Mycobacteria isolated from temperate stony corals
Dana Ulanova, Takuma Mezaki, Satoshi Kubota

TL;DR
This study isolates actinobacteria from temperate corals and finds that many belong to the Mycobacteriaceae family, suggesting a common coral-microbe relationship.
Contribution
The novel contribution is the identification of mycolic acid-containing actinobacteria, especially Mycobacteriaceae, in temperate stony corals.
Findings
Over 60% of isolated actinobacteria belonged to mycolic acid-containing genera.
Mycobacteriaceae family members were frequently found in the coral samples.
The study supports the global association of actinobacteria with coral hosts.
Abstract
In marine environment, actinobacteria are widely distributed in water and sediments, and form symbiotic relationships with higher organisms. In this study, we isolated 49 actinobacterial strains from three temperate stony corals , Pocillopora damicornis , Acropora hyacinthus and Acropora muricata . More than 60% of obtained actinobacterial isolates belonged to mycolic acid-containing genera, particularly members of the family Mycobacteriaceae . Our results combined with the previous studies demonstrated that these actinobacteria are frequently associated with coral hosts worldwide.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
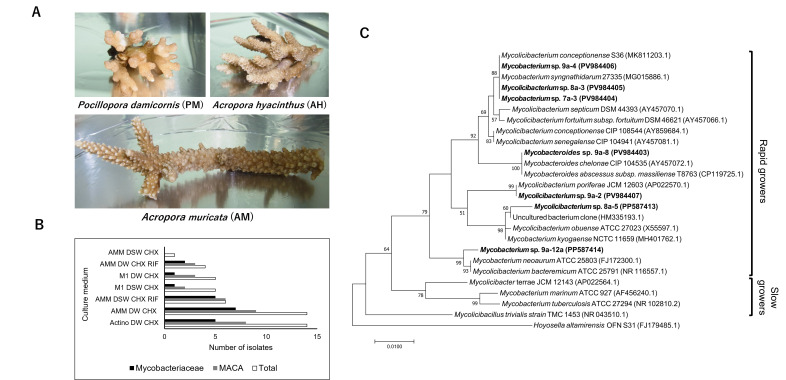 Figure 1
Figure 1|
Coral host |
Isolate name |
Accession number |
Isolation medium |
The closest BLAST hit |
e-value |
Query coverage |
Percent identity |
Accession number of the BLAST hit |
|
|
7a-1 |
|
Actino DW CHX |
|
0 |
100 |
100 |
|
|
7a-2 |
|
Actino DW CHX |
|
0 |
100 |
100 |
| |
|
7a-3 |
|
Actino DW CHX |
|
0 |
100 |
100 |
| |
|
7a-4b |
|
M1 DW CHX |
|
0 |
100 |
100 |
| |
|
7b-1a |
|
Actino DW CHX |
|
0 |
100 |
100 |
| |
|
|
8a-3 |
|
AMM DSW CHX RIF |
|
0 |
100 |
100 |
|
|
8a-5 |
|
AMM DW CHX |
Uncultured bacterium |
0 |
99 |
99.52 |
| |
|
|
0 |
100 |
99.41 |
| ||||
|
|
9a-2 |
|
AMM DW CHX RIF |
|
0 |
100 |
100 |
|
|
9a-3 |
|
AMM DW CHX RIF |
|
0 |
100 |
100 |
| |
|
9a-4 |
|
AMM DW CHX RIF |
|
0 |
100 |
100 |
| |
|
9a-5a |
|
Actino DW CHX |
|
0 |
99 |
100 |
| |
|
9a-7 |
|
Actino DW CHX |
|
0 |
100 |
100 |
| |
|
9a-8 |
|
Actino DW CHX |
|
0 |
100 |
100 |
| |
|
9a-10b |
|
AMM DW CHX |
|
0 |
100 |
99.88 |
| |
|
9a-12a |
|
AMM DW CHX |
|
0 |
100 |
99.38 |
| |
|
9a-24 |
|
M1 DSW CHX |
|
0 |
100 |
100 |
| |
|
9a-26 |
|
M1 DSW CHX |
|
0 |
100 |
99.52 |
| |
|
9a-30 |
|
AMM DSW CHX |
|
0 |
100 |
100 |
| |
|
9b-4a |
|
AMM DW CHX |
|
0 |
100 |
99.51 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Genomics and Phylogenetic Studies · Mycobacterium research and diagnosis
Description
Coral reefs are critical for the survival and functioning of marine organisms and ecosystems. Stony corals, the central component of reef ecosystem, are cnidarian organisms which have a close symbiotic relationship with diverse microorganisms, forming so-called coral holobiont. Coral-associated bacteria (CAB) were detected and isolated from all parts of the coral body (mucus, tissue and skeleton) and are known to provide coral hosts with nutrients, support coral larvae settlement and help to overcome stress conditions (Sweet et al., 2021; Voolstra et al., 2024). Most of the reported CAB belongs to *Pseudomonadota, Bacillota, Actinomycetota * and
