Revisions to the Gliophorus irrigatus complex (Agaricales, Hygrophoraceae, Gliophorus, section Unguinosae) including four new species, one new combination and comparisons of basidiome vs. eDNA distributions

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1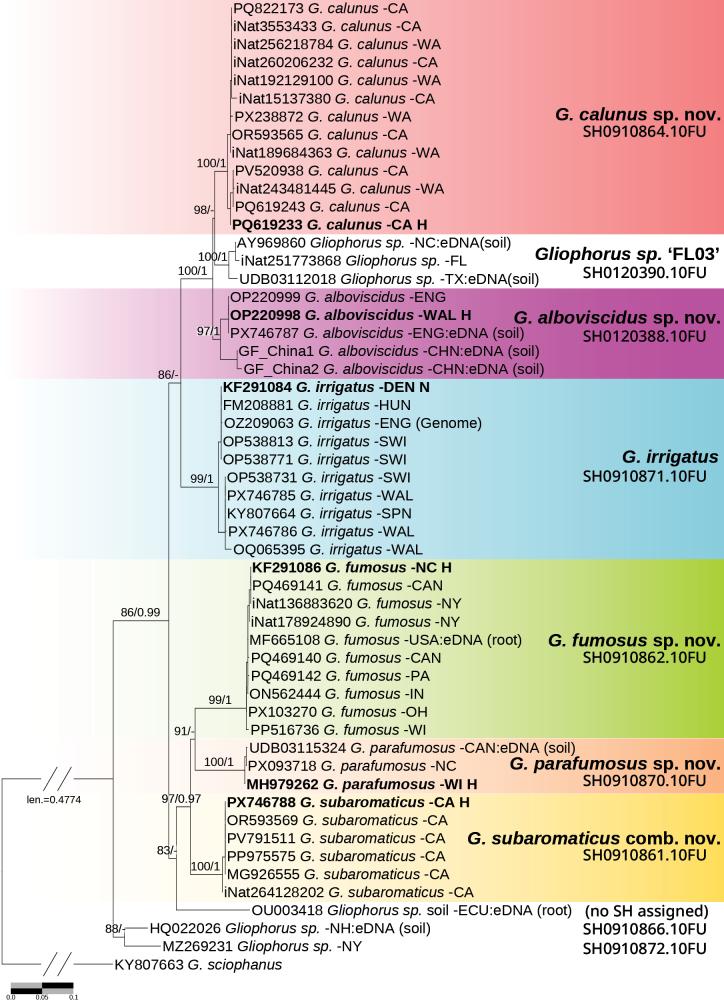 Figure 2
Figure 2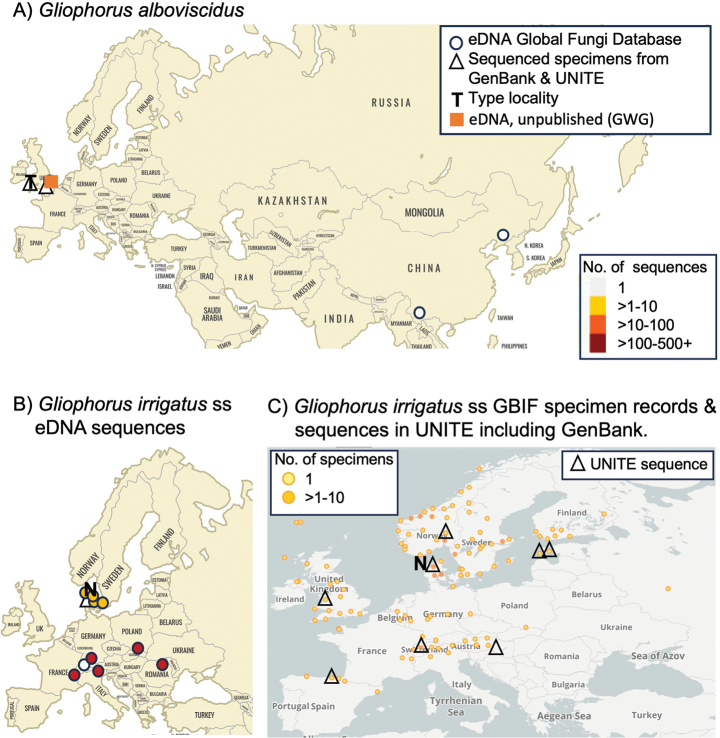 Figure 3
Figure 3 Figure 4
Figure 4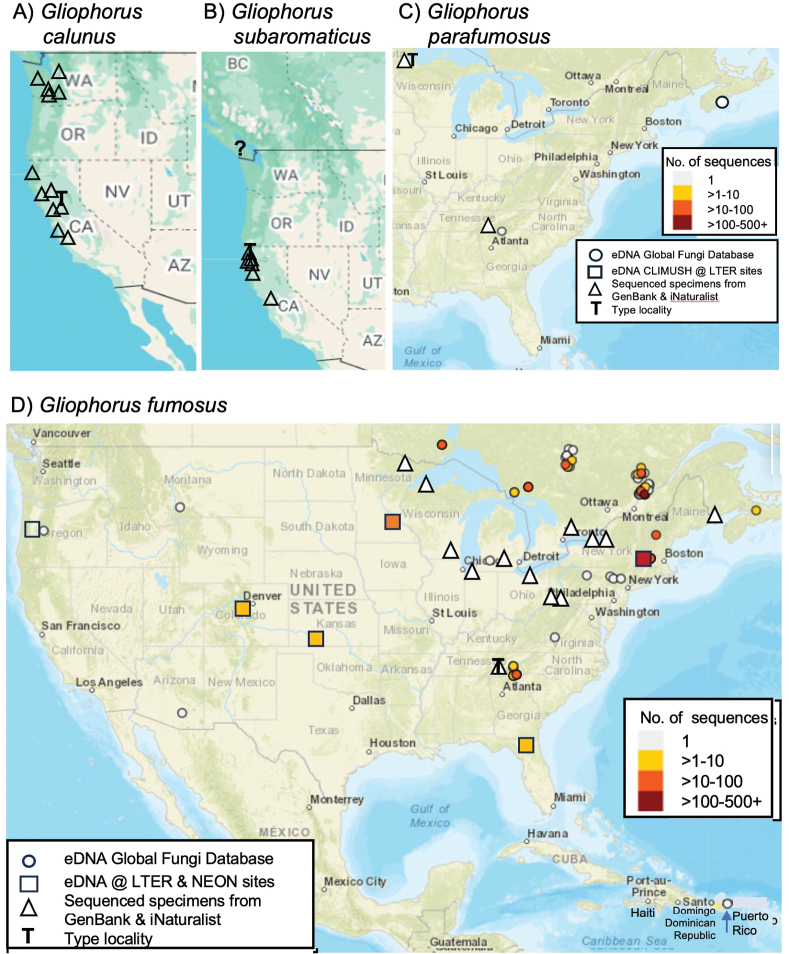 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9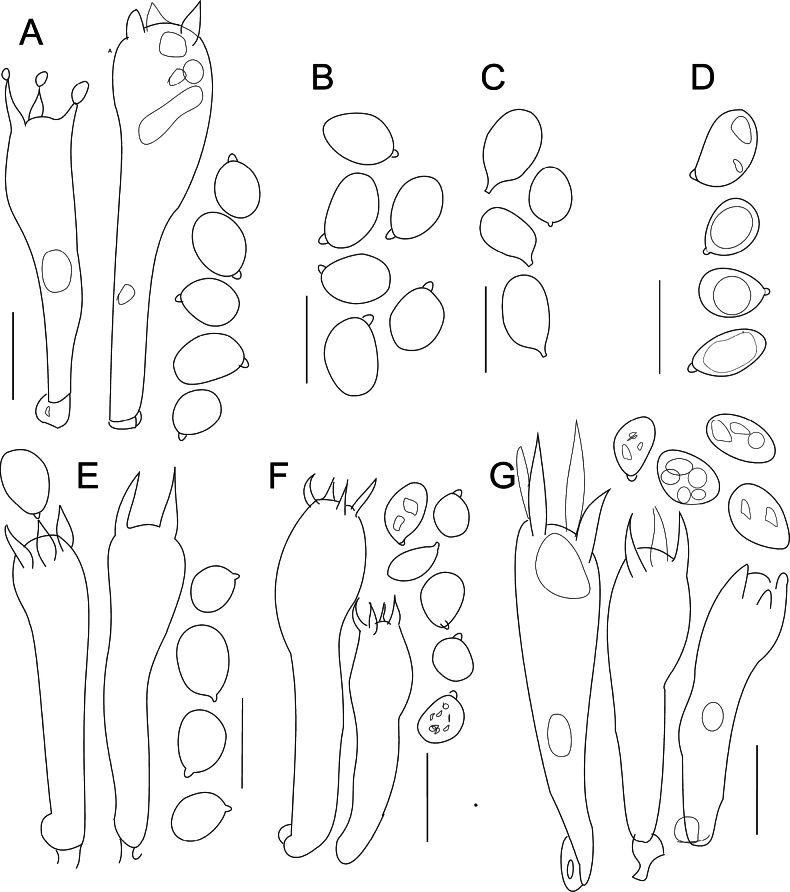 Figure 10
Figure 10| No. samples (No. sequence variants) at each % identity threshold | Distribution | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Region | SH (1.5%) | GenBank | 100% | 99.5% | 99% | 98% | 97% | 96% | 93% | Sequences >93% ID (>97% ID in red/bold) |
|
| ITS1 | SH0910869.1OFU |
| 0 | 0 | 0 | 0 | 0 | 0 |
| China, E. USA (NY.TX) |
|
| ITS1 | SH0910871.1OFU |
| 7(6) | 14(165) | 15(242) | 15(272) | 18(338) | 20(350) |
|
|
|
| ITS1 | SH0910862.1OFU |
| 10(3) | 14(97) | 15(278) | 15(386) | 15(387) | 15(387) |
| |
|
| ITS1 | SH0910870.1OFU |
| 4(1) | 4(2) | 4(2) | 4(2) | 4(2) | 4(2) |
|
|
|
| ITS1 | SH0910861.1OFU | ZAP119 | 0 | 0 | 0 | 0 | 0 | 3(4) |
