Parasite genotypes and host immune mediators in primary and recurrent episodes of vivax Malaria in Colombia
Yesica Yamile Duque-Isaza, Jaime Carmona-Fonseca, Eliana María Arango-Flórez, Yesica Yamile Duque-Isaza, Jaime Carmona-Fonseca, Eliana María Arango-Flórez

TL;DR
The study found that pregnant women in Colombia experience more frequent and genetically diverse recurrences of vivax malaria compared to non-pregnant individuals.
Contribution
The study identifies higher recurrence rates and parasite genetic diversity in pregnant women with vivax malaria and links this to immune mediator expression.
Findings
Pregnant women had a higher recurrence rate (41.6%) compared to non-pregnant individuals (8.7%).
Parasite genetic diversity was higher in pregnant women, but no exclusive alleles were found.
Fox-P3 immune mediator expression was significantly higher in primary infections in pregnant women.
Abstract
To identify Plasmodium vivax genotypes in the pregnant and non-pregnant population and explore their association with immune mediators. Two cohorts of patients with uncomplicated vivax malaria were followed for 120 days in Puerto Libertador and Tierralta, Cordoba, Colombia: 41 pregnant women and 46 non-pregnant individuals, all treated with standard treatment. Parasite genotypes (microsatellites Pv3.27, Pv3.502, and Pv1.501) and the expression of host immune mediators (IL-13, IL-10, TNF-alpha, IFN-gamma, IL-8, TGF-beta, Fox-P3, PD-L1) were compared between primary and recurrent infections. The frequency of recurrences was higher in pregnant women (41.6%) than in non-pregnant individuals (8.7%). Parasite genetic diversity was higher in pregnant women, although without exclusive alleles. Recurrences were genetically homologous (same alleles) in only 23% and 33% of cases in pregnant and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
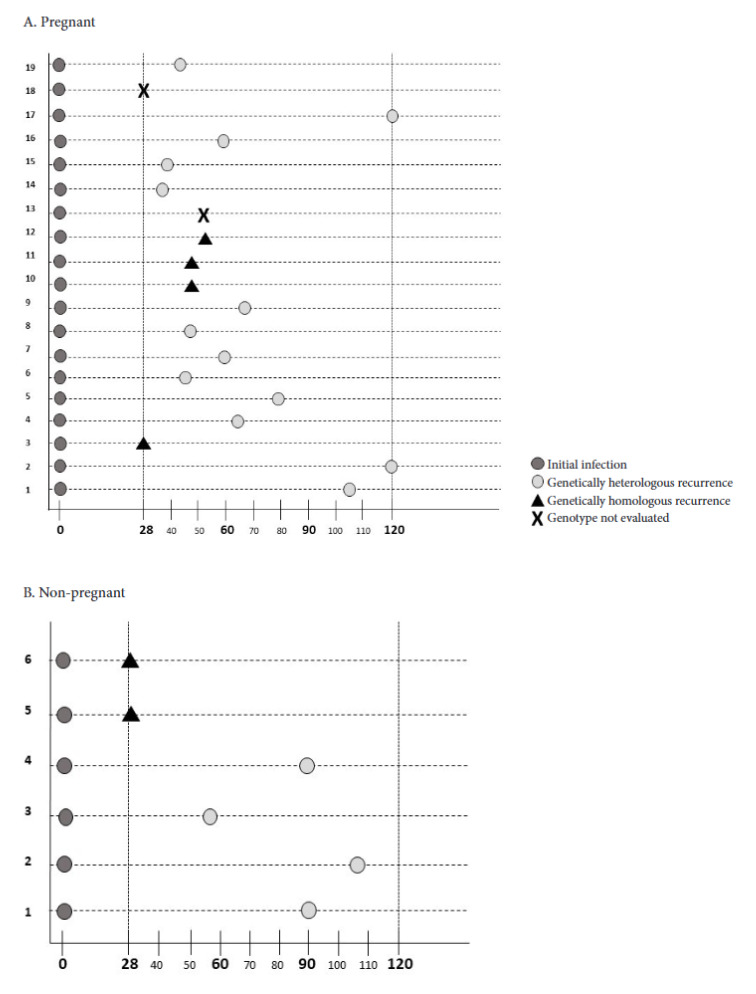 Figure 1
Figure 1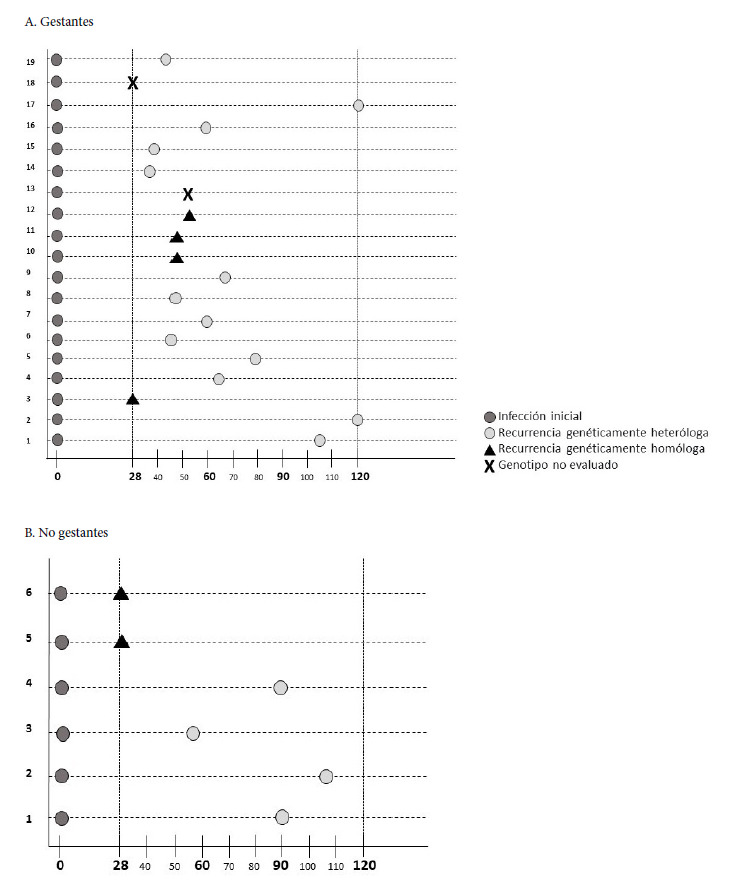 Figure 2
Figure 2- —Ministry of Science and Technology of Colombia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Vector-borne infectious diseases · Sperm and Testicular Function
INTRODUCTION
Infection by Plasmodium vivax during pregnancy is associated with disease in the woman (gestational malaria), placental damage (placental malaria) 1, and adverse maternal and perinatal outcomes 2^,^3. It is estimated that, each year, more than 90 million pregnancies worldwide are at risk of P. vivax infection 4. In particular, in the American continent, this species is responsible for more than 80% of malaria cases 5, with reports