Pisinnocaris subconigera—a valid species of early Cambrian fuxianhuiid
Huijuan Mai, Hong Chen, Ailin Chen, Jin Guo, Xianguang Hou, Yu Liu

TL;DR
This study confirms that Pisinnocaris subconigera is a distinct species of early Cambrian fuxianhuiid, not a juvenile form of another species.
Contribution
The study provides new morphological evidence supporting the taxonomic validity of Pisinnocaris subconigera.
Findings
P. subconigera has four short prothoracic segments with biramous appendages.
It has four opisthothoracic segments with four pairs of appendages and four limbless abdominal segments.
Both small and large individuals show consistent morphology, ruling out a juvenile identity.
Abstract
Pisinnocaris subconigera was first described as a rare, small euarthropod from the early Cambrian Chengjiang biota, southwestern China. The taxonomic validity of this species was later challenged due to the lack of essential morphological details to differentiate it from Fuxianhuia protensa, especially from the juvenile perspective. Here, we examined the holotype and additional specimens of P. subconigera with multiple imaging techniques, such as microscope optical imaging, micro-CT scanning and computer-based 3D rendering, and revealed the previously unknown ventral organization of P. subconigera. New findings include four short prothoracic segments each bearing a pair of biramous appendages, four opisthothoracic segments each with four pairs of appendages, and four limbless abdominal segments. Both the small and large individuals exhibit unique and consistent morphological…
Click any figure to enlarge with its caption.
 Figure 1
Figure 1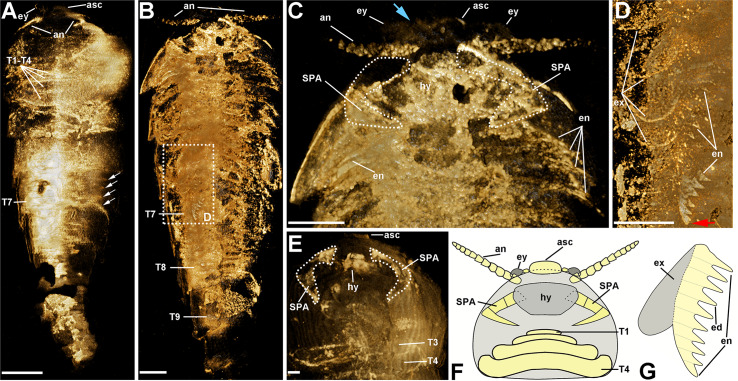 Figure 2
Figure 2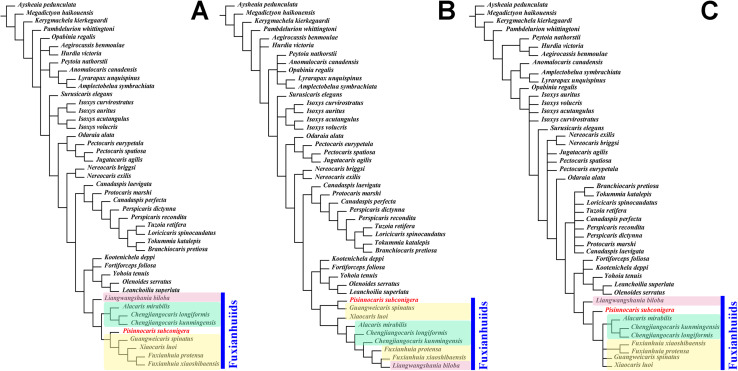 Figure 3
Figure 3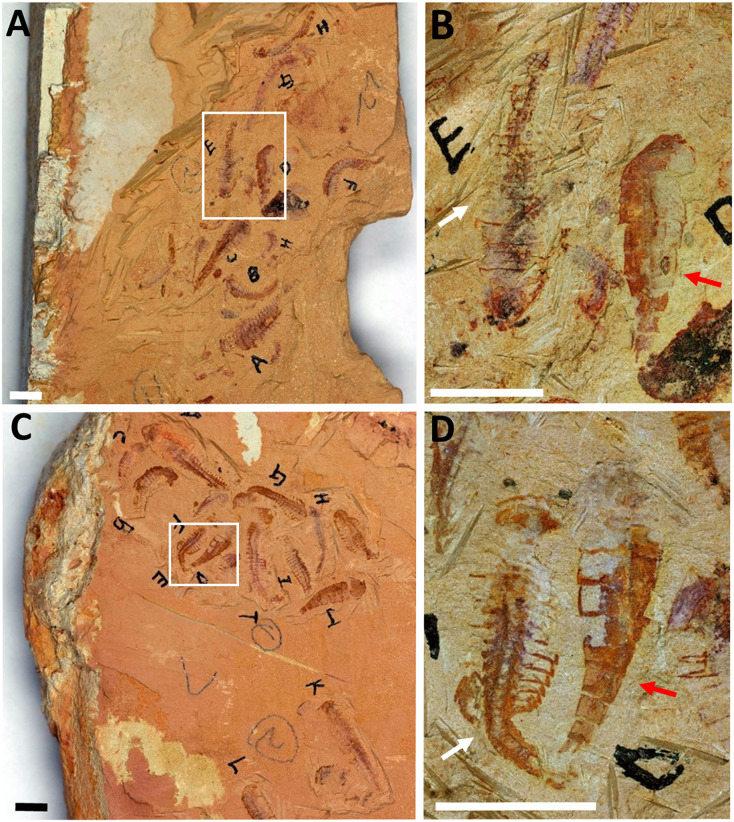 Figure 4
Figure 4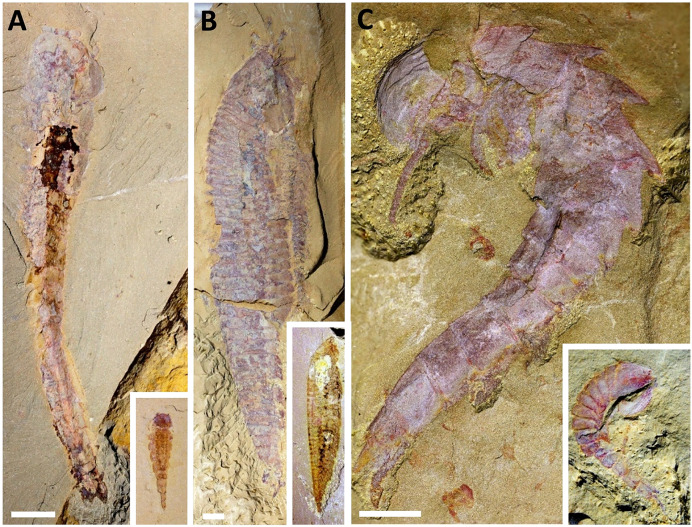 Figure 5
Figure 5- —Natural Science Foundation of Yunnan Province