| Nepal, SW China |
|
| ITS1 | SH0910864.1OFU |
| 0 | 0 | 0 | 0 | 0 | 0 |
| China, E. USA (NY,TX) |
|
| ITS2 | SH0910869.1OFU |
| 0 | 0 | 0 | 0 | 8(2)* |
|
| |
|
| ITS1 | SH0910871.10FU |
| 23(16) | 27(449) |
|
| ||||
|
| ITS2 | SH0910862.1OFU |
| 78(21) |
|
| |||||
|
| ITS2 | SH0910870.1OFU |
| 5(3) | 9(57) | 65(85) |
|
| |||
|
| ITS2 | SH0910861.1OFU | ZAP119 | 0 | 0 | 0 | 0 | 0 |
|
| S. China |
|
| ITS2 | SH0910864.1OFU |
| 0 | 0 | 0 | 0 | 0 |
|
| China, Canada |
| 1 | Pileus pale, white with Buff tint or pale Drab |
|
| – | Pileus strongly pigmented, brown or greyish-brown at least in centre |
|
| 2 | Distributed in the UK, possibly Eurasian based on eDNA (pale forms of |
|
| – | Distributed in north-western north America |
|
| 3 | Lamellae blue under UVf365 nm, lacking any odour |
|
| – | Lamellae white under UVf365 nm, under coastal redwood and often with unpleasant burnt plastic or rubber odour |
|
| 4 | Distributed in Europe and the UK |
|
| – | Distributed in eastern North America |
|
| 5 | Pileus centre dusky grey-brown or fuscous, lamellae broad |
|
| – | Pileus Drab Grey with darker striae to disc, faintly sulcate-striate to warm brown umbo; lamellae narrow, white; stipe long, slender (SH0910866.10FU; ITS: |
|
| 6 | Pileus margin straight when young, translucent-striate, rarely sulcate-striate, lamellae blue under UVf365 nm, mean spore length 6.7 µm |
|
| – | Pileus margin often inrolled and sulcate-striate; UVf365 nm unknown, mean spore length shorter, 6.4 µm |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications · Plant Diversity and Evolution
Introduction
In the revision of the higher-order classification in the family Hygrophoraceae by Lodge et al. (2014), Gliophorus was classified in Tribe Humidicuteae which is basal to Tribe Hygrocybeae and distal to Tribe Chromosereae in subfamily Hygrocyboideae. Lodge et al. (2014) only found Gliophorus appearing as a monophyletic clade in their 4-gene backbone ML analysis (ITS, LSU, SSU and rpb2), but bootstrap support was weak and not significant. Vizzini et al. (2012) also found that bootstrap and Bayesian support for a monophyletic Gliophorus were very low and not statistically significant. Both the 4-gene ML and Bayesian analyses by Lodge et al. (2014) focused on Tribe Humidicuteae and showed Gliophorus as a grade that was basal to Porpolomopsis and Humidicutis.