from northwestern Colombia documenting a frequency of gestational malaria of 35.8% and placental malaria between 20.9% and 27.7% 6^,^7. Studies that contribute to the knowledge of the genetics and pathogenicity of the parasite, as well as the host immune response, are indispensable for understanding the pathophysiological processes of this species. In general, knowledge about P. vivax is much less advanced than that of P. falciparum. However, available reports leave no doubt about the pathogenic role of P. vivax during gestation nor about the morbid effects it produces in pregnant women and their children 1^-^3^,^8^,^9.
All Plasmodium species can cause malaria recurrences, understood as the reappearance of parasitemia, with or without associated symptoms, after having received specific antimalarial treatment 10^,^11. These recurrences can be of three types: A) recrudescence, caused by the incomplete elimination of erythrocytic stages (therapeutic failure). It usually appears within 28 days of initiating treatment and can be caused by any species of Plasmodium10^,^12. B) Reinfection, generated by the inoculation of new sporozoites by the vector. It occurs after 28 days of initiating treatment in patients with complete cure of the initial episode, and can be caused by any species 10^,^12^,^13. C) Relapse, caused by the activation of hypnozoites (latent hepatic stages) and appears after 28 days of initiating treatment for P. vivax or P. ovale malaria, in patients who had a complete resolution of the primary episode 10^,^12^-^14.
Other suggested sources of relapses are erythrocytic stages sequestered in the spleen and bone marrow 15, sporozoites that survive in the skin, and merozoites present in the lymphatic system 16. In patients coming from non-endemic areas, the differentiation between recrudescence, reinfection, and relapse can be performed through clinical and parasitological follow-up. However, in residents of endemic areas, this differentiation is limited, since currently there are no accessible tools available that allow distinguishing reliably between relapses and reinfections 17.
Primaquine (8-aminoquinoline) as a hypnozoiticide, in combination with chloroquine (4-aminoquinoline), which acts as a blood schizonticide, constitutes the treatment indicated to cure vivax malaria and prevent relapses in Colombia and in most endemic areas. However, primaquine is contraindicated during pregnancy and lactation due to the risk of dose-dependent hemolysis in fetuses and newborns with undiagnosed glucose-6-phosphate dehydrogenase deficiency 18^,^19.