- —Yunnan Key Laboratory for Palaeobiology, Yunnan University
- —High-Level Talents of Liupanshui Normal University
- —Guizhou Science and Technology Fund
- —Guizhou Provincial Education Department
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPaleontology and Stratigraphy of Fossils · Paleontology and Evolutionary Biology · Geological formations and processes
Introduction
Fuxianhuiids are a group of early Cambrian euarthropods with a unique combination of characters: an eye-bearing anterior sclerite; a pair of antennae; a carapace overlapping a pair of specialized post-antennal appendages (SPAs) and several shortened prothoracic segments each carrying a pair of biramous limbs; opisthothoracic segments bearing more than one pair of biramous limbs and the abdominal segments bearing no limbs (Ortega-Hernández, Yang & Zhang, 2018; Chen et al., 2018). Thus far, all known fuxianhuiids are reported exclusively from the lower Cambrian (Series 2, Stages 3 and 4) in eastern Yunnan, China (Hou, 1987; Hou & Bergström, 1991; Chen, 2005; Luo et al., 2007; Yang et al., 2013, 2018; Chen et al., 2018; Liu et al., 2020a). Despite their limited distribution both in space and time, fuxianhuiids play a role in understanding early arthropod evolution—they have been considered stem-group arthropods based on the relatively simple limb morphology (e.g., Waloszek et al., 2005), or the sister-group to crown Mandibulata based on its head segmentation pattern and the morphology of the SPAs (Aria, Zhao & Zhu, 2021). Taxonomic controversies among some fuxianhuiids have also occurred. These include (1) Shankouia zhenghei (Waloszek et al., 2005) hypothesized to represent a synonym of Liangwangshania biloba (Chen et al., 2018); (2) Xiaocaris has been separated from Jianshania to represent the latest known genus of fuxianhuiid (Liu et al., 2020a); and (3) Pisinnocaris subconigera has been considered a junior synonym of Fuxianhuia protensa (Fu et al., 2018).
