Unveiling Valuable Secondary Metabolites from the Bioconversion of Banana (Musa balbisiana) Peel-Derived Biomass with Aspergillus niger. Metabolomic Insights into the Chemical Profiles
Jhuly Wellen Ferreira Lacerda, Giovanna Amaral Filipe, Lucas Pradi, Tatiane de Andrade Maranhão, Diogo Robl, Louis Pergaud Sandjo

TL;DR
This study explores how Aspergillus niger can convert banana peels into valuable secondary metabolites, offering a low-cost way to repurpose agricultural waste.
Contribution
The study identifies optimal conditions and key metabolites from fungal bioconversion of banana peel biomass using metabolomic analysis.
Findings
BP media yielded 76 secondary metabolites, 66 of which were produced by A. niger.
BPS media produced 99 metabolites, including 22 previously identified A. niger-derived compounds.
21-day BP and 28-day BPS fermentations were optimal for secondary metabolite production.
Abstract
Santa Catarina is Brazil’s fourth-largest banana producer, generating a significant amount of waste. Banana peel conversion into useful products is uncommon. However, it produces natural products that possess benefits for human health or can serve as biomass to produce fungi-derived metabolites. Therefore, Aspergillus niger was isolated from banana peels and then cultured on three distinct media composed of starch, banana peels (BP), and starch-enriched banana peels (BPS) for 14, 21, and 28 days at 28 °C. Their chemical profiles were established using liquid chromatography coupled to mass spectrometry data assisted by the GNPS database and MS-FINDER software. In BP media, 76 secondary metabolites were detected; among them, 66 were produced by the fungus. Twenty-seven compounds, such as nigragillin and pyranonigrin A, are A. niger chemomarkers, while other substances, except for the 13…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1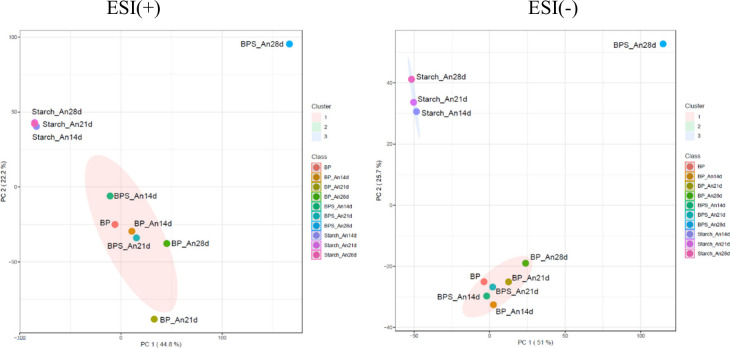 2
2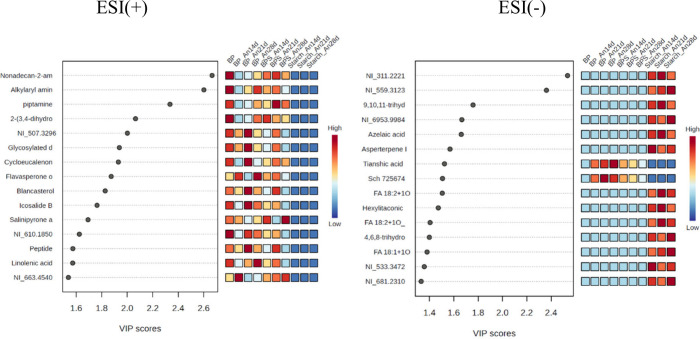 3
3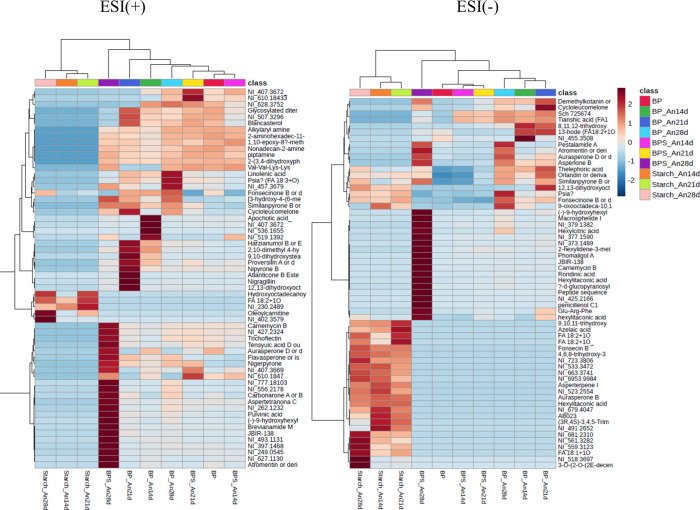 4
4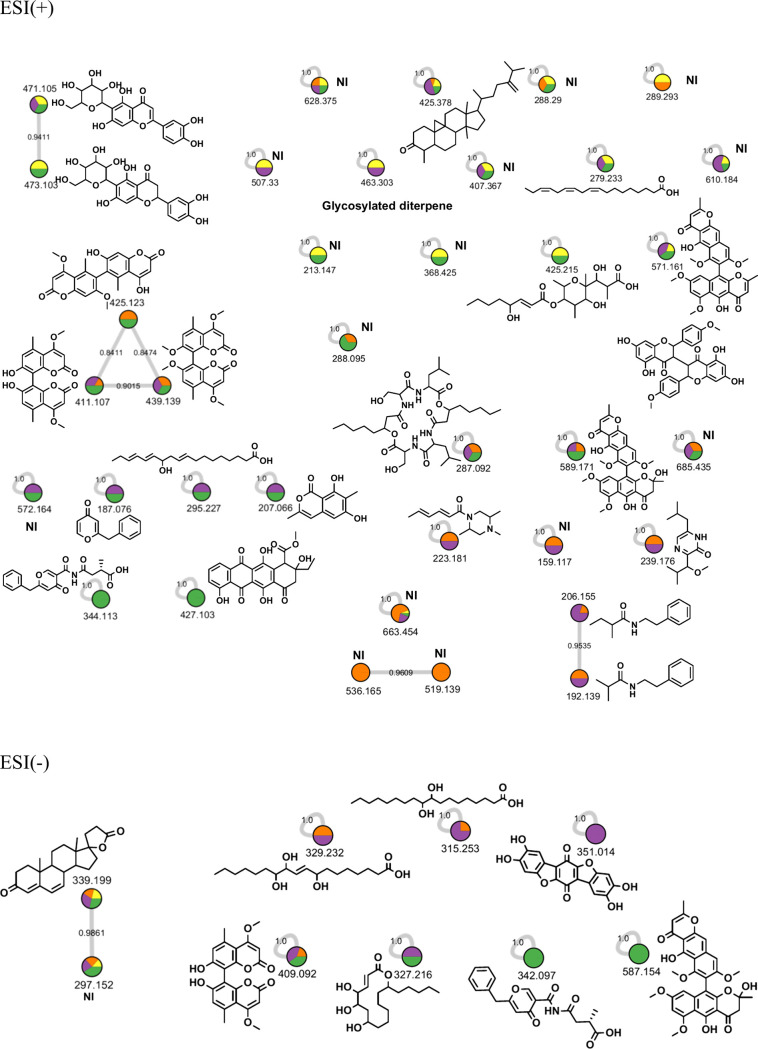 5
5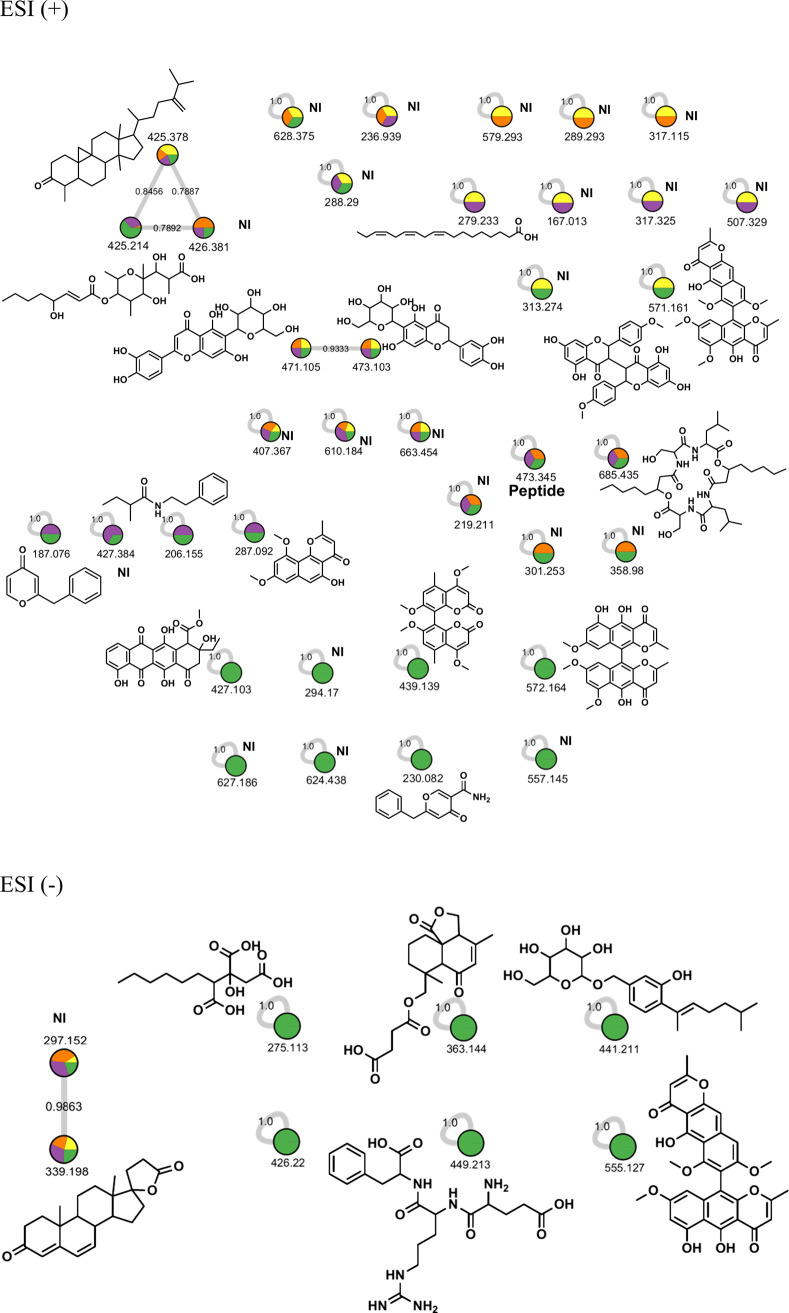 6
6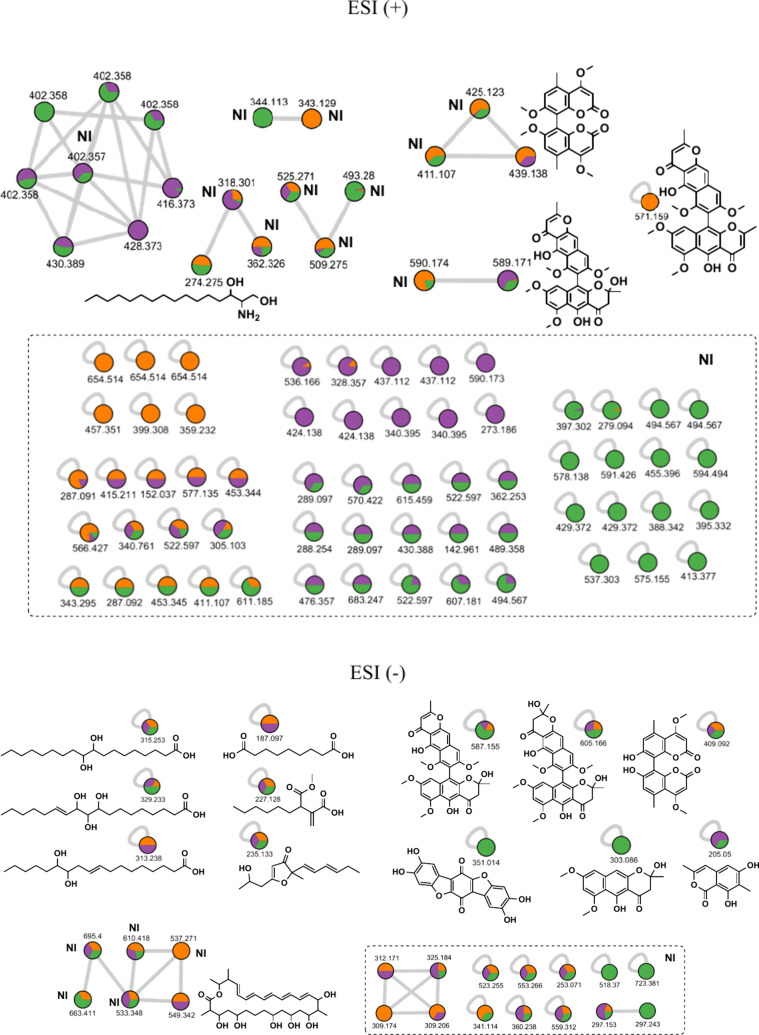 7
7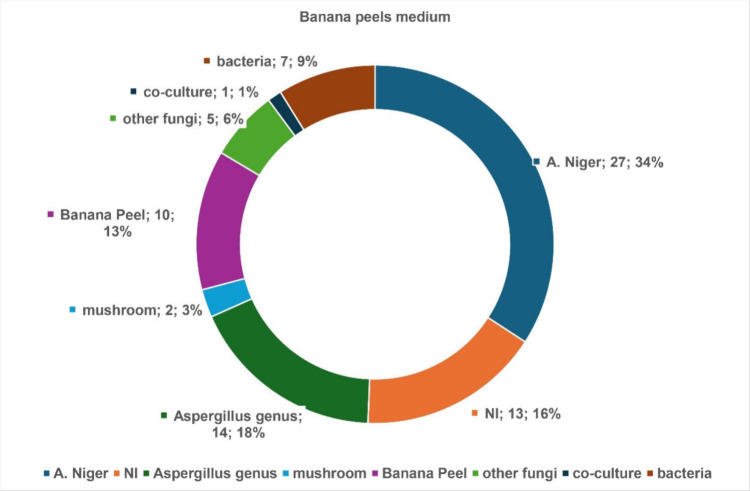 8
8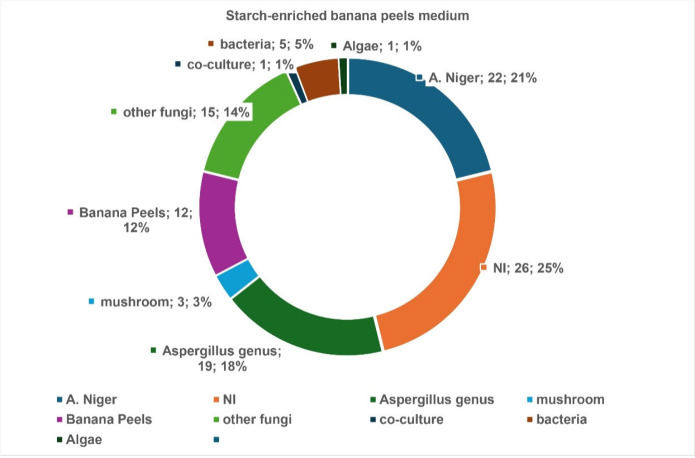 9
9| no. |
| Formula | Adduct type | Precursor | Error (ppm) | Proposed identification | Score | Class | Time (day) | Specie | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.76 | C13H22N2O | [M + H]+ | 223.1811 | –2.30 | Nigragillin isomer | 5.90 | pyrazine | 21 | NI | |
| 2 | 0.77 | C10H22O6 | [M + Na]+ | 261.1316 | –0.03 | NI | - | polyol | BP | NI | - |
| 3 | 1.63 | C13H22N2O | [M + H]+ | 223.1811 | –0.95 | Nigragillin | 5.92 | piperazine | 14, 21 |
|
|
| 4 | 2.21 | C10H9NO5 | [M + H]+ | 224.0557 | –1.57 | Pyranonigrin A | 6.26 | Pyranopyrrole | 28 |
|
|
| [M + H]− | 222.0399 | –1.56 | |||||||||
| 5 | 2.67 | C13H22N2O2 | [M + H]+ | 239.1758 | –1.66 | Ochramide B | <5 | Pyrazine | 14, 21 |
|
|
| 6 | 2.80 | C8H8O3 | [M + H]+ | 153.0553 | –4.47 | Phenoxyacetic acid or derivatives | 5.58 | Phenoxyacetic acid derivative | 28 |
|
|
| 7 | 4.98 | C18H10O7 | [M + H]+ | 339.0503 | –1.10 | Cycloleucomelone | 6.19 | Benzofurans | 21, 28 |
|
|
| [M-H]− | 337.0338 | 4.66 | |||||||||
| 8 | 5.04 | C18H12O6 | [M-H]− | 323.0550 | 3.43 | Atromentin or derivatives | 7.14 | P-benzoquinones | 28 |
|
|
| 9 | 5.04 | C13H11NO3 | [M + H]+ | 230.0819 | –2.75 | Carbonarone A or B | 6.21 | Pyridine carboxaldehyde | 21, 28 |
|
|
| 10 | 5.05 | C11H10O4 | [M + H]+ | 207.0657 | –2.50 | Similanpyrone B | 6.78 | Isocoumarins | 14, 21, 28 |
|
|
| [M-H]− | 205.0496 | 3.08 | 7.06 | ||||||||
| 11 | 5.05 | C16H17NO4 | [M + Na]+ | 310.1057 | –2.51 | Pyrophen | <5 | Kavalactone | 14, 21, 28 |
|
|
| 12 | 5.06 | C12H10O2 | [M + H]+ | 187.0759 | –2.92 | Nigerpyrone | - | Polyketide | 14, 21, 28 |
|
|
| 13 | 5.24 | C12H20O7 | [M-H]− | 275.1126 | –1.73 | Hexylcitric acid | - | alkylcitric acids | 28 |
|
|
| 14 | 5.30 | C22H18O8 | [M + H]+ | 411.1079 | –1.112.17 | Orlandin or derivatives | 7.28 | bicoumarin | 14, 21, 28 |
|
|
| [M-H]− | 409.0920 | 2.17 | |||||||||
| 15 | 5.47 | C18H8O8 | [M-H]− | 351.0136 | 2.96 | Thelephoric acid | 5.52 | Terphenyl | 14, 21, 28 | mushroom |
|
| 16 | 5.48 | C13H20O2 | [M + H]+ | 209.1541 | –2.37 | 2,10-dimethyl 4-hydroxy-6-oxo-4-undecen-7-yne | 5.44 | Enone | 14, 21 |
|
|
| 17 | 5.58 | C12H17NO | [M + H]+ | 192.1386 | –1.62 | 2-Methyl-N-(2-phenylethyl)propanamide | 8.01 | Alkamide | 14, 21 | NI | - |
| 18 | 5.60 | C13H22O2 | [M + H]+ | 211.1697 | –2.11 | Harzianumol B or E | <5 | Fatty alcohols | 14, 21 |
|
|
| 19 | 6.07 | C22H18O9 | [M + H]+ | 427.1027 | –0.80 | Maggiemycin | 7.23 | Tetracenequinones | 28 |
|
|
| 20 | 6.15 | C20H12O8 | [M + H]+ | 381.0604 | 0.25 | Asperlone B | 5.405.79 | Dinaphthalenone | 14, 21, 28 |
|
|
| [M-H]− | 379.0450 | 2.48 | |||||||||
| 21 | 6.316.33 | C18H17NO6 | [M + H]+ | 344.1132 | –0.98 | Pestalamide A | 7.216.88 | Pyridone | 28 |
|
|
| [M-H]− | 342.0970 | 3.82 | |||||||||
| 22 | 6.31 | C13H19NO | [M + H]+ | 206.1546 | –3.21 | 2-Methyl-N-(2-phenylethyl)butanamide | 6.84 | Alkamide | 14, 21 | Bacteria |
|
| 23 | 6.33 | C23H20O8 | [M + H]+ | 425.1232 | –0.25 | Isokotanin B | 6.59 | bicoumarin | 14, 28 |
|
|
| [M-H]− | 423.1071 | –2.11 | |||||||||
| 24 | 6.33 | C13H11NO3 | [M-H]− | 228.0656 | 4.44 | Carbonarone A or B | <5 | Pyranones and derivative | 28 |
|
|
| 25 | 6.50 | C23H14O9 | [M + H]+ | 435.0708 | 0.60 | [3-hydroxy-4-(6-methoxy-5,8-dioxonaphthalen-2-yl)-5-oxofuran-2-ylidene](4-hydroxyphenyl)acetic acid | 6.55 | Naphthoquinones | 21, 28 | mushroom |
|
| 26 | 6.62 | C18H32O5 | [M + Na]+ | 351.2143 | –1.26 | Sch725674 | 6.60 | Macrolide | 14, 21, 28 |
|
|
| [M-H]− | 327.2163 | 4.26 | |||||||||
| 27 | 6.64 | C18H30O3 | [M+H+ | 293.2116 | –0.24 | PsiAalfa | - | Lacton | 14, 21, 28 |
|
|
| 28 | 6.74 | C27H24N2O5 | [M + H]+ | 457.1759 | –0.22 | Aspernigrin B | 6.90 | Lactam | 14, 21 |
|
|
| 29 | 6.83 | C11H18O4 | [M-H]− | 213.1126 | 2.95 | hexylitaconic acid | 7.93 | Fatty acid | 21, 28 |
|
|
| 30 | 7.06 | C18H34O5 | [M + Na]+ | 353.2297 | –1.96 | 8,11,12-trihydroxyoctadec-9-enoic acid derivative | - | Fatty acid | 21, 28 |
|
|
| 31 | 7.06 | C14H20O3 | [M + Na]+ | 259.1312 | 0.72 | Asperfuranone A | - | polyketide | 14, 21 |
|
|
| 32 | 7.29 | C18H32O3 | [M + H]+ | 297.2428 | –1.28 | PsiAβ | 5.5 | lacton | 21 |
|
|
| 33 | 7.29 | C18H34O4 | [M + Na]+ | 337.2350 | –1.423.92 | 12,13-dihydroxyoctadec-9-enoic acid derivative | - | Fatty acid | 21 |
|
|
| [M-H]− | 313.2372 | 3.92 | |||||||||
|
| 7.35 | C31H26O11 | [M + H]+ | 575.1550 | –0.37 | Isoaurasperone F or Nigerasperone C | 6.16 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 35 | 7.53 | C14H25NO3 | [M + H]+ | 256.1912 | –1.88 | NI | 5.91 | Alkaloid | 21 | NI | - |
| 36 | 7.57 | C13H18O | [M + H]+ | 191.1437 | –3.46 | β-Damascenone | 5.05 | Enone | 21, 28 |
|
|
| 37 | 7.57 | C24H22O8 | [M + H]+ | 439.1388 | –0.13 | Kotanin | 7.29 | Bicoumarin | 14, 21, 28 |
|
|
| 38 | 7.63 | C24H22O8 | [M + H]+ | 439.1393 | –0.36 | Kotanin isomer | 7.33 | Bicoumarin | 14, 21, 28 |
|
|
| 39 | 7.64 | C18H34O4 | [M-H]− | 313.2373 | 3.61 | 8,11,12-trihydroxyoctadec-9-enoic acid derivative | 6.17 | Fatty acid | 14, 21 |
|
|
| 40 | 7.71 | C31H24O10 | [M-H]− | 555.1280 | 3.0 | Aurasperone D or derivative | - | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 41 | 7.80 | C15H22O3 | [M + Na]+ | 273.1468 | 0.50 | Proversilin A | - | Sesquiterpene | 14, 21 |
|
|
| 42 | 7.80 | C16H14O5 | [M + H]+ | 287.0920 | –2.10 | Flavasperone | 7.25 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 43 | 7.98 | C18H36O4 | [M + Na]+ | 339.2511 | 1.64 | 9,10-dihydroxystearic acid derivative | 7.55 | Fatty acid | 14, 21 |
|
|
| [M-H]− | 315.2531 | 3.11 | 7.21 | ||||||||
| 44 | 8.00 | C18H30O | [M + H]+ | 263.2376 | –2.51 | Norditerpene | - | Norditerpene | 14, 21 |