- Bacteroidota* phyla (Sweet et al., 2021). While the abundance and function of several CAB, especially those from the phylum Pseudomonadota , have been described
- (* Rosado et al., 2018; Voolstra et al *., * 2024 ) , the roles of many others remain unclear.
Actinobacteria are Gram-positive bacteria well-known for producing bioactive compounds, which are widely used as antimicrobial, anthelmintic and anticancer drugs in human and animal medicine. These bacteria are found in both aquatic and terrestrial environments and many of them are associated with plant and animal hosts (van der Meij et al., 2017). Actinobacteria have been detected in soft and stony corals, mainly from tropical regions, and demonstrated to be a part of the core microbiome of several coral hosts (Hernandez-Agreda et al., 2018). Since actinobacteria are potent producers of bioactive compounds, they were proposed to have a role in protecting corals against pathogens (Kuang et al., 2015). Yet, the diversity, cultivability, and functional roles of coral-associated actinobacteria, particularly in non-tropical environments, are poorly understood. In this study, we aimed to address the knowledge gap regarding actinobacteria associated with corals from temperate regions. To achieve this, we isolated and identified actinobacteria from three stony coral species commonly found in the temperate marine environment of Kochi Prefecture, Japan: *Pocillopora damicornis * (PD), Acropora hyacinthus (AH) and *Acropora muricata * (AM).
In total, 49 actinobacteria-like colonies were isolated from fragments of three stony coral species ( Fig. 1A ). The highest number of colonies were isolated from AMM and Actinomycete isolation agars prepared with addition of distilled water and cycloheximide (CHX), and the lowest from AMM agar prepared using seawater and CHX ( Fig. 1B ). 16S rRNA gene sequence analysis classified bacterial isolates into 14 actinobacterial genera. The highest number of actinobacterial colonies (39) and genera (11) were obtained from *A. muricata * (Table 1). Interestingly, most of the isolated colonies (31) belonged to mycolic acid-containing actinobacteria (MACA): Gordonia (5), Rhodococcus (5), and representatives of the Mycobacteriaceae family (21).
Strains of the Mycobacteriaceae family, Mycobacterium , Mycolicibacterium or Mycobacteroides spp. (Gupta et al., 2018), were isolated from each coral host (Table 1). Based on 16S rRNA gene analysis, our isolates were closely related to the rapidly growing (forming visible colonies in less than 7 days) genera Mycolicibacterium spp. and Mycobacteroides spp., which are saprophytes widely distributed in nature (Brown-Elliot and Philley, 2017, Gupta et al., 2018) ( Fig. 1C ). The isolates related to Mycolicibacterium spp. strains were obtained from all coral hosts, while the later was isolated only from A. muricata . Interestingly, no slow-growing mycobacterial species (requering more than 7 days to form visible colonies (Gupta et al. 2018)) were detected in this study.