Pisinnocaris subconigera, a rare, small euarthropod from the early Cambrian Chengjiang biota, was first reported by Hou & Bergström (1998), but with uncertainties pertaining to the higher taxonomic assignment (i.e., Class, Order and Family) (see Hou & Bergström, 1998). Due to the lack of specimens, P. subconigera has only been briefly mentioned in a few studies after its original description (see Xu, 2004; Zeng et al., 2014). In 2018, Fu et al. (2018) identified new individuals of P. subconigera preserved with specimens representing various ontogenetic stages of Fuxianhuia protensa (Fu et al., 2018). However, we hypothesize that those new individuals of P. subconigera are misinterpreted as juveniles of F. protensa, which then leads to questions about the taxonomic validity of P. subconigera (Fu et al., 2018).
Here, we restudy the holotype specimen of P. subconigera together with several new ones collected in recent years (Figs. 1 and S1). Multiple imaging techniques, such as microscope optical imaging, micro-CT scanning and computer-based 3D rendering help reveal new morphologies from the ventral surface of the animal, which provides new evidence to clarify the taxonomic validity and the phylogenetic position of this species.
Dorsal morphology of Pisinnocaris subconigera.(A) Holotype, NIGPAS 115417a, with four opisthothoracic tergites (T5–T8) and one of the paired leaf-like tail flukes. (B) CJHDM00070, lack of carapace reveals four reduced/shortened prothoracic (T1–T4) tergites underneath. The rest of the body consists of four opisthothoracic (T5–T8) and fourabdominal tergites (T9–T12), and a pair of leaf-shaped tail flukes. (C) YKLP 17301, displays four opisthothoracic tergites (T5–T8). (D) CJHMD00066a, the largest specimen found so far, showing four opisthothoracic tergites (T5–T8) and a pair of leaf-like tail flukes. Arrows indicate keels. Red triangle indicates folds on edges of the carapace. Abbreviations: an, antenna; asc, anterior sclerite; ca, carapace; ey, eye; hy, hypostome; Tn, trunk segment n; tl, tail flukes. Scale bars: 1 mm.
Materials and Methods
Materials
Specimens used in this study include fourteen nearly complete fossils of P. subconigera, two slabs with P. subconigera and F. protensa preserved together, one fossil of each Chengjiangocaris longiformis, Liangwangshania biloba and Guangweicaris spinatus and three small fossils of unidentified species. These specimens were collected from Chengjiang, Yunnan Province, China, and are deposited either at Yunnan Key Laboratory for Palaeobiology, Yuxi Normal University, or Chengjiang Fossil Museum. Details of the specimens can be found in Table S1. All studied fossils are preserved in yellowish mudstones from the Yu’anshan Member of the Chiungchussu Formation, Cambrian (Series 2, Stage 3), the main localities are shown in Fig. S2 (see also Hou et al., 2017). The specimens are partially pyritized which allows enough signal revealed with micro-computed tomography (micro-CT) scanning (Gabbott et al., 2004).
Methods
Optical photographs of specimens shown in all figures were captured using a Canon EOS 5DSR with an MP-E 100 mm objective lens.
Micro-computed tomography (Micro-CT)
X-ray tomography was used to scan three specimens with a Zeiss Xradia Versa 520 micro-CT scanner at the Yunnan Key Laboratory for Palaeobiology, Yunnan University, China. Scanning parameters are as the follows: NIGPAS 115417a (Holotype): beam strength: 50 kV/4w, no filter, pixel size: 3.65 µm, number of TIFF images: 2,799; YKLP 17301: beam strength: 50kV/4w, no filter, pixel size: 6.31 µm, number of TIFF images: 2,435; CJHMD00066a: beam strength: 80 kV/7w, no filter, pixel size: 16.54 µm, number of TIFF images: 3,511. Volumetric reconstructions of specimens from tomographic slices were performed in Drishti software (v 2.4) as described before (Zhang et al., 2022).