| – |
| 45 | 8.00 | C46H64O2 | [M + Na]+ | 671.4774 | –4.47 | Menaquinone 7 | - | Meroterpenoid | 21 |
|
|
| 46 | 8.00 | C18H34O3 | [M + H]+ | 299.2588 | –2.44 | 6-keto stearic acid derivative | 5.21 | Fatty acid | 14, 21 |
|
|
| 47 | 8.13 | C16H14O5 | [M + H]+ | 287.0922 | –2.80 | Flavasperone isomer | 7.79 | Naphtho-γ-pyrone | 14 |
|
|
| 48 | 8.20 | C32H28O11 | [M + H]+ | 589.1706 | –0.28 | Fonsecinone B | 6.36 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 49 | 8.20 | C31H24O10 | [M + Na]+ | 579.1260 | 0.30 | Aurasperone | 5.16 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| [M-H]− | 555.1282 | –1.66 | |||||||||
| 50 | 8.27 | C14H22O3 | [M + Na]+ | 261.1464 | –1.20 | Nipyrone B | 5.14 | α-pyrone derivative | 21 |
|
|
| 51 | 8.27 | C32H28O11 | [M + H]+ | 589.1703 | 0.24 | Fonsecinone B isomers | 6.06.63 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| [M-H]− | 587.1542 | 2.87 | |||||||||
| 52 | 8.86 | C32H26O10 | [M + H]+ | 571.1605 | –1.10 | Aurasperone A | 5.99 | Naphtho-γ-pyrone | 21, 28 |
|
|
| 53 | 9.10 | C18H30O3 | [M+H–H2O]+ | 277.2167 | –1.684.14 | Psiaα | 6.52 | Fatty alcohol | 14, 21, 28 |
|
|
| [M-H]− | 293.2110 | 4.14 | |||||||||
| 54 | 9.34 | C32H26O10 | [M + H]+ | 571.1608 | –1.98 | Aurasperone A isomer | 5.6 | Naphtho-γ-pyrone | 21, 28 |
|
|
| 55 | 9.35 | C18H28O3 | [M-H]− | 291.1952 | 4.68 | 9-oxooctadeca-10,12,15-trienoic acid derivative | 5.17 | Fatty acid | 14, 28 |
|
|
| 56 | 9.74 | C18H30O2 | [M + H]+ | 279.2324 | –1.95 | Linolenic acid | 7.39 | Fatty acid | 14, 21, 28 |
|
|
| 57 | 10.23 | C23H41N | [M + H]+ | 332.3312 | –0.07 | Piptamine derivative | 8.15 | alkylamine | BP, 14, 21, 28 | NI | – |
| 58 | 10.24 | C19H41N | [M + H]+ | 284.3316 | –1.49 | Nonadecan-2-amine derivative | 6.51 | alkylamine | BP, 14, 21, 28 | NI | – |
| 59 | 10.38 | C30H48O3 | [M + H]+ | 457.3679 | –0.61 | Triterpene |
| Triterpenoid | 28 | NI | – |
| 60 | 10.50 | C30H48O | [M + H]+ | 425.3776 | 0.45 | 4α,14α-dimethyl-24-methylene-cholest-7,9(11)-dien-3β-ol | 7.15 | Triterpene | 14 | NI | – |
| 61 | 10.90 | C17H24O4 | [M + H]+ | 315.1574 | 0.54 | Salinipyrone A | 5.40 | polyketides | 14, 28 | Bacteria |
|
| 62 | 11.29 | C20H34O8 | [M + H]+ | 403.2326 | 0.11 | Botcinic acid derivative | 7.12 | Polyketide | 28 |
|
|
| [M + Na]+ | 425.2150 | ||||||||||
| 63 | 11.87 | C18H30O2 | [M + H]+ | 279.2327 | –1.23 | Linolenic acid derivative | 6.89 | Fatty acid | BP, 21, 28 |
|
|
| 64 | 12.28 | C25H20O11 | [M + Na]+ | 471.1050 | –1.25 | Isoorientin | - | Flavonoid | BP, 14, 21, 28 |
|
|
| 65 | 12.28 | C21H22O11 | [M + Na]+ | 473.1027 | –1.3 | Eriodictyol 6-C-β- | - | Flavonoid | BP, 14, 21, 28 |
|
|
| 66 | 14.38 | C30H48O3 | [M + H]+ | 457.3662 | 3.33 | Triterpene | 5.05 | Triterpenoid | 14, 21 | NI | – |
| [M-H]− | 455.3508 | –3.78 | |||||||||
| 67 | 14.44 | C33H54O5 | [M + H]+ | 531.4075 | 4.80 | Related to Atlanticone B Esters | - | sesquitepene | 21 |
|
|
| 68 | 14.44 | C35H71NO3 | [M + H]+ | 554.5506 | 0.13 |
| 5.82 | ceramide | 21 |
|
|
| 69 | 14.96 | C24H38O4 | [M + H]+ | 391.2849 | –1.57–0.43 | Apocholic acid | 5.64 | Steroid | 14 | NI | – |
| [M + Na]+ | 413.2664 | –0.43 | |||||||||
| 70 | 15.12 | C22H43NO | [M + H]+ | 338.3422 | –1.34 | Erucamide | 5.84 | Alkamide | 28 |
|
|
| 71 | 15.12 | C30H48O | [M + H]+ | 425.3781 | –0.49 | 4α,14α-dimethyl-24-methylene-cholest-7,9(11)-dien-3β-ol isomer | 7.67 | Triterpenoid | 28 |
| – |
| 72 | 15.18 | C24H38O4 | [M + H]+ | 391.2848 | –0.09 | di-(2-ethylhexyl)phthalate | 5.39 | Phthalate | BP, 14 |
|
|
| 73 | 15.29 | C34H60N4O10 | [M + H]+ | 685.4361 | 3.10 | Icosalide B | 6.45 | Cyclic depsipeptide | BP, 21, 28 |
|
|
| 74 | 15.29 | C22H43NO | [M + H]+ | 338.3421 | –1.06 | Erucamide isomer | 5.28 | Alkamide | 14 |
|
|
| 75 | 15.29 | C24H38O4 | [M + Na]+ | 413.2653 | –3.95 | di-(2-ethylhexyl)phathalate isomer | - | Phthalate | BP, 21 |
|
|
| 76 | 15.29 | C30H48O | [M + H]+ | 425.3781 | –0.49 | Cycloeucalenone | 8.01 | Triterpenoid | BP, 21 |
|
|
| no. |
| Formula | Adduct type | Precursor | Mass error | Metabolite name | Score | Class | Time (day) | Species | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.53 | C14H19NO5 | [M + H]+ | 282.1344 | –2.85 | Harzianopyridone | 6.44 | pyridone | 28 |
|
|
| 2 | 0.73 | C16H23NO5 | [M + H]+ | 310.1651 | –0.65 | NI | – | alkaloid | 28 | NI | – |
| 3 | 0.77 | C10H22O6 | [M + Na]+ | 261.1316 | –0.03 | NI | – | polyol | BP | NI | – |
| 4 | 2.23 | C10H9NO5 | [M + H]+ | 224.0561 | –3.37 | Pyranonigrin A | 6.26 | Pyranones and derivative | 28 |
|
|
| [M-H]− | 222.0398 | 4.47 | 5.23 | ||||||||
| 5 | 3.68 | C11H16O6 | [M-H]− | 243.0867 | 2.92 | Tensyuic acid A or F | 6.6 | Fatty diacid | 28 |
|
|
| 6 | 3.76 | C11H18O5 | [M + Na]+ | 253.1052 | –2.41 | 9-hydroxyhexylitaconic acid | 7.47 | Fatty diacid | 28 |
|
|
| [M-H]− | 229.1070 | 2.30 | |||||||||
| 7 | 3.77 | C11H16O4 | [M + H]+ | 213.1128 | –3.13 | Aspergilactone A | 6.29 | Butenolide | 28 |
|
|
| 8 | 4.43 | C16H25NO4 | [M-H]− | 294.1698 | 4.34 | Penicillenol C1 | 5.0 |
| 28 |
|
|
| 9 | 4.56 | C16H23NO4 | [M + H]+ | 294.1708 | –2.44 | NI | – | Alkaloid | 28 | NI | – |
| 10 | 4.60 | C22H16O9 | [M + H]+ | 425.0866 | 0.26 | 1,2-diacetoxy-3-(4′-hydroxyphenyl)-4,7,8-trihydroxydibenzofuran | 5.81 | Terphenyl derivative | 28 | mushroom |
|
| 11 | 4.60 | C21H20O10 | [M + Na]+ | 455.0969 | 3.26 | purpurquinone A | – | polyketide | 28 |
|
|
| 12 | 4.864.95 | C14H22O7 | [M + Na]+ | 325.1264 | –2.07 | Roridinic acid | 5.58 | Fatty acid | 28 |
|
|
| [M-H]− | 301.1281 | 3.89 | |||||||||
| 13 | 4.86 | C26H38N6O12 | [M-H]− | 625.2447 | –3.59 | Peptide | – | Peptide | 28 |
|
|
| 14 | 4.95 | C18H15N3O3 | [M + H]+ | 322.1180 | 1.92 | Brevianamide M | 5.19 | Quinazoline | 28 |
|
|
| 15 | 5.04 | C18H12O6 | [M-H]− | 323.0550 | 3.43 | Atromentin | 7.38 | P-benzoquinone | 28 |
|
|
| 16 | 5.06 | C12H10O2 | [M + H]+ | 187.0762 | –4.54 | Nigerpyrone | – | polyketide | 21, 28 |
|
|
| 17 | 5.09 | C11H10O4 | [M + H]+ | 207.0657 | –0.16 | Similanpyrone B or derivatives | 6.95 | Isocoumarins | 21,28 |
|
|
| [M-H]− | 205.0501 | 3.07 | |||||||||
| 18 | 5.09 | C14H20O6 | [M+HCO2]− | 329.1231 | –1.65 | Phomaligol A | – | polyketide | 28 |
|
|
| 19 | 5.11 | C13H11NO3 | [M + H]+ | 230.0820 | –3.62 | Carbonarone A or B | 6.99 | g-pyrone | 28 |
|
|
| 20 | 5.11 | C16H8O3 | [M + H]+ | 249.0545 | 0.49 | NI | – | Polyketide | 28 | NI | – |
| 21 | 5.11 | C16H17NO4 | [M + Na]+ | 310.1048 | –2.35 | Pyrophen | Pyrone | 21, 28 |
|
| |
| 22 | 5.11 | C18H12O6 | [M + H]+ | 325.0709 | –0.96 | Atromentin derivatives | – | P-benzoquinones | 28 |
|
|
| 23 | 5.11 | C15H22O8 | [M + Na]+ | 353.1208 | –1.24 | NI | – | polyketide | 28 | NI | – |
| 24 | 5.11 | C31H24O13 | [M + Na]+ | 627.1130 | 2.46 | NI | – | Polyketide | 28 | NI | – |
| 25 | 5.21 | C12H20O7 | [M-H]− | 275.1128 | 2.99 | Hexylcitric acid | – | Acids | 28 |
|
|
| 26 | 5.30 | C22H18O8 | [M + H]+ | 411.1076 | –0.38 | Orlandin or isomer | 5.365.96 | coumarin | 21, 28 |
|
|
| [M-H]− | 409.0917 | 2.90 | |||||||||
| 27 | 5.36 | C19H24O8 | [M-H]− | 379.1382 | –2.88 | NI | – | polyketide | 28 | NI | – |
| 28 | 5.48 | C20H26O8 | [M-H]− | 393.1539 | 4.04 | Hamigeromycin A or C | 7.20 | Zearalenone | 28 | Fungus |
|
| 29 | 5.54 | C16H24O8 | [M + Na]+ | 367.1366 | –0.76 | Macrosphelide I | 7.07 | macrolide | 28 | Fungus |
|
| [M-H]− | 343.1387 | 3.32 | |||||||||
| 30 | 5.58 | C12H17NO | [M + H]+ | 192.1386 | –1.62 | 2-Methyl-N-(2-phenylethyl)propanamide | 8.08 | N-acyl amine | 21 | NI | – |
| 31 | 5.64 | C17H26O9 | [M + Na]+ | 397.1468 | –1.644.03 | NI | – | NI | 28 | NI | – |
| [M-H]− | 373.1489 | ||||||||||
| 32 | 5.78 | C18H15NO | [M + H]+ | 262.1232 | –2.14 | NI | NI | 28 | NI | – | |
| 33 | 5.78 | C18H12O5 | [M + H]+ | 309.0762 | –1.46 | Pulvinic acid | 5.61 | Pulvinone | 28 |