The quantity of recurrences by P. vivax could be influenced by immunity acquired during the primary episode and by the immunological history of previous episodes 20. Patients acquire natural immunity after primary infection, and experimental studies have demonstrated that immune protection is clone-specific 21. However, available information on the immune response in malaria recurrences is scarce 22^,^23. In Thailand, it was observed that both proinflammatory and anti-inflammatory cytokines presented higher levels in patients with P. vivax than in healthy individuals. Furthermore, in patients with homologous or heterologous relapses, IL-1β levels were higher during primary infection, while IL-6 and IL-10 levels were higher in heterologous relapses than in homologous ones 21.
The genetic diversity of P. vivax is much greater than that of P. falciparum24^,^25. Genotypically, malaria recurrences can be homologous or heterologous: in homologous ones, the parasite genotypes of the primary episode and the recurrence are exactly the same 26^,^27, while in heterologous ones, different parasite genotypes are found in the primary episode and in the recurrence 27. In Colombia, few studies have compared P. vivax genotypes between primary infection and recurrence 28^,^29, and none have been conducted in the pregnant population. The human host immune response to plasmodial infection is very complex, and when it occurs during gestation, this complexity is even greater due to the hormonal and immunological changes that occur during pregnancy, where it is necessary to maintain a balance between the proinflammatory and anti-inflammatory immune response 30.
This study aimed to compare P. vivax genotypes between the primary episode and the recurrence in the pregnant and non-pregnant population, as well as to explore the association between parasite genotypes present in those malaria episodes with the expression of proinflammatory and anti-inflammatory immune mediators in both groups.
KEY MESSAGESMotivation for the study. Vivax malaria constitutes a relevant challenge in public health in Colombia, especially in pregnant women, who are more prone to recurrences and complications. However, the genetic diversity of the parasite in primary and recurrent infections, as well as its relationship with the host immune response, has been scarcely studied.Main findings. Pregnant women presented a higher frequency of recurrences (41%) and a greater genetic diversity of the parasite compared to non-pregnant individuals. Likewise, a higher expression of Fox-P3 was observed during the primary infection of pregnant women, which may modulate protective immunity.Public health implications. Strengthening the prevention and control of P. vivax malaria in pregnancy is crucial to decrease recurrences and improve maternal and neonatal outcomes.
MATERIALS AND METHODS
Study site and population
This research was executed in Puerto Libertador (7°53′17″N 75°40′25″W) and Tierralta (8°10′22″N 76°03′34″W), in the department of Cordoba, Colombia. Both belong to the second malaria transmission focus in Colombia (Uraba-Bajo Cauca-Sinu-San Jorge), where P. falciparum and P. vivax co-circulate, although P. vivax predominates, with 65% of cases 31^,^32. Approximately 51% of vivax malaria cases reported in Colombia come from this region. Between the years 2000 and 2023, 674,294 accumulated cases were registered, with an annual parasite index for P. vivax (number of cases per 1000 inhabitants) of 13.9 32.
Study design
A descriptive, prospective, and longitudinal study was carried out, with two cohorts: 41 pregnant women and 46 non-pregnant men and women, all volunteers with a diagnosis of uncomplicated malaria caused exclusively by P. vivax. The infection was detected by thick blood smear (TBS) and confirmed by quantitative real-time PCR (qPCR). The inclusion of participants was progressive between 2016 and 2019, both at the malaria diagnostic post of the local hospital and in prenatal consultation.
Selection criteria
Inclusion criteria
Pregnant women of any gestational age were invited to participate, as well as non-pregnant men and women, who presented single infection by P. vivax confirmed by TBS and qPCR, who were classified as uncomplicated malaria cases, and who voluntarily consulted at the local hospital between 2016-2019. All participants consented to their voluntary participation by signing the informed consent, and in cases of minors, informed assent was also signed.
Exclusion criteria
Pregnant women with a history of comorbidities compatible with TORCHS syndrome, as registered in the clinical history, were excluded, as well as patients (pregnant or non-pregnant) who had received any antimalarial other than the standard treatment prescribed at the local hospital. Those who withdrew consent, developed any malaria complication during follow-up, or reported pregnancy during the course of the study within the non-pregnant cohort were also excluded.
Antimalarial treatment
The treatment supplied to the volunteers of this study was that recommended by the World Health Organization (WHO) and the health authorities of Colombia for uncomplicated P. vivax malaria. In non-pregnant patients, the combination of chloroquine for three days (total dose of 25 mg/kg) plus primaquine for 14 days (daily dose of 0.25 mg/kg) was administered. In pregnant women, only chloroquine was prescribed (total dose of 25 mg/kg divided over three days), due to the contraindication of primaquine during pregnancy 18^,^33. The treatment was dispensed by the staff of the malaria diagnostic post of the local hospital, and its administration was unsupervised.