Phylogenetic analyses
Our phylogenetic analyses are based on a previously published character matrix for annelids and their close relatives (Yang et al., 2018), which has been updated with the addition of new taxa and fossil data (Chen, 2020). Analyses are performed with TNT (Maximum Parsimony) and MrBayes (Bayesian Inference) (Ronquist et al., 2012; Du et al., 2019). Parsimony analysis was undertaken with the software TNT (version 1.5), where the traditional search, implied weights and equal weights were used. Other settings included a random seed of 1, k = 3 and a strict consensus rule. Bayesian analysis was conducted in MrBayes (version 3.2.7), use the same matrix but removed six consecutive measurement data (characters 0 to 5, which is non-classified data), with a run of 20.000.000 Markov Chain Monte Carlo generations containing four Markov chains under the MKV+ gamma model. In each run, trees were collected by a sampling frequency of every 1,000 generations and with the first 25% of the samples discarded as burn-in.
Results
Systematic Paleontology
Euarthropoda Lankester, 1904
Fuxianhuiida Bousfield, 1995
Constituent taxa. From the Chengjiang Biota (Cambrian Series 2, Stage 3): Fuxianhuia protensa Hou, 1987, Chengjiangocaris longiformis Hou & Bergström, 1991, Pisinnocaris subconigera Hou & Bergström, 1998, Liangwangshania biloba Chen, 2005, and Xiaocaris luoi Liu et al., 2020a; Xiaoshiba Biota (Cambrian Series 2, Stage 3): Fuxianhuia xiaoshibaensis Yang et al., 2013, Chengjiangocaris kunmingensis Yang et al., 2013, and Alacaris mirabilis Yang et al., 2018; Guanshan Biota (Cambrian Series 2, Stage 4): Guangweicaris spinatus Luo et al., 2007.
Family incert. Sedis
Genus Pisinnocaris Hou & Bergström, 1998
Type species. Pisinnocaris subconigera Hou & Bergström, 1998; by monotypy.
Emended diagnosis. As for type species by monotypy, see below.
Remarks. Pisinnocaris differs from other fuxianhuiids in the number of shortened prothoracic segments, with four such segments in Pisinnocaris, three in Fuxianhuia and Guangweicaris (Fuxianhuiidae), five in Chengjiangocaris and Alacaris (Chengjiangocariidae), and six in Liangwangshania. Among fuxianhuiids, Pisinnocaris has the lowest trunk segment number, only twelve (from prothorax to the posterior end of the body). All the other fuxianhuiids possess over twelve trunk segments, of which Liangwangshania bears more than forty trunk segments. Pisinnocaris is also unique in its conical body shape.
Pisinnocaris subconigera Hou & Bergström, 1998
1998 Pisinnocaris subconigera sp. nov.; Hou & Bergström, 1998: 395–401, text-fig. 2.
1999 Pisinnocaris subconigera Hou & Bergström, 1998; Hou et al., 1999: 91, pl. 124.
2018 Fuxianhuia protensa Hou, 1987; Fu et al., 2018, text-figs.1, 3, 13.
Type material: Holotype: NIGPAS 115417 (Fig. 1A). A small arthropod with large, subcircular carapace and tapering body with at least ten long tergites (Hou & Bergström, 1998).
Type locality and horizon: Eoredlichia Trilobite Zone of the Yu’anshan Member
(Chiungchussu Formation), Cambrian Series 2, Stage 3, Maotianshan Mountain, eastern Yunnan province.
Additional material: Since the original description of this species, few additional specimens have been reported (Xu, 2004; Zeng et al., 2014; Fu et al., 2018). To this we add fourteen specimens from the Xiaolantian section (shown in Fig. S2), China, Cambrian Series 2, Stage 3.
Emended diagnosis: Subcircular carapace, posterior margin without depression; anterior sclerite supporting stalked eyes; a pair of specialized post-antennal appendage; four shortened prothoracic segments; four opisthothoracic segments; four limbless abdominal segments; body gradually narrowed, terminated with a pair of leaf-like tail flukes.