|
|
| 34 | 5.78 | C18H12O7 | [M + H]+ | 341.0660 | –0.94 | Atromentic acid | – | Pulvinone | 28 |
|
|
| 35 | 6.05 | C22H18O9 | [M + H]+ | 427.1028 | –1.04 | Maggiemycin | 7.23 | Tetracenequinones | 28 |
|
|
| 36 | 6.05 | C24H20O8 | [M + H]+ | 437.1229 | 0.45 | inoscavin B | – | Styrylpyrone | 28 | Mushroom |
|
| 37 | 6.07 | C21H32O7 | [M-HCO2]− | 441.2113 | –2.62 | β- | – | Sesquiterpene | 28 |
|
|
| 38 | 6.19 | C20H12O8 | [M-H]− | 379.0445 | 3.79 | Asperlone B | 5.93 | Dinaphthalenone | 28 |
|
|
| 39 | 6.29 | C17H17NO4 | [M + Na]+ | 322.1054 | –1.41 | NI | – | Alkaloid | 28 | NI | – |
| 40 | 6.296.54 | C19H24O7 | [M + Na]+ | 387.1415 | –0.213.09 | JBIR-138 | 6.80 | Sesquiterpene esters | 28 |
|
|
| [M-H]− | 363.1438 | ||||||||||
| 41 | 6.29 | C22H28O8 | [M + H]+ | 421.1856 | 0.46 | Aspertetranone C | 5.11 | Naphthopyranones | 28 |
|
|
| 42 | 6.29 | C27H40O10 | [M + H]+ | 525.2720 | –4.91 | NI | – | – | 28 | NI | – |
| 43 | 6.31 | C13H19NO | [M + H]+ | 206.1547 | –3.70 | 2-Methyl-N-(2-phenylethyl)butanamide | 6.80 | Alkamide | 21, 28 |
|
|
| 44 | 6.33 | C23H20O8 | [M + H]+ | 425.1232 | –0.25 | Demethylkotanin | 6.59 | bicoumarin | 21, 28 |
|
|
| [M-H]− | 423.1071 | –2.11 | |||||||||
| 45 | 6.38 | C13H11NO3 | [M-H]− | 228.0661 | 2.26 | Carbonarone A or B | 5.29 | g-pyrone | 28 |
|
|
| 46 | 6.38 | C18H17NO6 | [M-H]− | 342.0973 | 2.95 | Pestalamide A | 6.58 | Pyranone | 28 |
|
|
| 47 | 6.50 | C23H14O9 | [M + H]+ | 435.0708 | 0.60 | [3-hydroxy-4-(6-methoxy-5,8-dioxonaphthalen-2-yl)-5-oxofuran-2-ylidene](4-hydroxyphenyl)acetic acid | 6.55 | pulvinone | 28 | mushroom |
|
| 48 | 6.58 | C19H24O5 | [M-HCO2]− | 377.1590 | 4.17 | Asperfuranone | – | Polyketide | 28 |
|
|
| 49 | 6.61 | C18H32O5 | [M + Na]+ | 351.2143 | –1.26 | Sch725674 | – | Macrolide | 14, 21, 28 |
|
|
| 50 | 6.61 | C18H28O3 | [M + H]+ | 293.2114 | –0.90 | NI | – | NI | 14, 21, 28 | NI | – |
| 51 | 6.68 | C13H20O6 | [M + Na]+ | 295.1158 | –2.17 | Tensyuic acid D or C | <5 | Fatty diacid | 28 |
|
|
| 52 | 6.68 | C14H20O6 | [M + Na]+ | 307.1158 | 0.14 | Pyrenocine L | – | Pyrone | 28 | Fungus |
|
| 53 | 6.68 | C26H34N4O7 | [M + H]+ | 515.2485 | 2.96 | Related to tyr-tryp-val-Ala | – | Peptide | 28 | NI | – |
| 54 | 6.70 | C12H20O4 | [M-H]− | 227.1280 | 3.87 | 2-hexylidene-3-methylsuccinic acid 4-methyl ester | 5.76 | Fatty diacid | 28 | Fungus |
|
| 55 | 6.81 | C20H30N6O6 | [M-H]− | 449.2135 | 2.33 | Related to Glu-Arg-Phe | – | Peptide | 28 | NI | – |
| 56 | 6.83 | C11H18O4 | [M + Na]+ | 237.1102 | –0.33 | Hexylitaconic acid | 7.98 | Fatty diacid | 28 |
|
|
| [M-H]− | 213.1127 | 2.49 | |||||||||
| 57 | 6.95 | C17H24O7 | [M + H]+ | 341.1616 | –3.25 | Aspergilloid E | – | Sesquiterpene | 28 |
|
|
| 58 | 6.95 | C21H16O9 | [M + Na]+ | 435.0713 | 4.82 | Mitorubrinic acid | – | Azaphilone | 28 |
|
|
| 59 | 7.08 | C18H32O4 | [M+H–H2O]+ | 295.2270 | –0.73 | 10,11-dihydroxyoctadeca-8,12-dienoic acid | 5.33 | Fatty acid | 14, 21 |
|
|
| 60 | 7.08 | C18H34O5 | [M + Na]+ | 353.2298 | –1.68 | 8,11,12-trihydroxyoctadec-9-enoic acid derivative | – | Fatty acid | 14, 21 |
|
|
| 61 | 7.31 | C33H34O13 | [M + H]+ | 639.2077 | –0.75 | NI | – | NI | 28 | NI | – |
| 62 | 7.42 | C32H30N2O8 | [M + H]+ | 571.2076 | –0.19 | Aspernigrin C | 6.69 | Kavalactones | 28 |
|
|
| 63 | 7.49 | C16H35NO2 | [M + H]+ | 274.2744 | –1.26 | Hexadecasphinganine | <5 | Sphingolipids | BP, 14, 21, 28 |
|
|
| 64 | 7.63 | C24H22O8 | [M + H]+ | 439.1391 | –0.81 | Kotanin | 7.13 | bicoumarin | 28 |
|
|
| 65 | 7.68 | C31H24O10 | [M-H]− | 555.1273 | –0.02 | Aurasperone D or derivative | – | Naphtho-γ-pyrone | 28 |
|
|
| 66 | 7.97 | C29H33NO10 | [M + H]+ | 556.2178 | –0.14 | NI | – | NI | 28 | NI | – |
| 67 | 7.80 | C16H14O5 | [M + H]+ | 287.0920 | –2.10 | Flavasperone or isomer | 7.25 | Naphtho-γ-pyrone | 28 |
|
|
| 68 | 8.13 | C16H14O5 | [M + H]+ | 287.0922 | –2.80 | Flavasperone or isomer | 7.79 | Naphtho-γ-pyrone | 21 |
|
|
| 69 | 8.13 | C31H24O10 | [M + H]+ | 557.1451 | –1.58 | Aurasperone D or derivative | 5.8 | Naphtho-γ-pyrone | 21, 28 |
|
|
| 70 | 8.19 | C23H32O9 | [M + H]+ | 453.2122 | –0.64 | Carnemycin B | 5.26 | Polyketides | 28 |
|
|
| [M-H]− | 451.1954 | 4.33 | 5.35 | ||||||||
| 71 | 8.37 | C13H10O5 | [2M+Na]+ | 493.1131 | –0.36 | myxotrichin C | – | Chromone | 28 |
|
|
| 72 | 8.48 | C22H34O8 | [M + H]+ | 427.2324 | 0.57 | Colletobredin C | – | Polyketide | 28 |
|
|
| [M-H]− | 425.2166 | 3.50 | |||||||||
| 72 | 8.48 | C33H60O6 | [M + Na]+ | 575.4262 | –4.45 | NI | – | NI | 28 | NI | – |
| 74 | 8.48 | C39H32O15 | [M + Na]+ | 763.1649 | 1.32 | NI | – | NI | 28 | NI | – |
| 75 | 8.76 | C20H20O6 | [M + H]+ | 357.1336 | –0.94 | Herqueichrysin | – | phenalenone | 28 |
|
|
| 76 | 8.87 | C32H26O10 | [M + H]+ | 571.1599 | –0.046 | Aurasperone A or derivatives | 5.44 | Naphtho-γ-pyrone | 21, 28 |
|
|
| 77 | 8.99 | C40H34O15 | [M + Na]+ | 777.1810 | 1.88 | NI | – | 28 | NI | – | |
| 78 | 9.24 | C21H37N | [M + H]+ | 304.3005 | –2.05 | Alkylaryl amine | 7.61 | Amine | BP, 14, 21, 28 | NI | – |
| 79 | 9.54 | C35H30O11 | [M + H]+ | 627.1865 | –0.66 | Talaroketal A | 6.98 | phenalenone | 28 | Fungus |
|
| 80 | 10.27 | C16H22O4 | [M + H]+ | 279.1597 | –2.21 | 1,10-epoxy-8α-methoxyermophilanolide | – | sesquiterpene | BP, 14, 21, 28 | Plant |
|
| 81 | 10.51 | C30H48O | [M + H]+ | 425.3783 | –1.19 | 4α,14α-dimethyl-24-methylene-cholest-7,9(11)-dien-3β-ol | 7.51 | triterpene | 21, 28 | NI | – |
| 82 | 10.58 | C17H14O5 | [M + H]+ | 299.0920 | –2.01 | Trichoflectin | 6.23 | Azaphilone | 21, 28 | fungus |
|
| 83 | 10.89 | C17H24O4 | [M + Na]+ | 315.1571 | –1.44 | Salinipyrone A | 5.40 | polyketides | 28 | bacteria |
|
| 84 | 11.29 | C20H34O8 | [M + Na]+ | 425.2150 | –1.02 | Botcinic acid | 7.12 | Polyketide | 14, 21, 28 | Fungus |
|
| 85 | 11.29 | C24H48N6O6 | [M + H]+ | 517.3712 | –0.75 | NI | – | Peptide | BP, 14, 21, 28 | NI | – |
| 86 | 11.30 | C22H44N6O5 | [M + H]+ | 473.3453 | 0.20 | Related to Val-Val-Lys-Lys | – | Peptide | BP, 14, 21, 28 | NI | – |
| 87 | 11.86 | C18H30O2 | [M + H]+ | 279.2322 | –1.23 | Linolenic acid | 7.39 | Fatty acid | BP, 21 |
|
|
| 88 | 12.14 | C16H33NO | [M + H]+ | 256.2641 | –1.21 | 2-aminohexadec-11-en-3-ol | – | Sphingolipid | BP, 14, 21, 28 |
|
|
| 89 | 12.32 | C25H20O11 | [M + Na]+ | 471.1051 | –1.03 | isoorientin | – | Flavonoid | BP, 28 |
|
|
| 90 | 12.32 | C21H22O11 | [M + Na]+ | 473.1027 | –1.30 | Eriodictyol 6-C-β- | Flavonoid | BP, 14, 21, 28 |
|
| |
| 91 | 15.09 | C16H26 | [M + H]+ | 219.2113 | –2.62 | NI | – | Sesquiterpene | 21 | NI | – |
| 92 | 15.09 | C21H10N2O4 | [M + H]+ | 355.0702 | 3.20 | 13-Docosenamide | – | Fatty amide | 21 |
|
|
| 93 | 15.09 | C10H16O10 | [2M+H]+ | 593.1578 | 2.16 | veracylglucan A | – | Glycoside | 21 |
|
|
| 94 | 15.11 | C22H43NO | [M + H]+ | 338.3420 | –0.77 | Erucamide | 5.65 | Alkamide | 21 |
|
|
| 95 | 15.12 | C30H48O | [M + H]+ | 425.3781 | –0.49 | 4α,14α-dimethyl-24-methylene-cholest-7,9(11)-dien-3β-ol isomer | 7.67 | Triterpenoid | 21 | NI | – |
| 96 | 15.18 | C21H32O | [M + H]+ | 301.2529 | –1.03 | NI | – | Diterpene | 14, 28 | NI | – |
| 97 | 15.18 | C24H38O4 | [M + H]+ | 391.2848 | –0.09 | di(2-ethylhexyl)phthalate | – | Phthalate | BP, 14, 21, 28 |
|
|
| 98 | 15.18 | C22H43NO | [M + H]+ | 338.3417 | 0.12 | Isomer of erucamide | 5.61 | Alkamide | 14, 28 |
|
|
| 99 | 15.29 | C30H48O | [M + H]+ | 425.3781 | –0.72 | Cycloeucalenone | 7.39 | Triterpene | BP |
|
|
| no. |
| Formula | Adduct type | Precursor | Error (ppm) | Proposed identification | Score | Class | Time (day) | Specie | ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3.64 | C12H14O6 | [M-H]− | 253.0716 | 0.64 | 4,6,8-trihydroxy-3-methoxy-3,7-dimethyl-4h-2-benzopyran-1-one | 7.53 | Hydroxybenzoic acid derivatives | 14, 21, 28 |
|
|
| 2 | 4.62 | C9H16O4 | [M-H]− | 187.0971 | 2.57 | Aspinonene | 5.82 | Pentaketide | 14, 21 |