Sample collection and follow-up
On the day of admission to the study (day 1), before the administration of the first dose of antimalarial treatment, venopuncture was performed on each volunteer to obtain a peripheral blood sample, to prepare: a) two TBS slides for microscopic diagnosis of malaria; b) three circles of Whatman® No. 3 filter paper for DNA extraction for confirmation of single P. vivax infection by qPCR and for genotyping of parasites with microsatellites; c) two vials, each with 100 μL of the leukocyte pellet mixed with 900 μL of TRIzol (Invitrogen), preserved in liquid nitrogen until use in RNA extraction for the quantification of immune mediator expression.
Follow-up for the detection of recurrences was performed on days 28, 60, 90, and 120 after the start of antimalarial treatment. At each follow-up, malaria symptomatology was evaluated and venopuncture was performed to collect the same samples obtained on the day of admission. Unscheduled visits by volunteers between days 2 and 120 were also considered for the detection of recurrences; that is, if any volunteer presented a positive TBS or malaria symptomatology on days other than 28, 60, 90, or 120, it was also considered a malaria recurrence and, on that day, the same blood samples as on the day of admission were collected.
Diagnosis of plasmodial infection
TBS slides were stained with Field stain and read by an experienced microscopist technician from the local hospital, following WHO recommendations 34. For parasitemia calculation, a parasite count was performed in a total of 200 leukocytes, using the constant of 8000 leukocytes/μL.
Confirmation of single P. vivax infection was done by qPCR, using the protocol and primers and probes for the 18S gene, which were designed by researchers from Edmonton, Canada 35. DNA extraction from filter paper was done using the Saponin- Chelex method previously described by other researchers 36.
Parasite genotyping
Amplification of microsatellites Pv3.27, Pv3.502, and Pv1.501 was done by semi-nested PCR according to Koepfli et al. 37, with some modifications: 1X PCR buffer (Qiagen), 3 mM (Pv3.27) or 3.5 mM (Pv3.502, Pv1.501) of MgCl2 (Qiagen), 200 μM of each deoxynucleotide triphosphate (dNTP) (Takara Bio), 0.25 μM of each primer, and 0.067 IU/ μL of recombinant Taq DNA polymerase (Fermentas). To confirm adequate amplification, 2 μL of each product were run on 2% agarose gel electrophoresis, stained with GelRed™ (Biotium), and bands were visualized on a UV transilluminator.
Analysis of the size of amplified products was performed by capillary electrophoresis on a 3500 genetic analyzer (Hitachi Applied Biosystems), with the internal standard GeneScan 600 LIZ (Applied Biosystems) and factory conditions for microsatellites. PCR products were diluted in Milli-Q water, 1:10 (Pv3.502) or 1:20 (Pv1.501 and Pv3.27). Electrophoresis data were analyzed with GeneMapper ID-X Software v1.5 (Applied Biosystems) and electropherograms were examined visually. Peaks above 300 relative fluorescence units (RFU) were considered true amplifications. Alleles were grouped manually according to their size, as follows: groups of 4 bp for Pv3.27; 7 bp for Pv1.501, and 8 bp for Pv3.502 38. Polyclonal infection was established when more than one peak was observed at a locus and the height of the minor peak was greater than 33% of the height of the predominant peak 39.
Classification of recurrences and comparison of parasite genotype
Recrudescence was considered when parasites were detected by TBS or qPCR between days 4 and 28 after initiating antimalarial treatment. Relapse/reinfection was considered when parasitemia (TBS or qPCR) was detected at any time between days 29 and 120 after initiating treatment.
For the comparison of parasite genotypes between isolates from the day of recurrence (detected between day 4 and 120 after initiating treatment) and those from the primary episode (detected on the day of admission or day 1), recurrences were considered genetically homologous when exactly the same alleles were detected in all microsatellites at both time points (primary episode and recurrence), and when allelic variants were detected in at least one microsatellite, they were classified as genetically heterologous recurrences 21^,^40.
Relative quantification of immune mediator expression
First, total RNA extraction was performed from the leukocyte pellet homogenized in TRIzol (Invitrogen), maintained at refrigeration temperature and following the manufacturer’s instructions. Then, relative quantification of the expression of IL-13, IL-10, TNF-α, IFN-γ, IL- 8, TGF-β, Fox-P3, PD-L1 was performed with the commercial kit Express OneStep Superscript® qRT-PCR kit (Invitrogen), on the StepOnePlus real‐time PCR system (Applied Biosystems), with the protocol previously reported 41.
Statistical analysis
Microsoft Excel v.10 and GraphPad Prism v.8 were used for analysis. No quantitative variable had parametric distribution, nor homogeneity of variances, according to Shapiro-Wilk and Levene, respectively; therefore, differences between unpaired groups (pregnant vs. non-pregnant and with recurrences vs. without recurrences) were evaluated with Mann-Whitney U and between paired groups (initial episode vs. recurrence) with Wilcoxon. Qualitative variables were compared with Chi-square. To analyze genetic diversity, the number of alleles per locus, the predominant allele, the percentage of polyclonal samples, and the expected heterozygosity (He) were calculated (He = [n/(n - 1) ] [1 - Σpi2]; n, number of alleles per locus and pi, frequency of allele i in the population). Statistical significance was always considered with p less than 0.05.