Description
Dorsal morphology
Our specimens indicate that the body size of Pisinnocaris subconigera ranges from 8.92 to 47.62 mm, while the number of trunk segments remains unchanged (Figs. 1 and S1). An anterior sclerite bearing a pair of lateral eyes is located in front of the subcircular carapace that accounts for approximately 1/4–1/3 of the total body length. The anterior margin of the carapace is rounded, with its lateral margins curving inward to form the posterior margin. Unlike in other fuxianhuiids, the median portion of the posterior margin is straight without bending forward. The ratio of the carapace’s width to length is 1.45–1.88, representing the second highest among fuxianhuiids after F. protensa. Lateral wrinkles can be formed due to dorsal-ventral compression, indicating a slightly swollen morphology of the front-middle region of the carapace (red triangle indicated in Figs. 1A and 1D). Notably, the carapace is overlapping four shortened prothoracic segments that can only be observed when most of the carapace is missing (Fig. 1B).
The trunk of Pisinnocaris subconigera is composed of twelve segments that can be further divided into three tagmata: a prothorax, an opisthothorax, and a limbless abdomen (Fig. 1). The prothorax consists of four shortened segments (T1–T4) whose tergites gradually increase in width and length towards the posterior. The first prothoracic segment is the smallest and can be observed in two specimens: one with the structure fully exposed (Fig. 1B), and the other imaged with CT scanning and 3D rendering techniques (Fig. 2A). Length of this somite is less than 0.1 mm in both specimens. The last two segments of the prothorax (T3, T4) are 5–8 times wider than length and can also be observed in the CT scans of the largest specimen (Fig. 2E). The posterior margin of each preceding segments usually abuts against the anterior part of the following ones.
Morphological details of Pisinnocaris subconigera.(A) Screenshot of a 3D model derived from a CT scan of the holotype shown in Fig. 1A. Dorsal view showing four anterior shortened segments (T1–T4) and four pairs of appendages (white arrows) associated with T7. (B) Screenshot of a 3D model derived from a CT scan of YKLP 17301 shown in Fig. 1C. Ventral view showing the arrangement of the appendages. Note that there’s no appendage associated with T9 and the subsequent segments. (C) Close-up of the head in B, showing the eyes, the antennae, the SPAs, the hypostome, and the first four pairs of biramous appendages (en). Blue arrowhead indicates the eyestalk. (D) Close-up of the trunk in B, showing four flap-like exopods (ex) preserved on T6, along with an almost complete endopod in T7 (en). Red arrowhead indicates the distal podomere of the endopod in subtriangular shape. (E) Screenshot of a 3D model derived from a CT scan of CJHMD00066a shown in Fig. 1D. Ventral view showing a pair of SPAs with the hypostome (hy) in-between. The last two shortened prothoracic segments (T3, T4) are also detected. (F) Reconstruction of the ventral side of the head. (G) Reconstruction of a biramous appendage consisting of a flap-like exopo d (gray) and an endopod bearing at least ten podomeres (yellow). Abbreviations: an, antenna; asc, anterior sclerite; ca, carapace; ed, endite; en, endopod; ex, exopod; ey, eye; hy, hypostome; SPAs, specialized post-antennal appendages; Tn, trunk segment n. Scale bars: 1 mm.
The opisthothorax comprised of four segments (T5–T8) whose tergites are wider than those of the prothoracic ones, but becomes narrower towards the posterior of the body. The first opisthothoracic segment (i.e., T5) bears the widest tergite throughout the entire body (Fig. 1), with a width-to-length ratio of approximately 3.5. Its length is equivalent to all the following tergites. As in the prothorax, the posterior margin of each tergite overlaps the anterior margin of the following one. The overlapping area is approximately 1/3 of the total length of the tergite. Each opisthothoracic tergite bears a keel extending towards the posterior (Fig. 1D). Subsequent segments form the limbless abdomen which tapers and terminates with the last segment forming a sub-square in shape. A pair of leaf-like tail flukes are attached to the last abdominal segment (Figs. 1A and D).
