X-Ray Crystal Structure of the N-Terminal Domain of Staphylococcus Aureus Cell-Cycle Protein GpsB
Nathan I. Nicely, Thomas. M. Bartlett, Richard W. Baker

TL;DR
This paper reports the crystal structure of a key cell-cycle protein in Staphylococcus aureus, revealing its unique shape and role in cell division.
Contribution
The study provides a high-resolution structure of the N-terminal domain of S. aureus GpsB, confirming its unique conformation and role in cell division.
Findings
The N-terminal domain of S. aureus GpsB forms an asymmetric dimer with a bent conformation.
The structure matches one of two conformations previously reported, validating the unique fold of S. aureus GpsB.
The findings support GpsB's role in organizing the cell division machinery in S. aureus.
Abstract
GpsB is a conserved cell-cycle regulator in the Firmicute clade of Gram-positive bacteria that coordinates multiple aspects of envelope biogenesis. Recent studies demonstrate interactions between GpsB and the key division cytoskeleton FtsZ, suggesting that GpsB links cell division to various aspects of cell envelope biogenesis in Staphylococcus aureus and potentially other Firmicutes. We determined a 1.7 Å resolution crystal structure of the N-terminal domain of Staphylococcus aureus GpsB, revealing an asymmetric dimer with a bent conformation. This conformation is nearly identical to one of two conformations reported by Sacco, et al., confirming the unique conformation of S. aureus GpsB compared to other gram-positive bacteria. This structural agreement provides strong validation of the S. aureus GpsB fold and supports its proposed role in organizing the cell division machinery.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
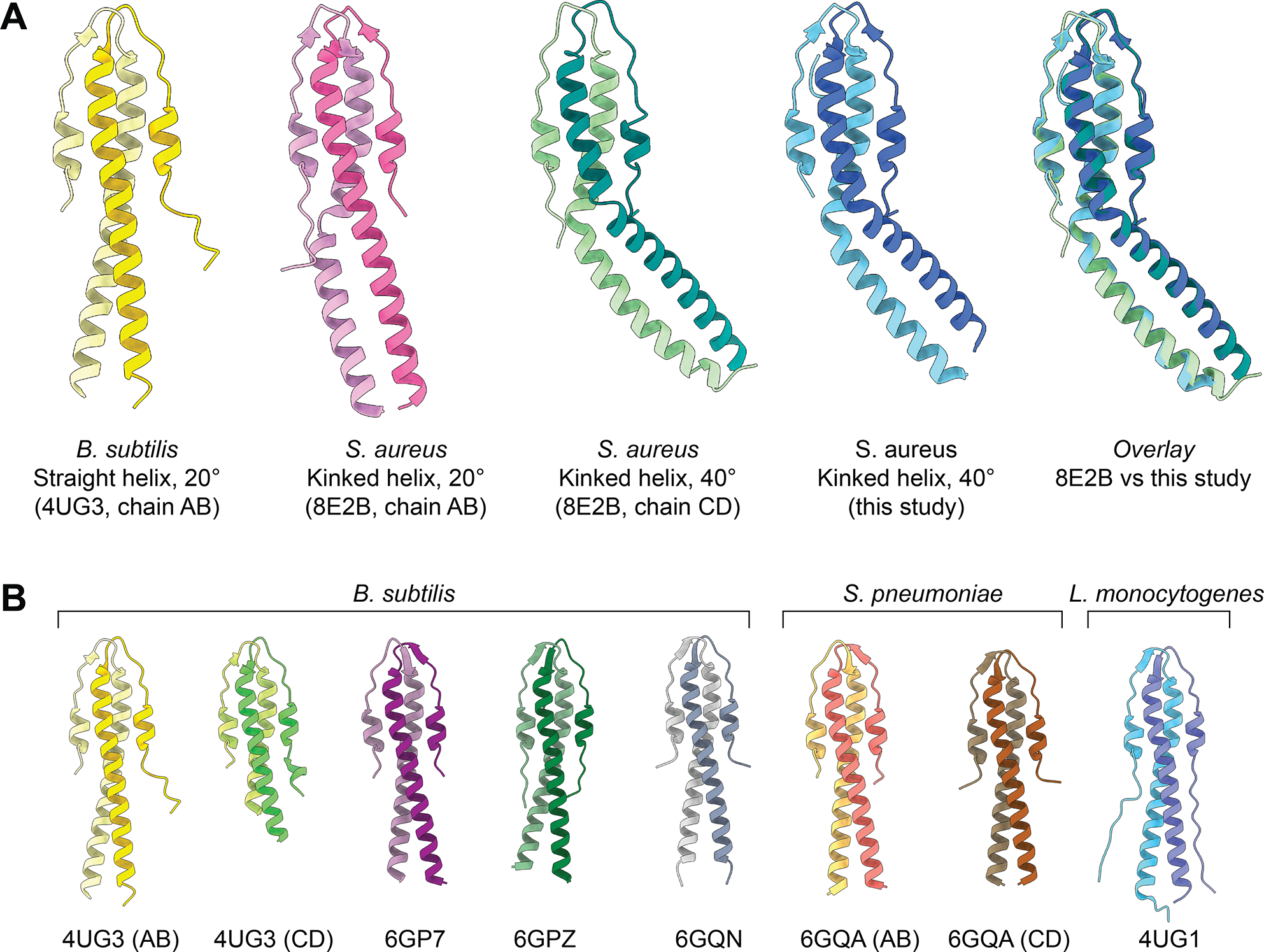 Figure 1
Figure 1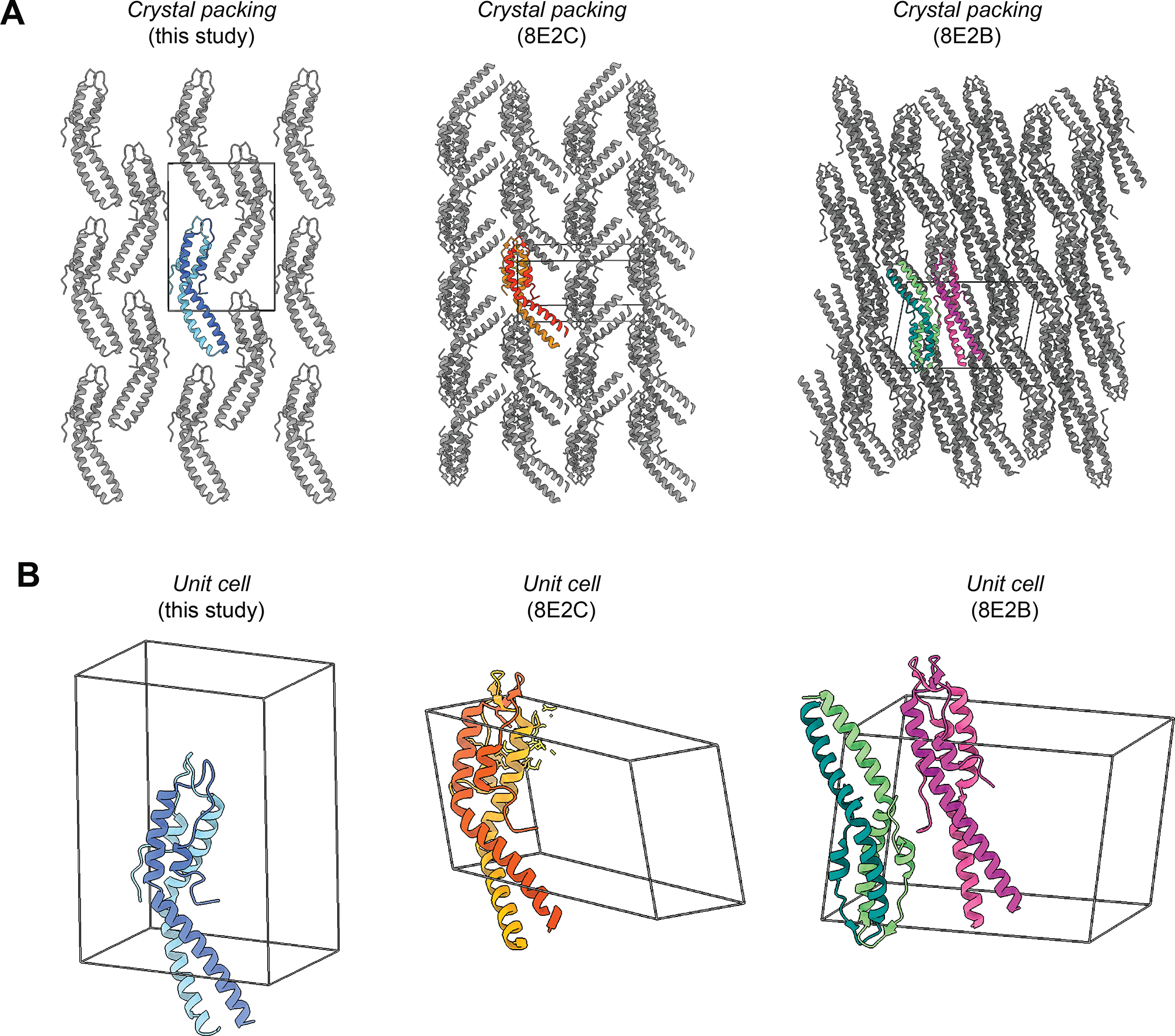 Figure 2
Figure 2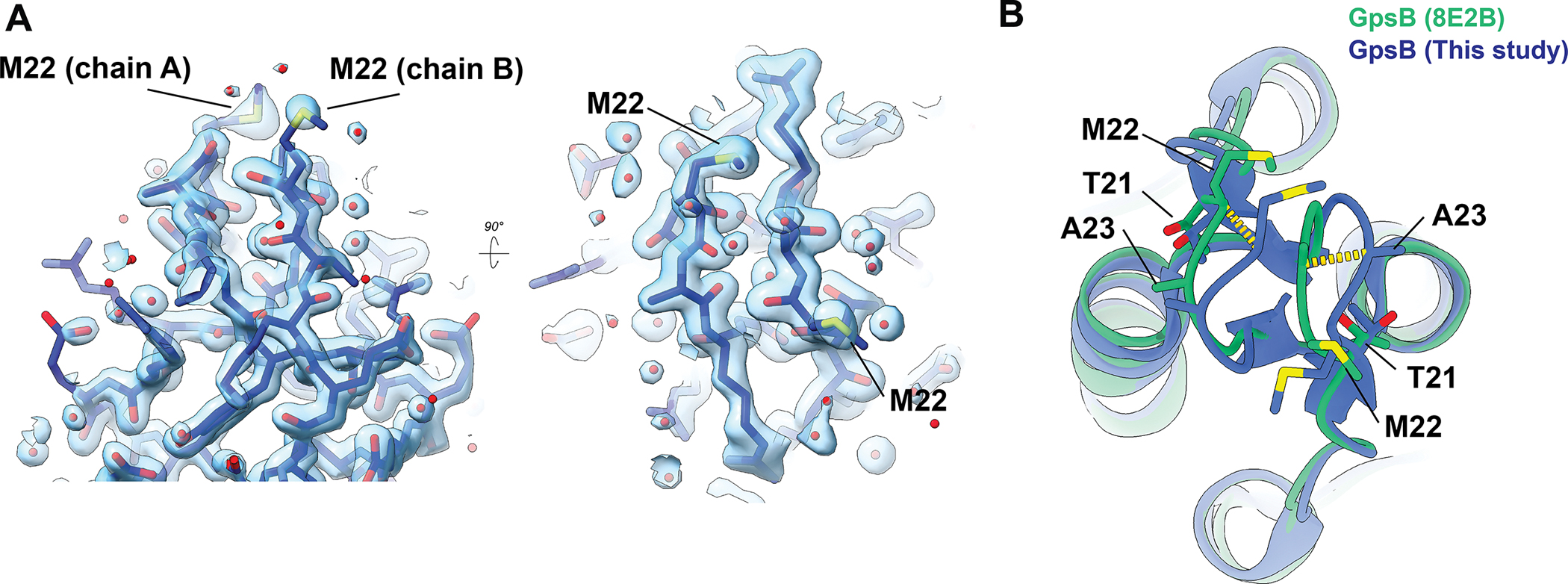 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical and Structural Characterization · Microbial Metabolism and Applications · Antimicrobial Resistance in Staphylococcus
Introduction
GpsB is a widely conserved adaptor protein in Gram‑positive Firmicutes (synonym Bacillota) that coordinates cell‑cycle progression by coupling cell envelope biogenesis to the cell‑division machinery. GpsB has two structured domains — an N-terminal dimerization domain and a C-terminal trimerization domain — which are separated by a flexible linker^1,2^. Thus, GpsB is thought to act as a hexamer that can bind to multiple cell division proteins simultaneously, thereby facilitating the spatiotemporal coordination of various molecular machines^3^. The N-terminal domain of GpsB, in addition to containing a membrane-binding loop that drives binding to the inner leaflet of the membrane^1^, also binds small peptides bearing a consensus (S/T)-R-X-X-R−(R/K) motif. This motif is found in proteins such as PBP1A^4^, FtsZ^5,6^, EzrA^7^, DivIVA^8^, TarO/G^6^, and FacZ^9^, allowing GpsB to coordinate division site localization of multiple protein complexes. Consistent with these binding motifs, GpsB directly binds and regulates FtsZ^5^, contributing to the correct placement of FtsZ and other Staphylococcal division proteins^9^ and coordinating cell envelope growth to division^6^, underlining its intriguing potential to communicate information between cell envelope synthesis and morphogenetic factors. Unsurprisingly, and presumably as a result of its coordination of these features, GpsB is necessary for normal S. aureus morphogenesis^10,11^. GpsB’s interactions with FtsZ^12^ and various cell envelope synthesis factors^4^ are conserved in other Firmicutes, suggesting its role as an adaptor between cell cycle and envelope growth is also broadly conserved.
