. Specifically, Extract A demonstrated efficacy against BCoV, whereas Extract C displayed prominent activity against BRV. These patterns likely stem from the differential susceptibility of each viral pathogen to specific herbal constituents, such as the specific contribution of S. baicalensis and G. jasminoides in Extract C. With respect to enveloped BCoV, inhibitory effects may involve interference with viral entry steps. For example, C.myrrha may mediate the modulation of host membrane mediators in influenza virus [57], and the blockade of β-coronavirus spike glycoprotein interactions as reported for SARS-CoV-2 [58]. Although these interactions at the entry level were not directly tested in this study, their relevance to BCoV inhibition warrants further investigation. With respect to nonenveloped BRV, intracellular pathways have been implicated for baicalin from S. baicalensis via the p-JNK–PDK1–AKT–SIK2 pathway [59]and for genipin from G. jasminoides that exhibits anti-rotavirus activity [41]. Given the complex nature of herb formulations, such interactions may exhibit antagonistic or nonlinear behavior under specific combinations or concentrations. This may explain the anomalous reduction in inhibitory activity observed at high concentrations of Extract C. Similar concentration-dependent antagonism has been observed for other herbal compounds [60], suggesting that rational formulation and dose optimization are essential for future studies.
In the LPS-induced RAW 264.7 macrophage inflammation model, the herbal extracts attenuated inflammatory responses, as reflected by decreased COX-2 expression and suppressed NO production. In particular, the inhibitory effect of Extract C was dose dependent, which is consistent with the report that B. serrata [40] and C. myrrha [35] are potent anti-inflammatory herbs. LPS-E. coli induces an inflammatory response mainly by activating the NF-κB signaling pathway, which also plays a key regulatory role in intestinal inflammatory conditions such as inflammatory bowel disease [61,62]. Studies have indicated that compounds such as S. baicalensis can exert anti-inflammatory effects through the NF-κB pathway, signaling the downregulation of COX-2 expression [63]. Although NF-κB pathway activation was not directly measured in this study, the observed reduction of COX-2 may suggest potential upstream interference with this pathway.
Moreover, the extracts also clearly suppressed NO production, suggesting their potential to block the downstream inflammatory responses. However, their effects on iNOS were more complex. Although iNOS is a direct target of NF-κB [64], its transcription is coregulated by multiple proinflammatory cytokines, including TNF-α, IFN-γ, and IL-6 [62]. Notably, in unstimulated RAW 264.7 cells, the extracts induced TNF-α and IL-6 expression. Therefore, predominant constituents may attempt to downregulate iNOS expression via NF-κB, whereas other components may be counteracted by cytokine induction, suggesting that it may have pathway-selective or multidirectional regulatory effects. Similar discrepancies between iNOS mRNA levels and NO production have been described in research on various natural compounds [65,66]. Although the direct regulatory mechanism of iNOS by the extract remains to be elucidated, the effective inhibition of COX-2 and NO by the extract supports its potential in inflammation-related disease models [67], which warrants further investigation and optimization in the future.
We used two immune cell models, RAW264.7 macrophages and bovine PBMCs, to evaluate the immunomodulatory effects of the herbal extracts in a comparative manner. As a widely used macrophage line [68], RAW264.7 cells provided a baseline for responses associated with macrophages and served as a reference for changes in PBMCs. Even in the absence of external stimulation, the extracts altered the expression of several cytokines, suggesting that certain herbal components may regulate macrophage phenotypes under basal conditions. However, RAW 264.7 cells are not a major source of specific cytokines, such as IL-5 or IL-21. Therefore, only limited changes were observed. In contrast, the PBMC model contains many immune cell types. It showed clearer and more diverse responses [68]. The levels of several cytokines and transcription factors, such as Th1, Th2, Th17, and Treg, changed significantly after extract treatment [24]. These findings suggest that the extracts may have broad immunomodulatory potential in complex immune systems.