Ventral morphology
Pisinnocaris subconigera bears a pair of small, ovoid eyes each connected to the ventral side of the anterior sclerite via a stalk (Figs. 1A–1C; 2A and 2C). Unlike Fuxianhuia protensa whose eyes, eye stalks, and anterior sclerite are located in front of the anterior margin of the carapace, these anteriormost structures of P. subconigera are partially covered by the carapace and thus less obvious from the dorsal perspective (Figs. 1 and 2). Similar to other fuxianhuiids, P. subconigera possesses a pair of antennae each consisting of a minimum of ten antennal podomeres. The proximal podomere is wider than its length, while the subsequent podomeres gradually taper and elongate, so the proximal portion exhibits a moniliform appearance, whereas the distal part displays a claviform shape. No antennal setae have been observed in any specimens studied herein, most likely due to the tiny size of them. A pair of specialized post-antennal appendages (SPAs) are only observable with the combination of CT scan and 3D rendering techniques (Figs. 2C and 2E). With the same techniques, a fractured hypostome is detected between the SPAs (Figs. 2C and 2E). Each prothoracic segment carries a small pair of biramous appendages (Figs. 2B and 2C). Four pairs of biramous appendages are found associated with every opisthothoracic segment (Figs. 2A, 2B and 2D). The biramous appendages gradually decrease in size towards the posterior of the body that terminates into a four-segmented abdomen without any appendages (T9–T12; Figs. 1 and 2B). All biramous appendages are composed of a flap-like exopod and an endopod bearing at least ten podomeres. Each podomere bears an endite medially (Figs. 2B, 2D and 2G). Similar to Guangweicaris (Chen et al., 2020), the distal podomere of the endopod in P. subconigera is in subtriangular shape (Red triangle indicated in Fig. 2D). This is different from the claw-like and the rounded distal podomere seen in Alacaris (Yang et al., 2018) and F. protensa (Fu et al., 2018), respectively.
Discussion
Morphological comparisons among fuxianhuiids and taxonomic validity of Pisinnocaris subconigera
Since the original report of Pisinnocaris subconigera in 1998, only a few additional specimens have been assigned or considered to be related to this species due to the rarity and poor preservation of the specimens (Xu, 2004; Zeng et al., 2014). We hypothesize that the similarities shared by different members of fuxianhuiids might have led to misidentify Pisinnocaris as a juvenile stage of Fuxianhuia protensa (Fu et al., 2018).
While sharing common features with other fuxianhuiids, Pisinnocaris subconigera also bears unique morphological characters to support its taxonomic validity (Table 1). According to our study, the most diagnostic feature for P. subconigera is considered to be the prothorax composed of four shortened segments instead of three. The number of prothoracic segments is key to identify fuxianhuiids at the genus or even family levels. Notably, the tapering body shape of P. subconigera is similar to Chengjiangocaris and Liangwangshania, but different from Fuxianhuia and Guangweicaris whose abdomen is much narrower than the opisthothorax. From the tapering shape of the posterior segments in P. subconigera, one would assume those segments to be limb-bearing ones and, hence, represent part of the opisthothorax. Computed tomography (CT) scans, however, reveal no limbs for any of those segments, suggesting an abdominal identity for this tagma (Fig. 2B). Therefore, shape of the posterior segments (abrupt narrowing vs tapering) alone cannot serve as an indicator for the presence/absence of an abdomen in fuxianhuiids.
Table 1: Morphological comparisons among fuxianhuiids.
Phylogeny
In order to clarify the phylogenetic position of Pisinocaris subconigera, we also performed matrix-based cladistic analysis. Both the trees from Maximum Parsimony (Figs. 3A and B) and Bayesian Inference (Fig. 3C) show that phylogenetic position of P. subconiger is consistent—inside the clade of fuxianhuiids. The interrelationship of different members of fuxianhuiids shown in the tree of implied weights is consistent with the Bayesian tree (Figs. 3A and 3C): among the nine fuxianhuiid members, Liangwangshania biloba is the sister group to all other fuxianhuiids; A. mirabilis, C. longiformis and C. kunmingensis cluster into a branch and F. protensa, F. xiaoshibaensis, X. luoi and G. spinatus cluster into another branch on the trees. P. subconigera exhibits a closer phylogenetic affinity to Fuxianhuiidae (Hou & Bergström, 1997) on the tree of implied weights. However, it is positioned closer to the stem lineage within the clade of fuxianhuiid on the Bayesian tree. The interrelationship of fuxianhuiids is less well-resolved in the tree of equal weights. The unclear details of the morphology, especially in the appendages of several other fuxianhuiids may be the reasons for this discrepancy. But this tree still supports that P. subconigera is a member of fuxianhuiids. By analyzing the characteristics at several nodes, it can be seen that the number of prothoracic segments plays a crucial role in classification. It is not only the most characteristic feature for fuxianhuiids as a whole being set apart from all the other Cambrian euarthropods, but also the feature that divides fuxianhuiids into four major groups (Table 1). Namely, there are six prothoracic segments in Liangwangshania, five in Chengjiangocarididae (Hou & Bergström, 1997), four in P. subconigera, and three in Fuxianhuiidae, suggesting may be four families in fuxianhuiids which currently include nine species in seven genera.