In this study, we used three media supplemented with distilled water or seawater, and two sets of antibiotics. Many marine bacteria, incl. actinobacteria, prefer seawater-based media for growth under the laboratory conditions. However, we obtained the highest number of actinobacterial isolates from distilled water-based media. One reason for this can be the overgrowth of seawater-based media by fast-growing marine bacteria, which form colonies within 1-2 days at 28°C. These fast growers outcompete actinobacterial colonies, which generally require more than two days to form. Rifampicin helps to suppress fast growers but inhibits some actinobacterial strains (Freel et al., 2012). Our results indicate that use of distilled water-based media may increase the isolation success rate of actinobacteria from marine environments.
Most of isolates from this study contain mycolic acids (MAs) - long fatty acids present in cell envelops, which provide protection to bacterial cells from environmental stresses (Dover et al., 2021). In mycobacteria, MAs were demonstrated to play a role in host immune system stimulation and biofilm formation (Marrakchi et al., 2014). Rhodococcus sp. and Gordonia sp., are known for their biodegradation and bioremediation properties (Van Der Geize and Dijkhuizen, 2004; Drzyzga, 2012). Such degradation capabilities may be used in coral-microbe symbiosis for the host protection from the pollutants present in the environment (Mahmoud and Kalendar, 2016). In addition, MACA often produce colored pigments, carotenoids, which protect cells from environmental stresses (Saubenova et al., 2024). Such pigmented bacteria may also provide protection to coral holobionts (Galasso et al., 2017).
Here, we newly report a number of rapidly growing Mycobacteriaceae strains cultured from three species of temperate stony corals. Mycobacteria were isolated from several tropical coral species (Li et al., 2014, 2022; Siro et al., 2022) and found to present one of core microbiome members in some corals (Hernandez-Agreda et al., 2018). Similarly to our study, Li et al. (2014) isolated fast-growing mycobacteria from Porites lutea samples collected at Luhuitou fringing reef. A report on oil-degrading bacteria from corals inhabiting oil-polluted areas of the Arabian Gulf, described the Mycobacterium sp. strain with crude oil-degrading ability from mucus sample of Acropora clathrata (Al-Dahash and Mahmoud, 2013). The authors suggested that this bacterium may participate in the protection of coral from oil pollution together with other oil-degrading bacteria. However, details on the phylogeny and function of the *Mycobacterium * genus in coral health and/or disease remain to be understood.
In conclusion, this study provides new insights into the diversity and distribution of coral-associated mycobacteria from temperate marine environments. Our results indicate that these actinobacteria may have a global distribution across different climate zones. This study is, however, limited to a few samples and more culture-dependent and independent research needs to be done to clarify the role of mycobacteria in coral hosts.
Table 1. Representative bacterial isolates from each coral host obtained in this study and their closest BLAST hits
**: **
Methods
Sample collection and bacterial isolation
Coral samples were collected on October 2019 by SCUBA diving offshore Nishidomari, Otsuki town, Kochi (32°46'41.0"N 132°43'54.9"E, water depth 3 m, temperature 25°C). The collected fragments were stored in local seawater at ambient temperature and processed on the next day of collection. Coral fragments were washed twice in sterile seawater to remove loosely attached bacteria and crushed in an alcohol-sterilized mortar. The obtained slurry was directly plated on following agar media: 1) AMM (Jensen et al., 2005) prepared using distilled water (DW) or deep seawater (DSW), supplemented with CHX (100 mg L ^-1^ ) to suppress fungal growth; 2) AMM prepared using DW or DSW, supplemented with CHX (100 mg L ^-1^ ) and rifampicin (RIF, 5 mg L ^-1^ ) to suppress fungal and fast-growing bacteria growth; 3) M1 (10x diluted AMM) prepared using DW or DSW, supplemented with CHX (100 mg L ^-1^ ); and 4) DIFCO ^TM^ Actinomycete isolation agar (BD) (Actino) prepared using DW and supplemented with CHX (100 mg L ^-1^ ). Inoculated plates were cultivated at 28°C for up to one month. Actinomycete-like colonies were transferred to fresh culture plates of the same medium until pure cultures were obtained. Colonies were considered to be actinobacteria-like if spores and/or aerial mycelium were formed (sporulating actinobacteria (e.g., Streptomyces )) or colonies had dry or waxy appearance with pink, light or dark-yellow coloration (e.g., Mycobacterium , Rhodococcus ).