|
|
| 3 | 5.05 | C11H10O4 | [M-H]− | 205.0503 | 1.61 | Similanpyrone B | 7.72 | Isocoumarins | 14, 21, 28 |
|
|
| 4 | 5.30 | C22H18O8 | [M-H]− | 409.0921 | 1.93 | Orlandin | 6.26 | Bicoumarin | 14, 21, 28 |
|
|
| 5 | 5.47 | C18H8O8 | [M-H]− | 351.0138 | 2.39 | Thelephoric acid | 5.92 | Terphenyl | 14, 21, 28 | mushroom |
|
| 6 | 5.79 | C12H16O3 | [M-H]− | 207.1024 | 1.29 | (3R,4S)-3,4,5-Trimethylisochroman-6,8-diol | 5.30 | Chromanes | 14, 21, 28 |
|
|
| 7 | 5.88 | C31H50O8 | [M-H]− | 549.3428 | 0.89 | AB023 | 7.69 | Macrolides | 14, 21, 28 |
|
|
| 8 | 6.12 | C36H58O10 | [M+HCO2]− | 695.3998 | – | NI | – | – | 14, 21, 28 | – | |
| 9 | 6.32 | C35H54O9 | [M+HCO2]− | 663.3741 | – | NI | – | – | 14, 21, 28 |
| – |
| 10 | 6.48 | C26H38O8 | [M+HCO2]− | 523.2554 | – | NI | – | – | 14, 21, 28 |
| – |
| 11 | 6.52 | C18H34O5 | [M-H]− | 329.2325 | 2.26 | 9,10,11-trihydroxyoctadec-12-enoic acid | 8.21 | Fatty acid | 14, 21, 28 |
|
|
| 12 | 6.67 | C16H16O6 | [M-H]− | 303.0862 | 3.98 | Fonsecin B | 6.73 | Naphthopyranones | 14, 21, 28 |
|
|
| 13 | 6.71 | C14H20O3 | [M-H]− | 235.1330 | 4.10 | Terrefuranone | 5.09 | Furanones | 14, 21, 28 |
|
|
| 14 | 6.72 | C12H20O4 | [M-H]− | 227.1283 | 2.55 | Hexylitaconic acid H | 5.13 | Fatty acid | 14, 21, 28 |
|
|
| 15 | 6.83 | C11H18O4 | [M-H]− | 213.1129 | 1.55 | Hexylitaconic acid | 5.14 | Fatty acid | 14, 21, 28 |
|
|
| 16 | 7.19 | C36H58O9 | [M+HCO2]− | 679.4047 | – | NI | – | – | 14, 21, 28 |
| – |
| 17 | 7.50 | C27H40O9 | [M-H]− | 507.2604 | –0.87 | Asperterpene I | <57.11 | Steroids | 14, 21, 28 |
|
|
| [M+HCO2]− | 553.2660 | –1.11 | |||||||||
| 18 | 7.74 | C30H48O5 | [M-HCO2]− | 533.3472 | – | NI | Steroid | 14, 21, 28 |
| – | |
| 19 | 7.84 | C32H30O12 | [M-H]− | 605.1658 | 1.07 | Aurasperone B | 6.67 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 20 | 8.22 | C14H31NO | [M + H]+ | 230.2489 | –4.62 | Related to Xestoaminol c | – | Sphingolipid | 14, 21, 28 |
| – |
| 21 | 8.31 | C18H34O4 | [M-H]− | 313.2379 | 2.65 | (Z)-12,13-dihydroxyoctadec-9-enoic acid | 7.52 | Fatty acid | 14, 21, 28 |
|
|
| 22 | 8.33 | C25H49NO5 | [M + H]+ | 444.3679 | 1.02 | Hydroxyoctadecanoylcarnitine | 5.13 | Acyl carnitines | 21,28 |
|
|
| 23 | 8.48 | C32H28O11 | [M + H]+ | 589.1703 | 0.24 | Fonsecinone B or derivatives | 5.94 | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| [M-H]− | 587.1562 | –0.53 | |||||||||
| 24 | 8.82 | C26H38O6 | [M-HCO2]− | 491.2652 | 2.40 | NI | NI | 14, 21 |
| – | |
| 25 | 8.96 | C32H28O11 | [M + H]+ | 589.1707 | –0.45 | Fonsecinone B or derivatives | – | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 26 | 9.09 | C32H26O10 | [M + H]+ | 571.1610 | –1.80 | Aurasperone A or derivatives | – | Naphtho-γ-pyrone | 14, 21, 28 |
|
|
| 27 | 9.10 | C18H28O3 | [M-H]− | 291.1960 | 1.94 | Related to 9-oxooctadeca-10,12,15-trienoic acid | 5.53 | Fatty acid | 14, 21 |
|
|
| 28 | 9.14 | C18H32O4 | [M-H]− | 311.2221 | 2.50 | NI | – | Fatty acids | 14, 21, 28 |
| – |
| 29 | 9.30 | C18H36O4 | [M-H]− | 315.2530 | 3.11 | 9,10-dihydroxystearic acid | 7.23 | Fatty acid | 14, 21, 28 |
|
|
| 30 | 9.34 | C33H58O14 | [M+HCO2]− | 723.3806 | 0.36 | NI | Monoglyceride disaccharide | 14, 21, 28 |
| – | |
| 31 | 9.43 | C18H30O3 | [M-H]− | 293.2112 | 3.46 | Psiaα | – | Fatty alcohol | 14, 21, 28 |
|
|
| 32 | 9.54 | C46H34O6 | [M-H]− | 681.2310 | –4.01 | NI | – | NI | 14, 21, 28 |
| – |
| 33 | 9.60 | C27H46O9 | [M+HCOO]− | 559.3123 | 0.17 | NI | – | Monoglyceride disaccharide | 28 |
| – |
| 34 | 9.77 | C18H32O3 | [M-H]− | 295.2273 | 1.92 | FA 18:2 + 1O | – | Fatty acid | 21, 28 |
|
|
| 35 | 9.78 | C25H47NO4 | [M + H]+ | 426.3576 | 0.44 | Oleoylcarnitine | 5.53 | Acyl carnitines | 28 |
|
|
| 36 | 10.20 | C23H47NO4 | [M + H]+ | 402.3579 | –0.29 | Related to | – | Ceramide | 21, 28 |
| – |
| 37 | 10.48 | C18H34O3 | [M-H]− | 297.2431 | 1.40 | FA 18:1 + 1O | 6.55 | Fatty acid | 28 |
|
|
| 38 | 10.48 | C27H53NO8 | [M-H]− | 518.3697 | 0.27 | Related
to | – | Cerebroside | 28 |
| – |
| 39 | 10.48 | C27H48O9 | [M+HCO2]− | 561.3282 | –0.29 | NI | – | Monoglyceride glycoside | 28 |
| – |
| 40 | 11.03 | C18H32O3 | [M + H]+ | 297.2426 | –0.60 | FA 18:2 + 1O | 6.80 | Fatty acid | 14, 21, 28 |
|
|
| [M-H]− | 295.2275 | 1.24 | |||||||||
| 41 | 12.55 | C35H62O8 | [M-H]− | 655.4420 | 1.02 | NI | <5 | – | 28 |
| – |
- —Conselho Nacional de Desenvolvimento Cient?fico e Tecnol?gico10.13039/501100003593
- —Conselho Nacional de Desenvolvimento Cient?fico e Tecnol?gico10.13039/501100003593
- —Funda??o de Amparo ? Pesquisa e Inova??o do Estado de Santa Catarina10.13039/501100005667
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Production and Characterization · Polysaccharides and Plant Cell Walls · Banana Cultivation and Research
Introduction
1
Growing urbanization and lifestyle changes have colossally increased waste production, especially food residue and waste (FRW) from industries, agriculture, and households. According to the UN Environment Program, 1052 million tons of organic waste/residue worldwide were from food, with almost 2% produced in Brazil in 2022. ?,?
Based on the report from the Brazilian Institute of Geography and Statistics, a unit of the Brazilian Ministry of Planning and Budget, Santa Catarina state in 2024 was the fourth largest producer of bananas in Brazil (IBGE, 2024).? Banana peel residues account for 14% of waste in banana farming.? Poor management of these wastes can cause environmental impacts (gas emissions, contaminated groundwater, and noxious odors) and consequences for economic income associated with natural resources such as water and land. A variety of sustainable techniques are currently used to reduce the impact of FRW. Although in some cases FRW is used to produce energy, compost, animal feed, and biofertilizers, in others, it is sent to landfills or incinerated. Other strategies, such as biotechnology processes that produce valuable bioactive compounds from FRW, contribute to the bioeconomy initiative. These bioactive compounds can be turned into food supplements or food ingredients known to possess health benefits; they can also contribute to drug research and discovery.? Therefore, FRW can undergo organic solvent extraction and purification to identify these valuable substances.? More effective management and wider integration into economic, research, and agricultural sectors are important to mitigate the impact of FRW, particularly banana peels. As shown in previous studies, banana peels have nutritive values and can be considered a functional food because of their content of minerals (iron, calcium, magnesium, and sodium), amino acids (valine, threonine, and phenylalanine), phenolic acids (gallic, tannic, ferulic, and caffeic acids), flavonoids (rutin and quercetin), carotenoids (β- and α-carotene), terpenoids (stigmasterol), and fatty acids (palmitic acid). ?,?
Thus, banana peels can also be used as biomass for microbial fermentation to generate high-value products.? This last process is known to produce flavoring for fermented beverages; lactic, citric, and succinic acids; enzymes (lipase, amylase, cellulase, and others); ethanol; and hydrogen. ?−? ? ? ? A previous study reported that solid- and liquid-state cultivation of Aspergillus niger using orange peels as a substrate produced protease.? In addition, Streptomyces sp. grown on cassava peel, groundnut shell, or wheat bran-based solid-state cultures produced neomycin, oxytetracycline, and meroparamycin. ?,? Therefore, the use of FRW as biomass is fundamental for a circular bioeconomy. Based on the aforementioned information, this work aimed to investigate the chemical response of A. niger when grown on a biomass composed of banana peels. The species A. niger was selected for this study because it is an airborne fungus with wide application in industrial biotechnology and because it is prevalent in household waste used for biomass, particularly in banana peels used in this work. Experiments using A. niger cultivated on FRW biomasses such as wheat bran, coffee pulp, pineapple canary waste, tomato waste, and apple pomace produced citric acid, lycopene, biosorbents, vanillic acid, vanillin, and polygalacturonase.? The profile of secondary metabolites varies according to the type of FRW, representing an advantage in the identification of new byproducts and valuable chemical substances with potential applications in drug discovery and as bioinoculants, fungicides, and biostimulants, among others. Previous studies used banana peels as biomass to grow A. niger to produce bioethanol, citric acid, and enzymes such as xylanase, amylase, and pectinase. ?−? ?