Phylogenetic analysis of fuxianhuiids.(A) Strict consensus of parsimonious trees calculated under implied weights (k = 3). (B) Strict consensus of parsimonious trees calculated under equal weights. (C) Bayesian phylogenetic analyses. A blue bar represents the clade of fuxianhuiids, P. subconigera is highlighted in red, the red box highlights that the species has six prothoracic segments, the green box highlights that the species has five prothoracic segments and the yellow box highlights that the species has three prothoracic segments.
Larval/juvenile forms
All specimens of Pisinocaris subconigera studied here have the same conical body shape and trunk segment number throughout the entire body and also within each tagma, despite the difference in their body size (Fig. S1). Combined with the discovery of the largest individual of P. subconigera with a total body length of 47.62 mm and the prothoracic segments and appendage morphology of other individuals revealed by micro-CT, we conclude that P. subconigera is not the larval or juvenile form of any other fuxianhuiids, but is a valid species on its own.
Another reason for considering P. subconigera a synonym of juvenile F. protensa is the co-occurance of P. subconigera and an adult of F. protensa on the same fossil slab (fig. 13 in Fu et al., 2018). Similar slabs are also shown here, on which P. subconigera and the juvenile of F. protensa (maybe stage 24 or 25, Figs. 4B and 4D) have similar body size, but differences in body shape and trunk segmentation between the two species are clearly observable (Fig. 4). As a consequence, it remains an open question whether extended parental care exists in F. protensa. We cannot rule out the possibility that they may simply be preserved together. More specimens of F. protensa with different developmental stages are key to clarifying the situation.
Pisinnocaris subconigera and Fuxianhuia protensa preserved together.(A) YRCP-R-0001, from Maotianshan, Chengjiang, showing several small F. protensa and P. subconigera were preserved together. (B) The white-framed part in A, showing a F. protensa (left one marked with white arrow) with about twenty-four trunk segments and a P. subconigera (right one marked with red arrow indicator) of eight trunk segments. The white arrow shows the smallest individual with at least twelve segments. (C) YRCP-R-0024, from Maotianshan, Chengjiang, showing several small F. protensa and P. subconigera were preserved together. (D) is the white-framed part in C, showing a F. protensa (left one marked with white arrow) with about twenty-five trunk segments and a P. subconigera (right one marked with red arrow) with eight trunk segments. Scale bars: 5 mm.
In fossil research, it is extremely difficult to accurately identify the larval/juvenile and adult forms due to the limited number of specimens and the preservation. The specimens of the same species often vary greatly in size, leading to predictions about larval, juvenile or adult forms. In addition to appearance and preservation of fossils, more details of morphological characteristics need to be considered, especially when conducting studies on ontogeny, such as changing trunk segmentation and morphological features of appendages (Fu et al., 2018; Liu et al., 2020b and Zhai et al., 2019). The same situation exists in fuxianhuiids. Small individual specimens have been found and they were presumed to the larval or juvenile form of fuxianhuiids (Fig. 5) often based on being highly similar in appearance. For example, in Fig. 5A, the large fossil individual has been identified as C. longiformis, and the small one exhibits identical morphological traits, including its long and narrow body, carapace shape, and tail structure. L. biloba, with the small individual specimens shown in Fig. 5B and G. spinatus with the small individual specimens shown in Fig. 5C display similar phenomena. In fact, we need more evidence, especially details of appendage morphology, to identify the species of these three small individuals and determine whether they are larve or juveniles, just as we did with P. subconigera in this study.
Adult and presumed larval/juvenile specimens of other fuxianhuiids.(A) Chengjiangocaris longiformis (YKLP17302) and a presumed larva/juvenile (YKLP17303). (B) Liangwangshania biloba (YKLP17304) and a presumed larva/juvenile (YKLP17305). (C) Guangweicaris spinatus (YKLP11566-arthro) and a presumed larva/juvenile (YKLP17306). Scale bars: 5 mm.