Ethical considerations
This study was considered minimum risk (Resolution 8430 of 1993) and was approved by the Bioethics Committee of the Medical Research Institute of the Faculty of Medicine of the Universidad de Antioquia (act 012 of August 23, 2018).
RESULTS
A total of 41 pregnant women (23 from Tierralta and 18 from Puerto Libertador) and 46 non-pregnant men and women (31 from Tierralta and 15 from Puerto Libertador) with uncomplicated vivax malaria, diagnosed by TBS and confirmed by qPCR, participated in this study. Pregnant women had an average age of 21 years (range: 14-42 years) and 22% (9/41) were in the first trimester, 54% (22/41) in the second trimester, and 24% (10/41) in the third trimester. In the non-pregnant group, 63% (29/46) were men and 37% (17/46) women, with an average age of 28 years (range: 8-84 years). Age and parasitemia both at the time of admission and recurrence were similar between pregnant and non-pregnant individuals (Table 1).
Table 1. Age and parasitemia levels in the primary episode and recurrence for the cohort of pregnant and non-pregnant patients with uncomplicated P. vivax malaria.VariablesPregnant (n=41) Non-pregnant (n=46) p (M-W)MinMaxMedIQRMinMaxMedIQRAge (years)14421917-238842114-430.670Parasitemia (p/µL) in initial episode633340041801600-80327202950024031738-48150.296Parasitemia (p/µL) in recurrent episode2862178216701110-484111502053021301173-153480.753Min: minimum, Max: maximum, Med: median, p/uL: parasites/microliter, IQR: interquartile range, p (M-W): p-value from Mann-Whitney U test.
Among the 41 pregnant women studied, 4.9% (n=2) presented recrudescences (parasitemia detected between day 4 and 28 after initiating antimalarial treatment), while the remaining 95.1% (n=39) were classified as therapeutic successes (no parasitemia, nor malaria symptoms between days 4 and 28 after initiating treatment). In the non-pregnant cohort, 4.3% (n=2) had recrudescence and 95.7% (n=44) were therapeutic successes.
The 120-day follow-up allowed the detection of 21 relapses/ reinfections in the total of participants with therapeutic success (n=83), that is, detection of parasitemia between days 29 and 120 after initiating treatment, for a prevalence of relapses/reinfections of 25.3% (21/83). However, in the pregnant cohort, the prevalence of relapses/ reinfections was 43.6% (17/39), while in the non-pregnant cohort it was 9.1% (4/44).
Table 2 shows P. vivax genetic diversity data, with each microsatellite analyzed and in each cohort studied, both in the primary episode and in the recurrence. The following variables showed statistically significant differences between pregnant and non-pregnant individuals, both in the primary and recurrent episode: amplification success percentage, number of different alleles detected, and percentage of polyclonal infections. Furthermore, in the pregnant cohort, the following variables showed differences between the primary episode and the recurrence: number of different alleles detected, predominant allele, and percentage of polyclonal infections. In the non-pregnant cohort, significant differences were found in the number of alleles detected, the percentage of polyclonal infections, and the He between the primary and recurrent episode.
Table 2. Parasite genetic diversity detected with the three microsatellites in each studied cohort (pregnant and non-pregnant) and in each evaluated episode (primary episode and recurrence). Data per microsatellite
1.5013.5023.271.5013.5023.27Median per cohort DPrimary episode (enrollment, day 1)Pregnant n=41 Non-pregnant n=46 GestanteNo gestante % amplification success9590936380919380Yes# of detected alleles199349626199YesPredominant allele (bp)10614433685144288--No% polyclonal samples171749247541724YesEcpected heterozygosity (He)0.950.930.980.920.800.950.950.92NoRecurrent episodePregnant n=19 Non-pregnant n=6 PregnantNon-pregnant amplification success95799583831009583Yes# of detected alleles882323883YesPredominant allele^a^ (bp)120168288 36885128 152 168260 288--No% polyclonal samples 371158017333717YesExpected heterozygosity (He)0.980.950.980.961.000.980.980.98NoD: statistically significant difference (p < 0.05) between pregnant and non-pregnant women evaluated with Mann-Whitney U or Chi-square tests, as appropriate for quantitative or qualitative variables.a In the recurrence, there was more than one predominant allele in microsatellites Pv3.502 and Pv3.27.