In general, the use of food waste for fungal cultivation not only produces valuable substances but also represents an economical process when compared with conventional techniques that produce metabolites derived from microorganisms using purchased items, including biomass (rice, agar, and starch) and other nutrients. In this study, the hypothesis was to investigate whether organic waste, such as banana peels, can serve as biomass for microorganisms, enabling the biosynthesis of biologically relevant compounds. Furthermore, emphasis was placed on the secondary metabolites of banana peel and their impact on the chemical response of A. niger.
To date, secondary metabolites generated by A. niger grown on biomass made of banana peels have never been investigated.
We herein report a comparative study using the dereplication technique and statistical tools to analyze LC-MS chemical profiles of A. niger cultivated on banana peels, starch-enriched banana peel, and starch. In addition to the culture media used for A. niger growth, fermentation periods of 14, 21, and 28 days were also used as comparison parameters.
Materials and Methods
2
Materials
2.1
Acetonitrile, methanol, water, isopropanol (LCMS grade), and leucine entkephalin were obtained from Servylab, a Sigma-Aldrich seller (Rio Grande do Sul, Brazil). The syringe filter (0.22 μM, hydrophobic), vials (2 mL), and hydrophobic and hydrophilic filter membranes for solvent filtration were furnished by Filtrilo (Paraná, Brazil). Ethyl acetate of analytical grade was furnished by Rauter (Gravata, Rio Grande do Sul).
Fungus Collection and Characterization
2.2
Banana peels (Musa balbisiana), popularly known as “banana figo”, were purchased from a local market in the city of Florianópolis, Santa Catarina, Brazil. To reproduce how microorganisms colonize and consume organic matter in nature and to make sure that the selected microorganism will grow on this biomass, the peels were washed in running water to remove contaminants, and then, a quantity (5 g) was ground in 50 mL of distilled water in a beaker. The beaker containing this organic material was kept in the lab for 7 days until colonies of microorganisms appeared. The predominant fungus was isolated on Potato Dextrose Agar plates and grown for 21 days at 28 °C. Further, the strain was identified by micromorphological characterization, applying the adapted Riddell technique,? and confirmed by the amplification and sequencing technique of the ITS gene (ITS region: ITS1), performed by the biotechnology company Neoprospecta in Florianópolis, Santa Catarina, Brazil. The sequence was registered in the NCBI database under GenBank number PX619933.
Substrate
Preparation and Inoculation
2.3
The spore suspension used as inoculum was prepared by adding Tween 80 (0.01% v/v to the fungus colony previously cultivated on BDA plates at 28 °C for 7 days, following the protocol described by Robl et al. (2015).? The spore counting was performed by using a Neubauer chamber. Initially, the concentrated suspension was diluted 10x in an Eppendorf tube. After homogenization using a vortex mixer, two 20 μL aliquots of the diluted solution were transferred to the chamber. By using a microscope with 40× magnification, the spores present in the five diagonal squares of the central quadrant of the chamber were counted. The spore concentration in the suspension was calculated using the following equation:
The spore suspension was diluted in order to reach the desired final concentration of 10^6^ spores·mL^–1^ in the culture medium.
Three flasks received a ratio of 50 g of banana peel to 1 L of deionized water as the culture medium. These flasks were covered with cotton and autoclaved at 121 °C, 1 atm for 30 min.? Soon after, at room temperature and within a laminar flow, the inoculum of A. niger was added to the culture media and incubated in B.O.D. at 28 °C. A similar procedure was performed with starch in a concentration of 50 g/L.
The mixture of starch and banana peels to formulate an alternative culture medium was motivated by previous works whose results indicate that varying carbohydrate sources (such as sucrose, lactose, mannitol, potato dextrose, flour, and starch) can influence fungal growth and the biosynthesis of specific biomolecules. ?,? Thus, the impact of supplementing the banana peel biomass medium with starch as a carbon source was assessed in relation to the profile of secondary metabolites. Starch quantity was not assessed as a culture parameter, but literature suggests 20 g/L is optimal for A. niger growth.? Thus, a 1:1 (w/w) banana peel/starch mix totaling 50 g in 1 L of distilled water to cultivate the fungus was prepared in three different flasks. The cultivation was carried out in the periods of 14, 21, and 28 days at a constant temperature of 28 °C. Thus, three culture media containing only banana peel (BP_An), three containing banana peels enriched with starch (BPS_An), and three containing starch (Starch) were obtained.
Preparation
of Crude Extracts
2.4
After each time of fungus cultivation, ethyl acetate was added to stop the fungus growth and perform the extraction. The organic extract was obtained with 200 mL of ethyl acetate assisted by sonication in an ultrasonic bath for 1 h, following the solid–liquid extraction process; these steps were repeated three times for each medium, thus obtaining a total of 6 extracts. The extracts were concentrated by rotary evaporation. As intended to obtain a blank for the extracts, this same extraction procedure was applied to samples from inoculated starch, noninoculated banana peel (BP), and noninoculated starch-enriched banana peels. The aim was to discriminate banana-peel- and starch-derived metabolites from those produced by the fungus.
Ultra-Performance Liquid Chromatography-Tandem
Mass Spectrometry Analysis (UPLC-ESI-MS/MS)
2.5
The obtained extracts were prepared by dissolving 1.6 mg in 2 mL of methanol/acetonitrile (1:1 v/v, LC-MS grade), resulting in a concentration of 800 μg/mL. Before analysis, all extracts were filtered through a 0.22 μm hydrophilic PTFE filter (Analytical).
An Acquity UPLC system class H (Waters Co., USA) equipped with a photodiode array detector (PDA), sample manager, and a quaternary solvent manager as well as a BEH C18 column (50 mm, 1.0 mm, particle size 1.7 μm (Waters)) was used for the separation. The column and the sample tray were maintained at temperatures of 40 and 20 °C, respectively. Each sample (2 μL) was injected and analyzed under gradient conditions at a flow rate of 0.3 mL/min. The mobile phase was composed of H_2_O containing 0.1% formic acid, pH 3.0 (A) and LC-MS-grade acetonitrile (B). The gradient was the following: between 0 and 1 min, 90% A/10% B; between 1 and 12 min, 10% A/90% B; between 12 and 14 min, 10% A/90% B; between 14 and 15 min, 90% A/10% B; and between 15 and 20 min, the initial mixture was applied to equilibrate the column.