Conclusions
- Pisinnocaris subconigera is a valid species of early Cambrian fuxianhuiid, not a larval/juvenile form of any other fuxianhuiid including Fuxianhuia protensa.
- Study of larval/juvenile forms of fossil species requires not only more specimens from various developmental stages but also consideration of more details of morphological characteristics.
Supplemental Information
10.7717/peerj.20483/supp-1Supplemental Information 1Supplementary Materials.
10.7717/peerj.20483/supp-2Supplemental Information 2Different size of P. subconigera individual.(A) A total of fourteen individuals are arranged from left to right according to their sizes. The specimen numbers are, in order: YRCP-R-0013-D, YRCP-R-0027-J, YRCP-R-0024-D, NIGPAS 115417a, YRCP-R-0007-A, YRCP-R-0024-J, YRCP-R-0001-D, YRCP-R-0020-F, YRCP-R-0019-D, YRCP-R-0036b, CJHMD00070, YRCP-R-0034, YKLP17301 and CJHMD00066a. (B) Scatterplot of the fourteen individuals, X axis represents the body length and Y axis represents the width of T5, T5 bears the widest tergite throughout the entire body. Trend line with blue dotted suggested that length-to-width ratio of different sizes individuals tends to be consistent.
10.7717/peerj.20483/supp-3Supplemental Information 3Distribution of the main localities of the Chengjiang biota, Yunnan Province and the stratum.(A) Red box shows the “Xiaolantian” section. (B) Stratigraphic column of the main localities of the Chengjiang biota (revised from Hou et al., 2017 and Jin et al., 2024)
10.7717/peerj.20483/supp-4Supplemental Information 4Information of the specimens analyzed in the present study.
10.7717/peerj.20483/supp-5Supplemental Information 5Characters list.
10.7717/peerj.20483/supp-6Supplemental Information 6Matrix for TNT.
10.7717/peerj.20483/supp-7Supplemental Information 7Matrix for Mrbayes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aria C Zhao F Zhu M Fuxianhuiids are mandibulates and share affinities with total-group Myriapoda Journal of the Geological Society 20211785 jgs 2020–jgs 224610.1144/jgs 2020-246 · doi ↗
- 2Chen A A new Fuxianhuia-like arthropod of the early Cambrian Chengjiang fauna in Yunnan Yunnan Geology 20052401108113(in Chinese with English abstract)10.3969/j.issn.1004-1885.2005.01.011 · doi ↗
- 3Chen H Restudy of the fuxianhuiid arthropods from Cambrian Series 2, Eastern Yunnan. Ph.D. dissertation, Yunnan University, Tunnan, China (in Chinese with English abstract)2020
- 4Chen A Chen H Legg DA Liu Y Hou X A redescription of Liangwangshania biloba Chen, 2005, from the Chengjiang biota (Cambrian, China), with a discussion of possible sexual dimorphism in fuxianhuiid arthropods Arthropod Structure & Development 201847555256110.1016/j.asd.2018.08.00130125735 · doi ↗ · pubmed ↗
- 5Chen H Legg DA Zhai D Liu Y Hou X New data on the anatomy of fuxianhuiid arthropod Guangweicaris spinatus from the lower Cambrian Guanshan Biota, Yunnan, China Acta Palaeontologica Polonica 202065113914810.4202/app.00508.2018 · doi ↗
- 6Du K Ortega-Hernández J Yang J Zhang X A soft-bodied euarthropod from the early Cambrian Xiaoshiba Lagerstätte of China supports a new clade of basal artiopodans with dorsal ecdysial sutures Cladistics 201935326928110.1111/cla.1234434622993 · doi ↗ · pubmed ↗
- 7Fu D Ortega-Hernández J Daley AC Zhang X Shu D Anamorphic development and extended parental care in a 520 million-year-old stem-group euarthropod from China BMC Evolutionary Biology 201818111710.1186/s 12862-018-1262-630268090 PMC 6162911 · doi ↗ · pubmed ↗
- 8Gabbott SE Hou X Norry MJ Siveter DJ Preservation of early Cambrian animals of the Chengjiang biota Geology 20043290190410.1130/G 20640.1 · doi ↗