Prior to 2024, structures for the N-terminal domain of GpsB had been described for Bacillus subtilis^4,13^, Listeria monocytogenes^4^, and Streptococcus pneumoniae^4^. These GpsB orthologs adopt a conserved fold, showing a long parallel two-helix bundle with two short helices that form a 4-helix ‘cap’ at one end^1^. Notably, the central helical bundle is a rigid helix, reinforcing the model of GpsB as a linear adaptor scaffold.
Recently, Sacco et al. revealed a novel conformation of S. aureus GpsB (SaGpsB)^14^, where the N‑terminal homodimer adopts an asymmetric dimer, in which two protomers display a kinked helix conformation, mediated by a hinge formed by a three‑residue insertion exclusive to Staphylococcus species. This hinge comprises a cluster of methionine residues (“MAD” or “MNN” insertion) not found in other Firmicutes, conferring conformational flexibility. Excising this insertion increases thermal stability and abolishes an overexpression lethal phenotype in Bacillus, suggesting functional tuning via flexibility. Thus, functional and structural divergence appears between S. aureus and other Gram‑positives. Whereas GpsB in other species is rigid, SaGpsB seems conformationally dynamic, possibly acting as a regulatory switch in divisome assembly.
We present here a 1.7 Å X-ray crystal structure of the N-terminal helical dimer of the S. aureus GpsB protein. This structure is in strong agreement with one of two conformations observed previously in Sacco et al., providing independent validation of this novel conformation. Notably, our structure agrees with the most structurally divergent conformation compared to the rigid helix conformations of B. subtilis, S. pneumoniae, and L. monocytogenes, highlighting the structural divergence between species.
Results
We independently crystallized the N‑terminal domain (residues 1–75) of SaGpsB and determined its structure at 1.7 Å resolution (Figure 1A, Table 1). Our analysis shows an asymmetric dimer with a kinked helix conformation, in excellent agreement with the GpsB dimers seen in 8E2B.pdb (chains C/D) and 8E2C.pdb (chains A/B) (Figure 1A). Root‑mean‑square deviation (RMSD) between our model and 8E2B chains C/D is ~0.53 Å over all matching Cα atoms (132 residues). RMSD between our model and 8E2C chains A/B is ~0.57 Å over all matching Cα atoms (132 residues). This analysis underscores the similarity of the two structures. While 8E2B contains two SaGpsB dimers in the asymmetric unit, a dimer with a kinked helix of approximately 20° and a dimer with a kinked helix of approximately 40°, our structure best matches the 40° bent helix conformation (Figure 1A). RMSD between our model and the 20° bent helix conformation (8E2B chains A/B) is ~7.56 Å over all matching Cα atoms (132 residues), highlighting the differences between these two structures. However, RMSD is ~0.53 Å when considering only residues 1–47, and ~0.57 Å when considering only residues 48–75, showing that the structural differences between the two complexes can be explained by a rigid motion between the two regions of the protein.