A mass spectrometer, Xevo G2-S QTof (Waters), bearing an electrospray ionization (ESI) probe operating in positive and negative ionization modes, coupled to the UPLC device was used to detect the chemical components of the extracts. Nitrogen was used as nebulizer gas; chromatograms were acquired with the cone gas flow of 100 L/h, the desolvation gas flow of 900 L/h, the sampling cone current of 40 V, and the source offset current of 80 V. The collision gas was argon, and the lockspray reference sample was leucine enkephalin with reference mass values at m/z 554.2615 (ESI−) and 556.2771 (ESI+). The desolvation and the ionization source were maintained during the analyses at 250 and 90 °C, respectively, while the capillary voltage was 3 kV. A range of 30–35 eV was used as the collision energy. Data were acquired over a range of 50–1500 Da in positive mode and 50–1500 Da in negative mode, at a scan time of 1.0 s over 20 min, and were processed with MassLynx V4.1 (Waters). The MS data were acquired in Fast Data-Dependent acquisition (Fast DDA) and MS^E^ (DIA) settings using argon as collision gas and applying the energy range of 25–35 eV (Supporting Information).
Data
Processing by MS DIAL/MS FINDER
2.6
A combination of MS-DIAL (version 4.92) and MS-FINDER (version 3.52) software was used to assist in establishing the chemical profile. The raw data from the Fast DDA analyses were loaded directly into MS-DIAL, where the information was selected according to the analyses performed by UPLC-MS, and sections for data processing and alignment were selected. The values applied to the parameters were: tolerance of 0.02 and 0.03 Da for MS1 and MS2, respectively; peak detection amplitude of 10000 for the minimum peak height and 0.1 Da for the mass cutoff width; for deconvolution, a sigma window value of 0.5 and an MS/MS abundance cutoff of 30 amplitudes; the retention time tolerance was set to 0.5 min and 0.02 Da for MS^1^ tolerance. The option to remove features based on solvent blank data was also applied, thus obtaining annotations referring only to the samples.?
The data obtained in MS-DIAL were exported to MS-FINDER, where the identification of features was provided by comparing experimental MS/MS fragmentation spectra from several spectral databases (COCONUT, PubChem, UNPD, ChEBI, NANPDB, KNApSAcK, NPA). To select the structure as a potential candidate, a score value greater than or equal to 5 among the fragments obtained and knowledge of the possible classes of compounds that the sample type can produce were considered.?
The features that did not lead to structure proposals by the software were subjected to the dereplication technique using MS^E^ fragmentation data in comparison with those reported in the literature. The molecular formulas were determined by using the MassLynx elemental composition tool. Each molecular formula was chosen with a tolerance lower than or equal to ±5 ppm between the calculated and measured mass values.
Molecular Network by GNPS
2.7
The raw Fast-DDA data acquired by UPLC-MS for the extracts were converted into the corresponding format (mzML) using MSConvert software (ProteoWizard, Palo Alto, CA, USA) and uploaded to the GNPS online platform using Core FTP LE software v. 2.2. Molecular network analysis was performed following the online workflow (https://ccms-ucsd.github.io/GNPSDocumentation/) described on the GNPS web site.? Molecular networks were obtained using a precursor ion mass tolerance of 0.02 Da and an MS/MS fragment ion tolerance of 0.03 Da. Network edges were produced only with a cosine score above 0.77 and a minimum match of 4 peaks in the fragmentation spectrum. The spectral data in the network were then compared with the GNPS spectral libraries. For matches of the GNPS library with the data, a score above 0.7 and at least 4 matching peaks were applied. The generated molecular networks were visualized using Cytoscape software, version 3.10.2.
Statistical
Analysis
2.8
For statistical analysis, peak intensity data deconvolved by MS-DIAL were extracted and reorganized into a table in CSV format. Univariate and multivariate statistical analyses were performed on the MetaboAnalyst 6.0 online platform (https://www.metaboanalyst.ca/) to discriminate significant metabolites between the culture media (BP_An, BPS_An, noninoculated BP and starch), as well as the metabolites at different growth times (14, 21, and 28 days). Data were normalized by square root transformation and Pareto scaling. Principal component analysis (PCA) and partial least squares discriminant analysis (PLS-DA) were used to characterize the distribution of the data set and determine the main metabolites responsible for the distribution, the latter based on the thresholds of the variable importance in projection (VIP) value >1 and p-value <0.05. Heat plots were applied to visualize the relationship between the samples and the abundances of the compounds in each sample.
Results
and Discussion
3
Fungal Isolation and Cultivation
3.1
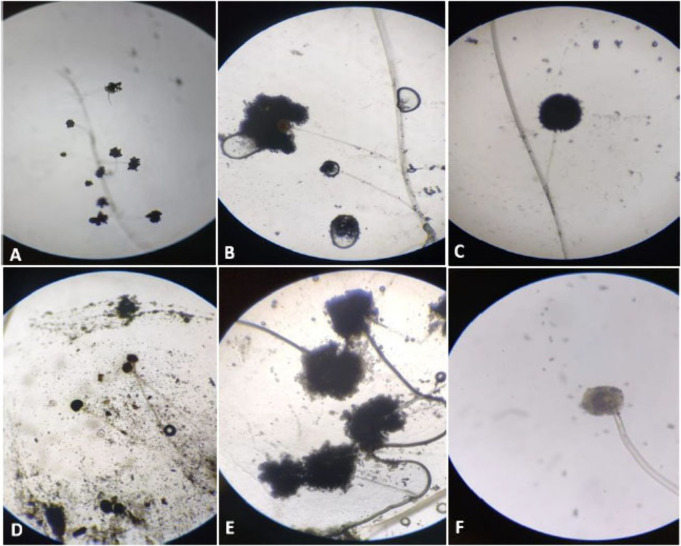
The isolated strain was identified through micromorphological features as an Aspergillus species (Figure and Figure S1–S3), applying an adapted Riddell technique.? Using the slide microcultivation technique, it was possible to verify the microscopic characteristics of the fungal isolate under an optical microscope. Classic characteristics of the fungus A. niger were observed, including abundant black conidiophores and conidia and a smooth hyaline wall.
Micromorphological aspect of the fungus evaluated at 7 and 14 days of cultivation (micro cultivation). (A–C) 7 days of micro cultivation, (D–F) 14 days of micro cultivation. (A and D) 20× magnification; (B, C, E. F) 40× magnification.
Molecular identification using amplification and sequencing techniques of the ITS gene region (ITS1) matched the strain of Aspergillus niger with 99% homology. Sequencing of this region of the fungus, renamed Aspergillus niger LQPN001, resulted in the following sequence:
TTACCGAGTGCGGGTCCTTTGGGCCCAACCTCCCATCCGTGTCTATTGTACCCTGTTGCTTCGCGGACCCGCCGCTTGTCGGCCGCCGGGGGGGCGCCTCTGCCCCCCGGGCCCGTGCCCGCCGGAGACCCCAACACGAACACTGTCTGAAAGCGTGCAGTCTGAGTTGATTGAATGCAATCAGTTAAAACTTTCAACAATGGATCTCTTGGTTCCG
The BLASTN search resulted in four species with 99.54% similarity of the ITS genetic code, all with E-value of 2–111 and 100% coverage of the searched gene: Aspergillus fetidus (CBS 121.28), Aspergillus awamori ATCC 16877 (CBS 557.65), Aspergillus niger ATCC 16888 (CBS 554.65), and Aspergillus welwitschiae (CBS:139.54).
The fungus, A. niger, was grown on Musa balbisiana banana peels, starch-enriched peels, and starch for 14, 21, and 28 days, producing nine crude extracts: BP_An14d, BP_An21d, BP_An28d, BPS_An14d, BPS_An21d, BPS_An28d, Starch_An14d, Starch_An21d, and Starch_An28d. The chemical profiles of these crude extracts, along with those obtained from banana peels, were established by using LCMS data. The LCMS profile of the starch-enriched medium without inoculation was similar to that of banana peels. However, this profile showed low sensitivity due to the low concentration of metabolites.
Statistical Analysis
3.2
To perform a comprehensive comparison correlating all variables and determining the differences between culture media and cultivation times, as well as identifying the main metabolites responsible for this variation, multivariate analyses were applied. Data obtained from the positive and negative ionization modes were used separately for the comparison.
Principal component analysis (PCA) revealed significant differences between the chemical profiles of the samples, forming clusters, as depicted in Figure. The first two principal components (PC1 and PC2) explained 65% and 76.7% of the total variance of the data obtained from the positive and negative mode analyses, respectively. These PC values best represent the correlation between the groups.
PCA plot with metabolites identified in the extracts obtained from the cultivation of the fungus A. niger in BP, BPS, and starch for 14, 21, and 28 days of growth in positive (ESI+) and negative (ESI−) ionization modes.
The PCA plot in positive mode analysis showed no proximity of BP_An21d to other samples. In contrast, it was grouped in a negative mode with BP and BPS samples except for BPS_An28d. This difference suggests that the BP_An21d sample contains metabolites that behave differently according to the ionization modes. Additionally, the BPS_An28d sample stood out for presenting a unique profile in both ionization modes. This sample was alone in the cluster, indicating a significantly different chemical composition profile. All samples from the starch medium formed a cluster in both ionization modes and were far from the BP and BPS samples.
The statistical analysis focused on a list of MS peaks and their corresponding areas ranked using t tests. From this analysis, the top 15 metabolites identified through the variable importance in projection (VIP) from partial least-squares discriminant analysis (PLS-DA) in both positive and negative modes were selected for correlation in the heatmap. The VIP plot produced by the PLS-DA models ranked metabolites based on their ability to differentiate between sample groups, including the biomass sample and those obtained from various fermentation times. The color scale (ranging from blue to red) indicates the relative amounts of each metabolite across various sample groups (red indicates a high quantity, and blue is not present). Previously identified metabolites are shown with their compound names, while additional metabolites are indicated by NI (Non Identified) followed by their precursor ion (m/z) found on the left side of the plot. The sample groups were based on fermentation media (starch and banana peels medium, BP; and banana peel medium enriched with starch, BPS) and duration (14, 21, and 28 d), are labeled at the bottom of the plot.
The VIP plot reveals nonadecan-2-amine (score >3) and the alkylaryl amine derivative, m/z 304.3005 (score
2.5) as the most concentrated compounds in the positive mode ionization (Figure). In contrast, it shows the fatty acid derivative (m/z 311.221) (score >1.8) and the monoglyceride disaccharide (m/z 559.3123) (score = 1.8) as the most prevalent in the negative mode ionization.
Fifteen most important metabolites from partial least squares discriminant analysis variable importance (PLS-DA VIP) from the cultivation of the fungus A. niger in BP, BPS, and starch at 14, 21, and 28 days of growth in positive (ESI+) and negative (ESI−) ionization modes.
The heatmap in Figure, displaying the top 60 metabolites based on the ANOVA p-value, was created using MetaboAnalyst 5.0 to forecast the distribution of metabolites during the fermentation timeline of 14, 21, and 28 days. In positive mode ionization, the chemical profile of BPS_An14d was close to that of banana peels (BP). Both were hierarchically close to BPS_An21d. The sequence of similarity in the hierarchy was followed by banana peel medium fermented for 28 days (BP_An28d), then BP_An14d, BP_An21d, BPS_An28d, Starch_An21d, Starch_An14d, and Starch_An28d. As observed in the PCA plot, fermentation periods of 21 to 28 days were suitable for A. niger to produce highly diverse secondary metabolites when grown on the banana peel and starch-enriched banana peel media, respectively. Moreover, a fermentation time of 28 days was the best for this fungus to produce more natural products.
Heat map plot with 60 top metabolites identified in the extracts obtained from the cultivation of the fungus A. niger in BP, BPS, and starch at 14, 21, and 28 days of growth in positive (ESI+) and negative (ESI−) ionization mode.
The hierarchical distribution was different for the negative mode of ionization. Four groups were observed; among them, the first was composed of BP_An21d, BP_an14d, and BP_An28d. This group was close to BPS_An21d, BPS_An14d, and BP. Both clusters were close in hierarchy to BPS_An28d. The samples Starch_An21d, Starch_An14d, and Starch_An28d formed a group that was the end of the hierarchy. The present analysis also revealed that the fungus grown on media prepared with banana peels, produced in all periods, is a secondary metabolite sensitive to the negative ionization mode. However, a fermentation time of 28 days was optimal for significantly producing fungus-derived metabolites on the starch-enriched banana peel medium.
Dereplication
of the Chemical Profiles
3.3
Crude extracts obtained from the fermentation, as well as those from banana peels and starch, were subjected to UPLC-ESI-QTOF-MS analysis. Generated mass spectrometry data (Figures S4–S9) were converted into a compatible format and then processed using MS-Dial, MS-FINDER, and GNPS.