Additionally, the number of alleles detected was significantly higher in pregnant women than in non-pregnant individuals, both in the primary episode and in the recurrence, and in pregnant women, furthermore, the predominant allele detected in the primary episode was totally different from the predominant allele detected in the recurrence; while in non-pregnant individuals, some predominant alleles were the same between the primary episode and recurrence. However, no allele was exclusively detected in any cohort; all alleles were shared.
The total of malaria recurrences was more than double in pregnant women (46%, 19/41) than in non-pregnant individuals (13%, 6/46), for a ratio of 3.5:1. In pregnant women, recurrences were of the relapse/reinfection type in 89% of cases (17/19) and of the recrudescence type in 11% of cases (2/19), for a ratio of 8.1:1. While in non-pregnant individuals, recurrences were relapses/reinfections in 67% of cases (4/6) and recrudescences in 33% of cases (2/6), for a ratio of 2:1. Additionally, recrudescences had a ratio of 1:3 between pregnant and non-pregnant individuals (11%:33%) and relapses/reinfections had a ratio of 1.3:1 between pregnant and non-pregnant individuals (89%:67%) (Table 3).
Table 3. Frequency and type of malarial recurrences in the cohort of pregnant and non-pregnant women.Type of recurrencePregnantNon-pregnantTotalp (χ²)Recrudescence (between days 4 and 28)2/19 (11%)2/6 (33%)4 (16%)0.184Relapse/reinfection (between days 29 and 120)17/19 (89%)4/6 (67%)21 (84%)0.004Total recurrences19/41 (46%)6/46 (13%)25 (29%)<0.001p (χ^2^): p-value of the Chi-square test for the comparison between pregnant and non-pregnant women
It was not possible to compare the parasite genotype between the primary and recurrent episode in two of the 19 pregnant women with malaria recurrence (11%), because adequate amplification of microsatellites was not obtained in one of the two episodes. In the remaining 17 pregnant women with recurrence, it was found that 23% had recurrences homologous to the initial episode and 77% had heterologous recurrences. For their part, in the six recurrent episodes detected in the non-pregnant cohort, it was found that 33% corresponded to homologous recurrences and 67% to heterologous recurrences. In summary, in both cohorts, heterologous recurrences are widely predominant, but the predominance is greater in pregnant women (3.3:1) than in non-pregnant individuals (2:1) (Figure 1).
Figure 1. Timing of recurrence detection and comparison of the parasite genotype between admission and recurrence in each study cohort. A. Pregnant cohort: primary episode, not evaluated (2/19), heterologous recurrence (13/17), homologous recurrence (4/17). B. Non-pregnant cohort: primary episode, heterologous recurrence (4/6), homologous recurrence (2/6).Y-axis: represents the number of each volunteer with recurrence. X-axis: follow-up day
It was not possible to determine the expression of immune mediators in all samples, because in some cases it was not possible to collect the sample in TRIzol or adequate amplification could not be achieved. Table 4 shows the comparison of immune mediator expression between the initial episode and the recurrence of both cohorts, in the samples that were successfully analyzed. The only immune mediator that showed significant differences in expression between the primary episode and the recurrence was Fox-P3, and only in the pregnant cohort, who showed significantly higher levels of Fox-P3 in the initial episode, compared to the recurrence (Table 4).
Table 4. Comparison of the relative expression of immune mediators in maternal peripheral blood between enrollment and recurrence in each studied cohort.Pregnant (n=34) Mediator (RU)Enrollment (n=34) Recurrence (n=13) Enrollment / recurrencep (M-W)MedianIQRMedianIQRTNF-α3.160.24-110.450.27-3.747.00.316IFN-γ8.271.82-166.674.79-7.761.20.249IL-81.330.60-2.702.280.10-280.60.764IL-1319.815.80-3213.622.55-421.50.650TGF-β6.510.52-211.981.01-293.30.986IL-100.170.07-0.700.10.03-0.521.70.199PD-L10.50.06-4.601.520.44-180.30.090Fox-P30.70.29-2.080.260.09-0.802.70.020Non-pregnant (n=30) Mediator (RU)Enrollment (n=30) Recurrence (n=2) Enrollment / recurrencep (M-W)MedianIQRMedianIQRTNF-α0.030.1-0.180.140.01-0.260.20.891IFN-γ3.210.80-7.012.771.03-4.511.20.896IL-82.671.28-5.843.590.56-6.620.70.901IL-131.880.64-3.201.240.76-1.721.50.541TGF-β0.020.01-0.170.230.01-0.440.10.904IL-10----
PD-L180.3238-19076.0617.52-1351.10.640Fox-P30.790.30-1.242.460.89-4.020.30.225IQR: interquartile range, RU: relative units of expression. Enrollment/recurrence: ratio between the median at enrollment and recurrence, p (M-W): p-value from Mann-Whitney U test.