As introduced earlier, the dynamic balance immune responses mediated by T helper cells is key to immune regulation. Extract C induced coordinated shifts across makers associated with Th1, Th2, Th17, and Treg cells, with early activation of transcriptional programs related to Th1 and Th2 followed by later enhancement of Th17 markers and concurrent suppression of factors linked to Treg cells. These results suggest that the effects of the extracts can modulate T cell differentiation depending on the treatment conditions. Such multidirectional response is often seen in natural products that work through multiple biological targets. For example, propolis has been reported to activate Th1- and Th2-associated cytokines [69]; B. serrata has been shown to downregulate TNF-α, IL-6, and IL-17, as well as T cell activation markers such as CD4 and CD8 [70]; and reports concerning S. baicalensis have indicated that natural components can alleviate inflammatory responses via the NF-κB and NLRP3 pathways [71]. Collectively, these findings support the potential of compound extracts to coordinate immune responses by modulating representative markers associated with T cell differentiation.
Conclusion
In conclusion, this in vitro study demonstrates that the three tested herbal extracts possess significant antiviral activity against BCoV and BRV, alongside the ability to modulate inflammatory mediators and pathways associated with T helper cells in RAW264.7 and PBMCs. The observed response patterns, which were dependent on concentration and context, highlight the multicomponent and multitarget potential of these formulations. Collectively, these findings provide a mechanistic basis for further investigation of these compound herbal extracts as potential therapeutic agents for neonatal calf diarrhea.
Supporting information
S1 FigAnalysis of cytokine gene expression in RAW 264.7 cells stimulated with a positive control.The positive control contained 1 μg/mL lipopolysaccharide derived from Escherichia coli. Asterisks (**p < 0.01, ***p < 0.001) indicate significant differences compared with the negative control group at the same time points.(TIF)
S2 FigAnalysis of immunomodulatory gene expression in bovine peripheral blood mononuclear cells stimulated with a positive control.The positive control contained 1 μg/mL lipopolysaccharide derived from Escherichia coli. Asterisks (**p < 0.01, ***p < 0.001) indicate significant differences compared with the negative control group at the same time points.(TIF)
S3 TableSupporting materials for figures.(ZIP)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cho Y-I, Yoon K-J. An overview of calf diarrhea - infectious etiology, diagnosis, and intervention. J Vet Sci. 2014;15(1):1–17. doi: 10.4142/jvs.2014.15.1.1 24378583 PMC 3973752 · doi ↗ · pubmed ↗
- 2Uetake K. Newborn calf welfare: a review focusing on mortality rates. Anim Sci J. 2013;84(2):101–5. doi: 10.1111/asj.12019 23384350 PMC 7159576 · doi ↗ · pubmed ↗
- 3Zhu Q, Qi S, Guo D, Li C, Su M, Wang J, et al. A survey of fecal virome and bacterial community of the diarrhea-affected cattle in northeast China reveals novel disease-associated ecological risk factors. m Systems. 2024;9(1):e 0084223. doi: 10.1128/msystems.00842-23 38108282 PMC 10804951 · doi ↗ · pubmed ↗
- 4Heinrichs AJ, Heinrichs BS. A prospective study of calf factors affecting first-lactation and lifetime milk production and age of cows when removed from the herd. J Dairy Sci. 2011;94(1):336–41. doi: 10.3168/jds.2010-3170 21183043 · doi ↗ · pubmed ↗
- 5Ali A, Liaqat S, Tariq H, Abbas S, Arshad M, Li W-J, et al. Neonatal calf diarrhea: A potent reservoir of multi-drug resistant bacteria, environmental contamination and public health hazard in Pakistan. Sci Total Environ. 2021;799:149450. doi: 10.1016/j.scitotenv.2021.149450 34426357 · doi ↗ · pubmed ↗
- 6Yimer M, Gezhagne M, Biruk T, Dinaol B. A review on major bacterial causes of calf diarrhea and its diagnostic method. J Vet Med Anim Health. 2015;7(5):173–85. doi: 10.5897/jvmah 2014.0351 · doi ↗
- 7Wu Q, Li J, Wang W, Zhou J, Wang D, Fan B, et al. Next-generation sequencing reveals four novel viruses associated with calf diarrhea. Viruses. 2021;13(10):1907. doi: 10.3390/v 13101907 34696337 PMC 8537473 · doi ↗ · pubmed ↗
- 8Constable PD. Treatment of calf diarrhea: antimicrobial and ancillary treatments. Vet Clin North Am Food Anim Pract. 2009;25(1):101–20, vi. doi: 10.1016/j.cvfa.2008.10.012 19174285 PMC 7127407 · doi ↗ · pubmed ↗