The data obtained are compiled in Tables–?, in which the structures proposed by the aforementioned software and databases received scores from 0 to 10 (highest rate). Structure assignments without a score were possible using the dereplication technique in conjunction with previous studies reported in the literature. LCMS data from banana peels (BP) and samples from the fermentation of A. niger on banana peels for 14 (BP_An14d), 21 (BP_An21d), and 28 (BP_An28d) days revealed the presence of 76 secondary metabolites, among them, 10 substances were found in BP. The structures of banana peel metabolites were assigned as a polyol (m/z 261.1316), piptamine (m/z 332.3312), nonadecan-2-amine (m/z 284.3316), linolenic acid derivative (m/z 279.2327), isoorientin (m/z 471.1050), eriodyctiol-6-C-beta-d-glucopyranoside (m/z 473.1027), di(-2-ethylhexyl)phathalate (m/z 391.2848), icosalide B (m/z 685.4361), di(2-ethylhexyl)phthalate (m/z 413.2653), and cycloeucalenone (m/z 425.3781). In addition to secondary metabolites from banana peels, twenty-seven others were previously reported in A. niger, ?,? while 18 were formerly described in other Aspergillus species. ?,? Nigragillin was found at 1.63 min with a m/z of 223.1811, and the same mass value also appeared at 0.76 min, suggesting either a pyrazine or a piperazine derivative. The low fragmentation limits the consistent characterization of this secondary metabolite. However, pyrazines such as 2-methoxy-3,5-diisobutylpyrazine, 2-methoxy-3,6-disec-butylpyrazine, and 2-methoxy-3,6-diisobutylpyrazine, with the same elemental composition, were obtained from the bacteria, Carpophilus humeralis.? Pyranonigrin A (m/z 224.0557) together with other components, namely, phenoxyacetic acid (m/z 153.0553), cycloleucomelone (m/z 339.0503), atromentin (m/z 323.0550), carbonarone A (m/z 230.0819), pyrophen (m/z 310.1057), nigerpyrone (m/z 187.0759), hexylcitric acid (m/z 275.1126), orlandin (m/z 411.1079), pestalamide A (m/z 344.1132), demethylkotanin (m/z 425.1232), aspernigrin B (m/z 457.1759), hexylitaconic acid (m/z 213.1126), nigerasperone C (m/z 575.1550), kotanin and its isomer (m/z 439.1388), aurasperone D (m/z 555.1280), flavasperone (m/z 287.0920), fonsecinone B (m/z 589.1706), nipyrone (m/z 261.1464), and aurasperone A (m/z 571.1605) previously identified in A. niger were also found in this study. ?,? Most of the naphtho-γ-pyrones were detected alongside their positional or functional isomers (Table).
1: Identification of Metabolites Using MS-FINDER Software and UPLC-ESI-HRMS Analysis in Positive and Negative Modes for Extracts Obtained from the Fungus A. niger Grown on Banana Peel (BP), at 14, 21, and 28 Days
Part of the other metabolites produced by A. niger in this study were previously reported in other Aspergillus species. Ochramide B, 2,10-dimethyl 4-hydroxy-6-oxo-4-undecen-7-yne (m/z 209.1541), similanpyrone B (m/z 207.0657), a pyrazine, a β-hydroxyenone, and an isocoumarin, produced respectively by A. ochraceus
?,? and A. similanensis,? were detected in the present study. Moreover, PsiAβ (m/z 297.2428) and PsiAα (m/z 293.2116), previously obtained from A. nidulans, were also detected in this study.? Other secondary metabolites, including asperlone B, Sch725674 (m/z 351.2143), asperfuranone A (m/z 259.1312), a 12,13-dihydroxyoctadec-9-enoic acid derivative (m/z 337.2350), 8,11,12-trihydroxyoctadec-9-enoic acid derivative (m/z 313.2373), proversilin A (m/z 273.1468), 9,10-dihydroxystearic acid derivative (m/z 339.2511), 6-keto stearic acid derivative (m/z 299.2588), 9-oxooctadeca-10,12,15-trienoic acid derivative (m/z 291.1952), linolenic acid (m/z 279.2324), and N-[2-Hydroxy-1-(hydroxymethyl)octadecyl]hexadecanamide (m/z 554.5506) formerly identified in a partially sequenced Aspergillus were also characterized.
Based on literature searches, two of the annotated compounds, thelephoric acid and 3-hydroxy-4-(6-methoxy-5,8-dioxonaphthalen-2-yl)-5-oxofuran-2-ylidene](4-hydroxyphenyl)acetic acid, were previously identified in the mushrooms, Polyozellus multiplex ? and Xerocomus badius,? respectively. This study also detected botcinic acid and a harzianumol derivative, previously identified in the fungi Botrytis cinerea and Trichoderma harzianum, respectively. ?,? In addition, menaquinone 7 and salinipyrone A, two bacteria-derived metabolites, ?,? were detected in the A. niger profile.
Table also displays metabolites that were not described in any microorganisms, which include 2-methyl-N-(2-phenylethyl)propanamide (m/z 192.1386), 2-methyl-N-(2-phenylethyl)butanamide (m/z 206.1546), 8,11,12-trihydroxyoctadec-9-enoic acid derivative (m/z 353.2297), a nor-diterpene (m/z 263.2376), two isomers of 4α,14α-dimethyl-24-methylene-cholest-7,9(11)-dien-3β-ol (m/z 425.3776), two isomeric triterpenes (m/z 457.3662), apocholic acid (m/z 391.2849), and cis/trans-erucamide derivatives (m/z 338.3422).
The structure assignment of part of the proposed compounds was checked to sustain the MS-FINDER proposal by using the reported fragmentation pattern? of ions listed in Tables S1 and S2. Schemes S1–S6 display the structure of fragment ions that led us to the chemical profile in Tables and ?.
2: Identification of Metabolites Using MS-FINDER Software and UPLC-ESI-HRMS Analysis in Positive and Negative Modes for Extracts Obtained from the Fungus A. niger Grown on Starch-Enriched Banana Peel Medium (BPS), at 14, 21, and 28 Days
The peak at a retention time (tR) of 1.63 min with m/z 223.1811 [C_13_H_22_N_2_O + H]^+^ was assigned to the metabolite nigragilin. Its tandem mass spectrum displays m/z 129.1386 [C_7_H_17_N_2_ + H]^+^ corresponding to N-methylpiperazinium. This ion resulted from the loss of the side chain (94 Da). Another fragment at m/z 95.0494 [C_6_H_7_O]^+^ was assigned as an acylium ion, formed through α-cleavage of the C–N bond of the amide functionality and elimination of the N-methylpiperazine moiety.
The ion at tR 5.04 min with m/z 323.0550 [C_18_H_12_O_6_ – H]^−^ present only in BP_An28d and BPS_An28d was annotated as atromentin. Its MS^2^ spectrum showed the fragment ion m/z 295.0615 [C_17_H_12_O_5_ – H]^−^ yielded after the precursor lost CO (28 Da).
The peak at tR 6.15 min with m/z 379.0450 [C_20_H_12_O_8_ – H]^−^, present in the three cultivation times, was assigned as asperlone B. In its tandem mass spectrum, m/z 361.0362 [C_20_H_10_O_7_ – H]^−^ and 333.0400 [C_19_H_10_O_6_ – H]^−^ were observed, consistent with the loss of H_2_O (18 Da) and CO (28 Da), respectively.
The metabolite pestalamide A, known as an *A. niger-*derived compound, was annotated in positive mode with m/z 344.1132 [C_18_H_18_NO_6_ + H]^+^. Its MS^2^ spectrum showed a fragment m/z = 230.0826 [C_13_H_10_NO_3_ + H]^+^. This ion was produced after the precursor lost the 2-methylbutane-dioic acid moiety. The ion m/z 230.0826 in turn produced the lighter ions m/z 213.0548, 185.0512, and 157.0647 after eliminating NH_3_ (17 Da), NH_3_/CO (45 Da), and NH_3_/2CO (73 Da), respectively. In negative mode, the metabolite pestalamide A (m/z 342.0970) gave fragment ions m/z 228.0634 [C_13_H_11_NO_3_ – H]^−^ and m/z 185.0620 [C_12_H_10_O_2_ – H]^−^ in its MS/MS spectrum. These fragments corresponded to sequential elimination of the 2-methylbutane-dioic acid moiety and NH_3_ (17 Da), respectively.
The peak at tR 6.38 min with m/z 213.1126 [C_11_H_18_O_8_ – H]^−^ in negative mode was attributed to hexylitaconic acid. In its MS^2^ spectrum, the observed fragment m/z 169.1219 [C_10_H_18_O_2_ – H]^−^ differs from the precursor by CO_2_ (44 Da).
The peak at tR 7.98 min with m/z 339.2511 [C_18_H_36_O_4_ + Na]^+^ and 315.2531 [C_18_H_36_O_4_ – H]^−^, in positive and negative mode, respectively, was annotated as 9,10-dihydroxystearic acid. In the negative mode, the obtained fragment m/z 297.2421 [C_18_H_34_O_3_ – H]^−^ corresponded to the loss of H_2_O (18 Da).
Starch was added to the culture media as a carbohydrate source to enhance the fungal catabolism and metabolism. Out of this process, 99 secondary metabolites were detected and are reported in Table. Part of these compounds were plant metabolites and were annotated as hexadecasphinganine (m/z 274.2744), an alkylarylamine (m/z 304.3005), 1,10-epoxy-8alfa-methoxyvermophilanolide (m/z 279.1597), an isomer of Val-Val-Lys-Lys (m/z 473.3453), a peptide (m/z 517.3712), linolenic acid (m/z 279.2322), 2-aminohexadec-11-en-3-ol (m/z 256.2641) alongside isoorientin (m/z 471.1050), eriodyctiol-6-C-beta-d-glucopyranoside (m/z 473.1027), etalon (m/z 391.2848), and cycloeucalenone (m/z 425.3781). Part of the metabolites found exclusively in banana peel-derived media were not detected in those composed of starch-enriched banana peel. On the other hand, these starch-enriched media also yielded compounds that were not detected in media without starch. This discrepancy can be associated with microorganism biochemical activities that change according to the chemical properties of the culture medium.? In this way, the medium chemical composition modulates different metabolism activities. Growing A. niger on starch-enriched banana peel media afforded 22 secondary metabolites previously identified in the same species; 19 additional compounds herein reported were already found in other Aspergillus species (with genomes either fully or partially identified). In addition, 25 of these fungus-derived metabolites were already described in mushrooms, bacteria, fungi, association fungi/bacteria, and algae species. Among the previously reported A. niger secondary metabolites, 18 were found in both media. However, tensyuic acid A (m/z 243.0867), the peptide (m/z 625.2447), aspernigrin C (m/z 571.2076), and tensyuic acid D (m/z 295.1158) were generated only in starch-enriched banana peel media.
In banana peel media, the biosynthesis of previously identified metabolites also occurred. These compounds included 9-hydroxyhexylitaconic acid (m/z 229.1070), penicillenol C1 (m/z 294.1698), brevianamide M (m/z 322.1180), pulvinic acid (m/z 309.0762), asperfuranone (m/z 377.1590), and aspergilloid E (m/z 341.1616) previously reported in Aspergillus species with the fully identified genome. ?−? ? ? ? ?
Table also displays substances previously found in Aspergillus species with partially identified genomes. Two of these compounds, namely, asperlone B (m/z 379.0445) and sch725674 (m/z 351.2143), were biosynthesized in both cultures, whereas aspergilactone A (m/z 213.1128), purpurquinone A (m/z 455.0969), phomoligol A (m/z 329.1231), β-d-glucopyranosyl aspergillusense A (m/z 441.2113), asperlone B (m/z 379.0445), JBIR-138 (m/z 363.1438), aspertetranone C (m/z 421.1856), mitorubrinic acid (m/z 435.0713), carnemycin B (m/z 451.1954), and 2-aminohexadec-11-en-3-ol (m/z 256.2641) were found only in starch-enriched media. These culture media also produced harzianopyridone (m/z 282.1344), which was previously identified in Trichoderma harzianum.? In addition, 1,2-diacetoxy-3-(4′hydroxyphenyl)-4,7,8-trihydroxydibenzofuran (m/z 425.0866), roridinic acid (m/z 301.1281), hamigeromycin A (m/z 393.1539), and macrosphelide I (m/z 343.1387) were formerly identified in the mushroom Sarcodon laevigatum ? and the fungi Cordyceps sp.,? Hamigera avellamea,? and Periconia byssoides.? Atromentic acid (m/z 341.0660) from Boletus permagnificus,? inoscavin B (m/z 437.1229) from Innonotus xeranticus, [3-hydroxy-4-(6-methoxy-5,8- dioxonaphthalen-2-yl)-5-oxofuran-2-ylidene](4-hydroxy phenyl)acetic acid (m/z 435.0708) from Xerocomus badius,? pyrenocine L (m/z 307.1158),? 2-hexylidene-3-methylsuccinic acid – methyl ester (m/z 227.1280) from Halorosellinia oceanica,? myxotrichin C (m/z 493.1131) from Myxotrichum spp ? colletobredin C (m/z 427.2324) from Colletotrichum Aotearoa,? herqueichrysin (m/z 357.1336) from Penicillium herquei,? talaroketal A (m/z 627.1865) from Talaromyces stipitatus,? trichoflectin (m/z 299.0920) from Trichopezizella nidulus,? salinipyrone A (m/z 315.1571) from Salinispora pacifica,? and botcinic acid (m/z 425.2150) from Botrytis cinerea ? were also annotated in this second medium. The structures of 26 detected compounds could not be characterized.
The GNPS database was also used to annotate the metabolites and correlate those of the same classes using molecular networks. In this technique, each compound is represented by a node and each node contains a pizza diagram according to its proportion in different analyzed samples. The network connects substances that are stereoisomers, positional, and functional isomers.?