The comparison of immune mediator expression between pregnant and non-pregnant individuals both at admission (primary episode) and at recurrence is presented in Table 5. In summary, at admission, pregnant women showed significantly higher levels in the expression of TNF-α, IFN-ɣ, IL-8, IL-13, and TGF-β than non-pregnant individuals, while PD-L1 had higher expression in non-pregnant individuals than in pregnant women. In the recurrent episode, TGF-β expression was significantly higher in pregnant women than in non-pregnant individuals, and the other immune mediators had similar expression in both cohorts (Table 5).
Table 5. Comparison of the relative expression of immune mediators in peripheral blood between the cohort of pregnant and non-pregnant women at the time of enrollment and malarial recurrence. Enrollment (n=64)
Mediator (RU)Pregnant (n=34) Non-pregnant (n=30) Pregnant/non-pregnantp (M-W)MedianIQRMedianIQRTNF-α0.870.18-3.920.030.01-0.2629<0.001IFN-γ8.256.50-9.973.370.79-7.112.4<0.001IL-85.201.20-9.161.610.72-2.353.20.006IL-1317.314.91-291.880.76-3.229.2<0.001TGF-β4.220.27-160.020.01-0.17210<0.001PD-L10.340.06-4.6181.2446-2020.004<0.000Fox-P30.820.31-2.140.750.29-1.171.10.312 Recurrence (n=15)
Mediator (RU)Pregnant (n=13) Non-pregnant (n=2) Pregnant/non-pregnantp (M-W)MedianIQRMedianIQRTNF-α 0.450.22-3.740.140.01-0.273.20.153IFN-γ6.165.02-7.522.771.03-4.522.20.121IL-80.970.30-4.013.590.56-6.620.20.923IL-1314.693.51-461.240.76-1.7211.80.153TGF-β2.480.97-350.230.01-0.4510.80.044PD-L12.800.41-1976.0618-1350.030.181Fox-P30.250.08-0.892.460.89-4.020.10.102IQR: interquartile range, RU: relative units of expression, Pregnant/non-pregnant: ratio between the median of pregnant and non-pregnant women, p (M-W): p-value from Mann-Whitney U test.
To explore the association between the profile of immune mediators with the parasite genotype of the primary episode and the recurrence, data from participants who had complete paired information of parasite genotyping (the three microsatellites) and immune mediators at admission and recurrence were compared, which corresponded to only 9 of the 19 pregnant women with recurrence and 2 of the 6 non-pregnant individuals with recurrence. With this comparison, it was not possible to evidence any statistically significant difference in the expression of immune mediators between those who had homologous and heterologous recurrences (data not shown).
DISCUSSION
This study compared P. vivax genotypes between the primary episode and the recurrence in the pregnant and non-pregnant population, and also explored the association between those parasite genotypes with the expression of proinflammatory and anti-inflammatory immune mediators. The findings showed that vivax malaria recurrences are considerably more frequent in pregnant women than in non-pregnant individuals, 46% versus 13%, respectively. In pregnant women, there was greater parasite genetic diversity, both in the primary episode and in the recurrent one, as well as a predominance of recurrences heterologous to the primary episode. Likewise, pregnant women showed higher levels of proinflammatory and anti-inflammatory immune mediators in the primary episode, and a significant decrease of Fox-P3 in the recurrence; while in non-pregnant individuals, immune mediators remained relatively stable between the initial episode and the recurrence.
These findings confirm the high genetic diversity of P. vivax in Colombia, both in the pregnant and non-pregnant population 39^,^42^-^46. Although the predominant alleles in each microsatellite studied were not exclusive to pregnant women, predominant alleles exclusive to the primary episode or the recurrence were found in pregnant women, which is consistent with previous Colombian studies in non-pregnant individuals, which show parasite haplotypes exclusive to recurrences 29.
In both cohorts, polyclonal infections were prevalent in primary episodes and recurrences, which coincides with previous Colombian studies in pregnant women 44^,^46 and non-pregnant individuals 28^,^44^,^45, and with international studies that highlight that P. vivax infections have high genetic complexity [25]. Pregnant women showed a higher frequency of polyclonal infections in the recurrence than in the initial episode, while non-pregnant individuals showed greater polyclonality in the primary episode than in the recurrence. This could be explained because pregnant women have a higher probability of presenting multiclonal recurrences due to not receiving primaquine, which acts on hypnozoites.
The frequency of relapses/reinfections detected in pregnant women was very high (41%, 17/41), even higher than reported in other Colombian studies: 29% with 120-day follow-up 33 and 18% to 24% with 180-day follow-up 28^,^47. The use of qPCR in addition to microscopy for infection detection in this study may explain that higher frequency of recurrences.