As shown in Figure 1B, all previously reported structures of GpsB from B. subtilis, L. monocytogenes, and S. pneumoniae all display a straight helix conformation. Several of these structures were co-crystallized with a small (S/T)-R-X-X-R−(R/K) motif peptide, although the presence of the peptide does not appear to affect the overall conformation of the helical bundle. Both described SaGpsB conformations vary significantly from the nearly straight helices observed in BsGpsB*, LmGpsB, and Sp*GpsB.
Importantly, our analysis shows a unique crystal packing morphology compared to the other reported structures of SaGpsB, 8E2B and 8E2C (Figure 2). This confirms that the kinked-helix conformation is not an artifact of specific crystal-packing conditions. Our data therefore supports the model proposed by Sacco et al., whereby the hinge-mediated flexibility serves as a dynamic switch in S. aureus. Although no ligand was present in our crystals, the conformation observed is virtually identical to SaGpsB bound to the (S/T)-R-X-X-R−(R/K) motif of PBP4 (8E2C.pdb), suggesting the asymmetric dimer is intrinsic to SaGpsB’s fold and not induced by ligand binding. Thus, this conformation likely represents the physiologically relevant state that mediates binding to partners.
The only significant difference between our structural analysis is slight heterogeneity in the membrane-bending loop (residues ~17–27) (Figure 3), which may reflect dynamic motion of the hinge region, as suggested in Sacco, et al. The high-quality electron density shows the conformation we observe is not a model building artifact, or due to low confidence in map quality at this interface (Figure 3A). Comparing the same region between our structure and the same SaGpsB structure from 8E2B shows a clear loop displacement between the two independent structures, showing that this region of the protein is able to adopt multiple conformations. This heterogeneity is expected for a loop region proposed to insert into the inner leaflet of the membrane^1^. An important caveat is that differences in the loop conformation observed in our structure may be influenced by the crystal packing arrangement. Further analysis of this membrane binding loop is required to understand the functional consequences, if any, of this structural plasticity.
Conclusion
Our independent crystal structure of the SaGpsB N‑terminal domain validates and reinforces the asymmetric, hinge‑mediated conformation first described by Sacco et al. While a rigid helical conformation is a defining features of other bacteria like B. subtilis, S. pneumoniae, and L. monocytogenes, Staph. aureus GpsB has a clear structural plasticity that is an intrinsic feature of the protein. These findings support the hypothesis that hinge flexibility enables regulatory control of divisome component assembly by modulating GpsB interactions with other binding partners. Together, this work affirms GpsB’s role as a dynamic adaptor in S. aureus cell‑division and provides a solid structural foundation for further functional studies.
Materials & Methods
GpsB 1–75 purification
GpsB residues 1–75 (from S. aureus strain NCTC 8325; Uniprot Q2FYI5; SAOUHSC_01462) were recombinantly purified as a fusion with an N-terminal 10xHis-SUMO tag. Briefly, BL21 (DE3) E. coli transformed with the expression plasmid were grown in Lysogeny Broth (LB) with 50 μg/mL kanamycin to mid log phase (OD_600_ ~0.6) and induced with 0.5 mM Isopropyl β-D-1-thiogalactopyranoside (IPTG) at 18° C overnight. Cells were harvested in lysis buffer (20 mM HEPES, pH 7.5; 500 mM NaCl, 20 mM Imidazole, 1 mM DTT, 1 mM PMSF), lysed by sonication, and clarified via centrifugation at 28,000 g. Lysate was applied to Ni^+^-NTA resin (GoldBio), washed with high salt buffer (20 mM HEPES, pH 7.5, 1000 mM NaCl, 20 mM Imidazole), low salt buffer (20 mM HEPES, pH 7.5, 100 mM NaCl, 20 mM Imidazole), and eluted with elution buffer (20 mM HEPES, pH 7.5, 100 mM NaCl, 300 mM Imidazole). Protein was dialyzed overnight into crystallization buffer (20 mM HEPES, pH 7.5, 100 mM NaCl, 1 mM DTT), along with SUMO protease to cleave the purification tag. The following day, the uncleaved protein and free His-SUMO tag were removed by passing the eluent over Ni+-NTA resin. The protein was further purified by anion exchange chromatography using a 5 mL HiTrap Q FF column (Cytiva) in a background buffer of 20 mM HEPES, pH 7.5 using a linear gradient from 100 mM NaCl to 500 mM NaCl over 20 column volumes. Fractions containing GpsB were pooled, concentrated, and applied to a Superdex75 size exclusion chromatography column (Cytiva) in a background buffer of 20 mM HEPES, pH 7.5, 100 mM NaCl. The protein was concentrated to 5 mg/mL.