GNPS analysis of BP_An14d, BP_An21d, and BP_An28d detected 45 nodes; among them, 28 match substances in its database (Figure). In the positive mode ionization, the plant secondary metabolites isoorientin and eriodictyol-6-C-β-d-glucopyranoside form a cluster as both differ by one unsaturation. Although six bicoumarins were detected and identified (Tables and ?), only kotanin, isokotanin B, and orlandin were detected in GNPS. These metabolites formed a cluster as they are structurally related and differ from one another by the number of methoxy groups. Two alkamides, including 2-methyl-N-(2-phenylethyl)propanamide and 2-methyl-N-(2-phenylethyl)butanamide, also formed a cluster, as the difference between both compounds is 14 Da (methyl group). Other compounds detected by GNPS did not form clusters, including those of the negative mode ionization. The MS-FINDER database proposed the structure of aurasperone A for m/z 571.1605, which was found in 21 and 28 days of fermentation. However, GNPS identified for the same mass value, a biflavonoid known as 4′,4′′′-di-O-methylisochamaejasmin. This database also revealed that m/z 571.1605 is present in banana peels.
Molecular network from positive (ESI+) and negative (ESI−) ionization modes of extracts from A. niger grown on banana peel (BP), for 14, 21, and 28 days. Where yellow = BP, orange = 14 days, purple = 21 days, and green = 28 days.
Analysis of BPS_An14d, BPS_An21d, and BPS_An28d led to the detection of 45 nodes; among them, 20 were matched secondary metabolites in the GNPS database (Figure). Like the above-mentioned analysis, a cluster was formed among the two flavonoids, isoorientin and eriodictyol-6-C-β-d-glucopyranoside. A group was also formed by cylcoeucalenone, botcinic acid, and an unidentified compound. However, the combination might be due to the misinterpretation of the database, since one is a triterpene and the other is a polyketide. Part of the metabolites composed of flavasperone (m/z 287.0920), hexylcitric acid (m/z 275.1126), JBIR-138 (m/z 363.1438), β-d-glucopyranosyl aspergillusene A (m/z 441.2113), and the peptide related to Glu-Arg-Phe (m/z 449.2135) was found only in this analysis.
Molecular network from positive (ESI+) and negative (ESI−) ionization modes of extracts from A. niger grown on banana peel with starch (BPS), for 14, 21, and 28 days. Where in yellow = BP, orange = 14 days, purple = 21 days, and green = 28 days.
The studied fungus was also cultured on starch for 14, 21, and 28 days, affording Starch_An14d, Starch_An21d, and Starch_An28d, respectively (Table). LCMS analyses led to the detection of 41 compounds; among them, 29 were characterized. The fungus on starch medium produced fewer secondary metabolites than when grown on BP and BPS. Fifteen of these metabolites were already identified from other Aspergillus species, while eight were unique to A. niger and four to other species (Figure). The GNPS analyses depicted a node between the unidentified metabolites m/z 430.3890, 428.3730, 416.3730, and 402.3579, a sphingolipid identified as N-(propionyl)-4-hydroxyeicosasphinganine. The first metabolite (m/z = 430.3890) differs from the sphingolipid by 18 Da, indicating the presence of an OH group. Then, the metabolite m/z 428.3730 has a double-bond equivalence less than that of the first metabolite. The last metabolite (m/z 416.3730) is 14 Da higher than the sphingolipid, indicating a longer side chain by an additional CH_2_ group. GNPS obtained from ESI(+) identified kotanin (m/z = 439.1380) and two related bicoumarins at m/z = 425.1230 (demethylkotanin) and 411.1380 (orlandin) in the same cluster. This analysis also depicted three steroids m/z 509.2750 (asperterpene I), 525.271, and 493.2800 forming a cluster the same as the sphingolipids at m/z 274.2750, 318.3010, and 362.3260. Aurasperone A (m/z 571.159) and fonsecinone B (m/z 589.171) differ by 18 Da, despite no node being observed between them.
3: Identification of Metabolites Using MS-FINDER Software and UPLC-ESI-HRMS Analysis in Positive and Negative Modes for Extracts (Starch_An14d, Starch_An21d, and Starch_An28d) Obtained from the Fungus A. niger Grown on Starch at 14, 21, and 28 Days
Molecular network extracts in positive (ESI+) and negative (ESI−) ionization modes of A. niger grown on starch for 14, 21, and 28 days, where orange = 14 days, purple = 21 days, and green = 28 days.
GNPS obtained from ESI(−) also revealed the presence of polyhydroxylated fatty acids, a fatty diacid, and two aliphatic polyketides. In addition to two naphthopyrones, a naphthopyranone, a bicoumarin, an isocoumarin, and a terphenyl derivative were also characterized. A macrolide with a structure related to the five others was found.
The growth of a microorganism depends on the nutritional value of the culture medium, alongside other factors, such as pH, temperature, time of cultivation, and solid/liquid conditions. Therefore, the modification of one of these parameters can affect or promote growth with variation in metabolite profiles.?
Banana peels and starch media yielded seven identical compounds, namely, similanpyrone B (m/z 205.0503), orlandin (m/z 409.0921), thelephoric acid (m/z 351.0138), hexylitaconic acid (m/z 213.1129), fonsecinone B (m/z 589.1703), aurasperone A (m/z 571.1610), and Psiaα (m/z 293.2112). Only three compounds (similanpyrone B, orlandin, and hexylitaconic acid) were simultaneously found in starch-enriched banana peel and starch media.
Comparison of secondary metabolite profiles from both culture media revealed that BP_An medium produces more chemomarkers (27 compounds) from A. niger than the starch-enriched medium (22 compounds) (Figures and ?). This latter promoted significantly the production of chemo-markers (19 compounds) already reported in the Aspergillus genus than BP_An medium (14 compounds). The increase in the carbohydrate amount was beneficial to produce more secondary metabolites (87 compounds in BPS_An against 66 in BP). The BPS_An medium generated 26 nonidentified (NI) compounds, while BP_An produced half of this number.
Illustration of the origin distribution across various species for the secondary metabolites produced in BP media (BP_An14d, BP_An21d, and BP_An28d).
Distribution of the secondary metabolites produced in BPS media (BPS_An14d, BPS_An21d, and BPS_An28d) among various species.
Part of the NI metabolites did not produce sufficient fragment ions, enabling their characterization, and others could not be matched either with a known compound or a class of compounds. NI compounds might be produced after the activation of silent gene expression. Also, during the study, four triterpenes were detected, among them two with m/z 457.3679 (14, 21, and 28 days) and two with m/z 425.3776 (14 and 28 days). These compounds could be biotransformation products of the triterpene cycloeucalenone found in banana peels.
As shown in Tables–?, some of the compounds from the cultures are produced by A. niger for the first time, although already identified in other microorganisms. The variation in these chemical profiles suggests that the studied A. niger may have undergone gene alteration. Fungi generate an array of natural compounds, and these products can change if the fungi undergo mutations caused by stress. An earlier study showed that sodium azide induced mutagenic changes in A. niger, which were monitored using RAPD-PCR analysis.? One of the strategies to mutate these genes and promote one-strain-many-compounds (OSMAC) is to systematically change the culture conditions. ?,? It is also noteworthy to point out that banana peel secondary metabolites could interfere with gene expression or deletion. Isoorientin, a flavonoid from banana peels, possesses antimicrobial activity and is known to cause mitochondrial dysfunction in cancer cells. ?,? Thus, its presence in the culture medium could compel the fungus to develop adaptive mechanisms that include gene expression. The second flavonoid, eriodictyol-6-C-β-d-glucopyranoside, was not studied for its antifungal activity. However, previous work suggested that flavonoids are part of the plant metabolites produced in response to fungal infection.? In addition, banana peel extract from an incompletely identified Musa species showed antifungal activity by inhibiting 72% of A. niger growth.?
The present work used a food residue (banana peels) as a biomass to culture A. niger. During the investigation, some metabolites were produced, as depicted in Tables–?. Part of these metabolites showed pharmacological properties in previous studies. Aurasperone displayed IC_50_ of 12.25 μM and antiviral activity against SARS-CoV-2.? It also presents a high selectivity index of 2641.5. This compound also proved to be an antioxidant and was shown to possess a protective effect on CHO cell lines against H_2_O_2_.? Hexylitaconic acid inhibited with IC_50_ of 50 μg/mL the human double minute 2 (HDM2), a protein overexpressed in many human cancers that interferes with the tumor-suppressing functionality of p53.? This compound also promoted up to 13 and 66.3% of the germination and biomass in cauliflower seedlings.? Pyranonigrin A showed anti-inflammatory activity by reducing the level of NO production. ?,?
Banana peel is rich in nutrients, including minerals, carbohydrates (xylose, pectin, cellulose), amino acids, and fatty acids ?,? that could favor the growth of A. niger. However, it also contains secondary metabolites that possess antimicrobial activity, causing a strong stress to the fungus growth. ?,? The chemical profile of the peel’s crude extract showed the presence of secondary metabolites, namely, isoorientin, eriodictyol 6-C-β-d-glucopyranoside, hexadecasphinganine, 1,10-epoxy-8α-methoxyermophilanolide, and di(2-ethylhexyl)phthalate (Tables and ?). Isoorientin was found in an ethanolic extract of Mentha arvensis that displayed strong antibacterial activity against bacteria from the ESKAPE consortium.? In addition, this metabolite showed strong antimicrobial and antibiofilm activity against E. coli by interacting with the extracellular polymeric substances.? Flavonoids with an antibiofilm property also affected the A. flavus cell wall by causing osmotic stress as shown in a previous report. In response to this stress, the fungus increased its production of ergosterol.? In addition, A. flavus exposure to d-glucal led to the suppression of aflatoxin biosynthesis and greatly increased kojic acid production.? The stress induced by banana peels’ secondary metabolites might explain the presence of wall cell metabolites such as cholesterol, polyhydroxylated fatty acid derivatives, and compounds not previously found in the studied fungus.
The starch-enriched banana peel medium yielded more metabolites, with 28 days of fermentation proving to be the optimal condition. Sabat and co-workers similarly found antifungal profiles against Verticillium wilt of Penicillium steckii extracts produced on different carbohydrate-enriched media.? Vandermolen and co-workers also demonstrated that three fungal strains presented different secondary metabolite profiles after growing them separately on rice, grits, oatmeal, and wheat germ flour.? Other previous studies revealed that to overcome stress caused by pH, nutrients, temperature, osmosis, and oxidative agents, fungi can increase, suppress, and promote the synthesis of some secondary metabolites. ?,?
Overall, the presence of banana peels in BP and BPS media seemed to increase the production of fungal secondary metabolites. Banana peels, secondary metabolites, seem to play a key role in the expression of genes that regulate several biosynthesis pathways.
Conclusion
4
Suitable management of food residue and waste can bring numerous beneficial outcomes, such as renewable energy and valuable biomolecules with applications in drug discovery, veterinary medicine, and agriculture. Thus, the present study aimed to unveil biomolecules produced by A. niger when grown on banana peels (BP) used as biomass. The study also enriched BP (BPS) with starch to understand the chemical changes that can occur when the fungus grows in this medium. The combination of in silico statistical tools (metaboanalyst) and databases for dereplication (GNPS, MS-FINDER, and MS-DIAL) enabled us to establish and compare the chemical profile according to cultivation periods (14, 21, and 28 days) and culture medium. This unprecedented study reveals that the chemical profiles of A. niger grown on BP and BPS were composed of 76 and 99 substances, respectively. Alongside banana peel metabolites, polyketides, cyclopeptides, citric acid derivatives, alkamides, and sesquiterpenes were characterized, among others. Statistical analysis revealed that banana peel medium can produce secondary metabolites with pharmacological potential independent of the fermentation period. However, the starch-enriched banana peel medium optimally produced fungal compounds during the 28-day fermentation.
The present study provides a cheap process to induce and modulate fungal valuable molecule production and a method to turn waste into valuable products.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1United Nations Environment Programme (2024) An inside look Brazil’s push to end food waste. https://www.unep.org/news-and-stories/story/inside-look-brazils-push-end-food-waste. Accessed in April 2025.
- 2United Nations Environment Programme (2024) Food Waste Index Report 2024 . Think Eat Save: Tracking Progress to Halve Global Food Waste. Food Waste Index Report 2024 | UNEP - UN Environment Programme . Accessed in April 2025.
- 3Instituto Brasileiro de Geografia e Estatística (IBGE), Produção de banana 2024 . Online access: https://www.ibge.gov.br/explica/producao-agropecuaria/banana/br. Accessed in November 2025.
- 4Ribeiro G. R.Steinmetz R. L. R.Wisbeck E.Biogas production from solid residues generated in bioethanol production using banana biomass Rev. Bras. Eng. Agríc. Ambient.2025293 e 28235510.1590/1807-1929/agriambi.v 29n 3e 282355 · doi ↗
- 5Schieber A.Side Streams of Plant Food Processing As a Source of Valuable Compounds: Selected Examples Annu. Rev. Food Sci. Technol.201789711210.1146/annurev-food-030216-03013528068488 · doi ↗ · pubmed ↗
- 6Parra-Pacheco B.Cruz-Moreno B. A.Aguirre-Becerra H.García-Trejo J. F.Feregrino-Pérez A. A.Bioactive Compounds from Organic Waste Molecules 20242910224310.3390/molecules 2910224338792105 PMC 11123749 · doi ↗ · pubmed ↗
- 7Hikal W. M.Said-Al Ahl H. A. H.Bratovcic A.Tkachenko K. G.Sharifi-Rad J.KačániováM.Elhourri M.Atanassova M.Banana Peels: A Waste Treasure for Human Being Evidence-based complementary and alternative medicine: e CAM 20222022761645210.1155/2022/761645235600962 PMC 9122687 · doi ↗ · pubmed ↗
- 8Wani K. M.Dhanya M.Unlocking the potential of banana peel bioactives: extraction methods, benefits, and industrial applications Discover Food 2025512510.1007/s 44187-025-00276-y · doi ↗