Heterologous recurrences were much more frequent than homologous ones, both in pregnant women and in non-pregnant individuals, which is consistent with the high number of polyclonal infections detected in both cohorts and coincides with other studies reporting that heterologous recurrences are generally polyclonal and occur in most endemic areas 40^,^48. These findings also agree with another Colombian study that included P. vivax patients resident in Turbo (endemic municipality) and Medellín (non-endemic municipality) and reported that isolates from recurrent infections were genetically different from isolates of the initial infection 29. But they contrast with another work carried out in Turbo that reported that 93% of recurrent infections by P. vivax were homologous 28.
To our knowledge, this is the first work exploring the association between the host immune profile with the parasite genotype in primary and recurrent episodes, both in pregnant and non-pregnant individuals in Colombia. No statistically significant differences were found in the level of expression of immune mediators between initial and recurrent infection, nor between homologous and heterologous recurrences. Therefore, there was no association of the host immune profile with plasmodial genotypes in initial and recurrent infection; but, perhaps, this is due to the small sample size available for analysis, as there are studies reporting the existence of differences between levels of immune mediators in recurrence, depending on whether it is genetically homologous or heterologous to the initial episode 21^,^22.
As a strength, the multifaceted approach to the problem in pregnant women is highlighted, having a cohort of non-pregnant individuals as a comparison group. Similarly, limitations such as the relatively small sample size, the low quantity of molecular markers used, and the lack of knowledge of the nutritional status of participants and their infection status with other parasites such as intestinal ones, which are conditions that can modulate the immune response, are accepted. However, these limitations do not invalidate the findings of the study, since comparisons are made between patients from the same region, who are exposed to similar socioeconomic conditions, generally precarious as in almost all malaria-endemic areas of the world, so it is expected that their nutritional status and infection with intestinal parasites would also be similar.
In conclusion, pregnant women are exposed to a high frequency of recurrences with high genetic variation, which may affect the development of effective protective immunity and generate adverse maternal-neonatal outcomes. This raises the need not only to strengthen actions for prevention of P. vivax malaria in pregnant women, but also to optimize antimalarial treatment and find alternatives to the hypnozoiticidal regimen currently available in almost all malaria-endemic areas.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1López-Guzmán C Carmona-Fonseca J Submicroscopic placental malaria histopathology and expression of physiological process mediators Rev Peru Med Exp Salud Publica 20203722022810.17843/rpmesp.2020.372.475932876209 · doi ↗ · pubmed ↗
- 2Bôtto-Menezes C Silva Dos Santos MC Lopes Simplício J Menezes de Medeiros J Barroso Gomes KC de Carvalho Costa IC Plasmodium vivax Malaria in Pregnant Women in the Brazilian Amazon and the Risk Factors Associated with Prematurity and Low Birth Weight A Descriptive Study P Lo S One 201510 e 014439910.1371/journal.pone.014439926675007 PMC 4687654 · doi ↗ · pubmed ↗
- 3Agudelo OM Aristizabal BH Yanow SK Arango E Carmona-Fonseca J Maestre A Submicroscopic infection of placenta by Plasmodium produces Th 1/Th 2 cytokine imbalance, inflammation and hypoxia in women from north-west Colombia Malar J 20141312212210.1186/1475-2875-13-12224673747 PMC 3972514 · doi ↗ · pubmed ↗
- 4BardajíA Martínez-Espinosa FE Arévalo-Herrera M Padilla N Kochar S Ome-Kaius M Burden and impact of Plasmodium vivax in pregnancy A multi-centre prospective observational study P Lo S Negl Trop Dis 201711 e 000560610.1371/journal.pntd.000560628604825 PMC 5481034 · doi ↗ · pubmed ↗
- 5Global Malaria Programme (GMP) World malaria report 2024. Addressing inequity in the global malaria response World Health Organization 2024
- 6Cardona-Arias JA Higuita-Gutiérrez LF Carmona-Fonseca J Clinical and Parasitological Profiles of Gestational, Placental and Congenital Malaria in Northwestern Colombia Trop Med Infect Dis 2023829229210.3390/tropicalmed 806029237368710 PMC 10302000 · doi ↗ · pubmed ↗
- 7Carmona-Fonseca J Arango E Maestre A Placental malaria in Colombia histopathologic findings in Plasmodium vivax and P. falciparum infections Am J Trop Med Hyg 2013881093110110.4269/ajtmh.12-036323546807 PMC 3752808 · doi ↗ · pubmed ↗
- 8Carmona-Fonseca J Agudelo-García O Arango-Flórez E Eficacia terapéutica y eventos adversos de tratamientos para malaria vivax y malaria falciparum en gestantes en las regiones de Urabá y Alto San Jorge, Colombia, 2008-2011 Rev Colomb Obstet Ginecol 2013642737