Strain identification
The 16S rRNA gene amplification reaction mixture contained 1× EmeraldAmp Max PCR master mix (TaKaRa) and 0.3 μM of 16Seu27F (Čermák et al., 2008) and 1492R (Lane D.J., 1991) primers. A bacterial colony was added to the mixture by a toothpick. The cycling condition used was initial denaturation at 95 °C for 2 min, followed by 30 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 90 s. DNA sequencing was performed by ABI PRISM™ 3100 Genetic Analyser, according to manufacturer’s recommendations. Sequences were processed using Geneious sequence analysis software ver. 5.5.4 (Kearse et al., 2012) and analyzed using the BLAST program (Altschul et al . , 1990) (blastn, nucleotide collection database, as on April 1 ^st^ , 2024). The 97% percent identity and 90% query coverage were used as cutoffs when evaluating the blastn hits. Phylogenetic trees were constructed using MEGA7 software (Kumar et al., 2016).
16S rRNA gene sequences of the representative isolates were deposited in the GenBank under accession numbers PP587411- PP587415 and PV984398-PV984411.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al-Dahash Lulwa M. Mahmoud Huda M. 201371 Harboring oil-degrading bacteria: A potential mechanism of adaptation and survival in corals inhabiting oil-contaminated reefs Marine Pollution Bulletin 7220025-326X 36437410.1016/j.marpolbul.2012.08.02923014479 · doi ↗ · pubmed ↗
- 2Altschul Stephen F. Gish Warren Miller Webb Myers Eugene W. Lipman David J. 1990101 Basic local alignment search tool Journal of Molecular Biology 21530022-283640341010.1016/s 0022-2836(05)80360-22231712 · doi ↗ · pubmed ↗
- 3Brown-Elliott Barbara A. Philley Julie V. 2017224 Rapidly Growing Mycobacteria Microbiology Spectrum 512165-049710.1128/microbiolspec.tnmi 7-0027-2016 PMC 1168746028084211 · doi ↗ · pubmed ↗
- 4Čermák Ladislav KopeckýJan NovotnáJitka Omelka Marek Parkhomenko Natalya PlháčkováKamila Ságová-MarečkováMarkéta 2008101 Bacterial communities of two contrasting soils reacted differently to lincomycin treatment Applied Soil Ecology 4020929-139334835810.1016/j.apsoil.2008.06.001 · doi ↗
- 5Dover Lynn G. Thompson Amy R. Sutcliffe Iain C. Sangal Vartul 20211223 Phylogenomic Reappraisal of Fatty Acid Biosynthesis, Mycolic Acid Biosynthesis and Clinical Relevance Among Members of the Genus Corynebacterium Frontiers in Microbiology 121664-302X 10.3389/fmicb.2021.802532 PMC 873373635003033 · doi ↗ · pubmed ↗
- 6Drzyzga Oliver 201253 The strengths and weaknesses of Gordonia : A review of an emerging genus with increasing biotechnological potential Critical Reviews in Microbiology 3841040-841X 30031610.3109/1040841 x.2012.66813422551505 · doi ↗ · pubmed ↗
- 7Freel Kelle C. Edlund Anna Jensen Paul R. 20111128 Microdiversity and evidence for high dispersal rates in the marine actinomycete ‘ Salinispora pacifica ’ Environmental Microbiology 1421462-291248049310.1111/j.1462-2920.2011.02641.x 22117917 PMC 3272088 · doi ↗ · pubmed ↗
- 8Galasso Christian Corinaldesi Cinzia Sansone Clementina 20171123 Carotenoids from Marine Organisms: Biological Functions and Industrial Applications Antioxidants 642076-3921969610.3390/antiox 604009629168774 PMC 5745506 · doi ↗ · pubmed ↗