GpsB 1–75 crystallization, data collection, and model building
Protein was tested for crystallization against common commercially available crystal screens using a Mosquito dropsetter (SPT Labtech) with drops composed of 200 nl protein and 200 nl reservoir solution set over 30 μl reservoir volumes. Crystals were observed within one week over a reservoir solution composed of 1.0 M Succinic Acid, 0.1 M HEPES pH 7.5, 1 %w/v PEG 2000 MME. The crystals were briefly soaked in reservoir supplemented with 15% ethylene glycol then cryocooled in liquid nitrogen. Diffraction data were collected at Southeast Regional Collaborative Access Team (SER-CAT) 22-ID beamline at the Advanced Photon Source, Argonne National Laboratory, using an incident beam of 1 Å in wavelength. Data were reduced in HKL-2000^15^. The structure was phased by molecular replacement using Phaser^16^ with PDB 8e8b as the search model^14^. Real space rebuilding were done in Coot^17^, and reciprocal space refinements and validations were done in PHENIX^18^. Coordinates and structure factors have been deposited in the Protein Data Bank (PDB) with accession number 9PV2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Halbedel S; Lewis RJ Structural Basis for Interaction of Div IVA/Gps B Proteins with Their Ligands. Mol. Microbiol. 2019, 111 (6), 1404–1415.30887576 10.1111/mmi.14244 · doi ↗ · pubmed ↗
- 2Hammond LR; White ML; Eswara PJ ¡v IVA La Div IVA! J. Bacteriol. 2019, 201 (21), e 00245–19.31405912 10.1128/JB.00245-19PMC 6779457 · doi ↗ · pubmed ↗
- 3Cleverley RM, Rismondo J, Lockhart-Cairns MP, Van Bentum PT, Egan AJ, Vollmer W, Halbedel S, Baldock C, Breukink E, Lewis RJ. Subunit Arrangement in Gps B, a Regulator of Cell Wall Biosynthesis. Microb Drug Resist. 2016 Sep;22(6):446–60.27257764 10.1089/mdr.2016.0050 PMC 5111876 · doi ↗ · pubmed ↗
- 4Cleverley RM; Rutter ZJ; Rismondo J; Corona F; Tsui H-CT; Alatawi FA; Daniel RA; Halbedel S; Massidda O; Winkler ME; Lewis RJ The Cell Cycle Regulator Gps B Functions as Cytosolic Adaptor for Multiple Cell Wall Enzymes. Nat. Commun. 2019, 10 (1), 26130651563 10.1038/s 41467-018-08056-2PMC 6335420 · doi ↗ · pubmed ↗
- 5Eswara PJ; Brzozowski RS; Viola MG; Graham G; Spanoudis C; Trebino C; Jha J; Aubee JI; Thompson KM; Camberg JL; Ramamurthi KS An Essential Staphylococcus Aureus Cell Division Protein Directly Regulates Fts Z Dynamics. e Life 2018, 7, e 38856.30277210 10.7554/e Life.38856 PMC 6168285 · doi ↗ · pubmed ↗
- 6Hammond LR; Sacco MD; Khan SJ; Spanoudis C; Hough-Neidig A; Chen Y; Eswara PJ Gps B Coordinates Cell Division and Cell Surface Decoration by Wall Teichoic Acids in Staphylococcus Aureus. Microbiol. Spectr. 2022, 10 (3), e 0141322.35647874 10.1128/spectrum.01413-22PMC 9241681 · doi ↗ · pubmed ↗
- 7Steele VR; Bottomley AL; Garcia-Lara J; Kasturiarachchi J; Foster SJ Multiple Essential Roles for Ezr A in Cell Division of Staphylococcus Aureus. Mol. Microbiol. 2011, 80 (2), 542–555.21401734 10.1111/j.1365-2958.2011.07591.x · doi ↗ · pubmed ↗
- 8Bottomley AL; Liew ATF; Kusuma KD; Peterson E; Seidel L; Foster SJ; Harry EJ Coordination of Chromosome Segregation and Cell Division in Staphylococcus Aureus. Front. Microbiol. 2017, 8, 1575.28878745 10.3389/fmicb.2017.01575 PMC 5572376 · doi ↗ · pubmed ↗