Chakraborty et al. (2018) did not find monophyletic support for the genus Gliophorus using only the ITS region, but they did find support for a monophyletic section Unguinosae using only two species (G. irrigatus s.s. from Denmark and the new species described here, G. fumosus from the USA), as well as a monophyletic section Glutinosae. However, section Gliophorus, which includes the type species, G. psittacinus (Schaeff.) Herink was paraphyletic. He and Yang (2021) dated the divergence of Gliophorus psittacinus and Humidicutis sp. to about 45 million years ago, but they only analysed one species per genus.
ITS sequences of Gliophorus species within the same section are remarkably divergent (up to 20–30%) including sister species – rates typically associated with divergences between genera and families. The reason for such high sequence divergence in Gliophorus is unknown, but it also occurs in other genera in the Hygrophoraceae, including Lichenomphalia and Hygrocybe (Lodge et al. 2014). Based on multiple lines of evidence, Lodge et al. (2014) concluded that most genera in the Hygrophoraceae are biotrophic, a trait that might contribute to high rates of ITS sequence divergence.
This study began with the discovery of two pale collections of Gliophorus in the UK that were initially thought to be albino variants of Gliophorus irrigatus until ITS sequences proved otherwise.
Gliophorus irrigatus (Pers.) A.M. Ainsw. & P.M. Kirk 2013 (Agaricales, Hygrophoraceae) was first recognised by Persoon as Agaricus irrigatus in pine woodland (Persoon 1801), presumably near Leiden or Göttingen where he had been based up to 1801. It is likely that the specific name referred to the highly viscid pileus (“glutino adhaerens”). Fries (1821) also accepted and described A. irrigatus from pine woodland in “Germaniae” (reference to Persoon’s earlier discovery), as well as from grassland in “Smoland” (Småland, southern Sweden).
However, Fries also named a similar species Agaricus unguinosus (unguen = soft fatty or oily substance) from Scania, also in southern Sweden (Skåne), the latter considered by Persoon to be a variety of A. irrigatus (Persoon 1828). Whilst A. irrigatus clearly has priority, more recent publications have varied in their treatment of the two species. For instance, Arnolds (1990b), considered the two species to be distinct, whilst Kovalenko (1988) mentions only A. unguinosus. However, Boertmann (1995, 2010) stated that the less viscid pileus of A. irrigatus was simply the result of ‘desiccation and erosion’, noting also that A. irrigatus has priority. He later formally synonymised Hygrocybe unguinosus into H. irrigata (Boertmann 2002).
The genus Gliophorus was created by Herink (1959) who also recognised only G. unguinosus and created Sect Unguinosae to accommodate this species. However, Herink’s renaming of G. unguinosus was invalid (nom. invalid, Art. 41.4), later corrected by Kovalenko (1988). Currently Gliophorus irrigatus (Pers.) A.M. Ainsw. & P.M. Kirk 2013 (IF/MB#550234) is the accepted name (Kirk 2013), but the section name remains valid.
Gliophorus irrigatus is reported globally, with most records from Europe (Boertmann 2010) and North America (Bessette et al. 2012), but records based only on morphology also exist from Australia (Young 2005), Asia (Japan) and Africa (Kenya) (https://www.gbif.org/species/8097905). In Europe, it is most commonly found in grassland habitats, but elsewhere in broadleaf and coniferous woodlands (Griffith et al. 2013; Halbwachs et al. 2018).
Upon finding that ITS sequences of pale G. irrigatus-like collections from Wales diverged strongly from G. irrigatus in Europe, the authors examined a similar, pale Buff to white species originally described as Hygrophorus unguinosus var. subaromaticus (Hesler and Smith 1942), but later renamed as Hygrophorus subaromaticus (A.H. Sm. & Hesler) Largent which was discovered in California under Sequoia (Largent 1985). It was distinctive in having a faint, but disagreeable odour and a near-absence of clamps on hyphae (though these are also rare in G. irrigatus). Apart from the type specimen from 1937, there are six records from northern California (GBIF:https://doi.org/10.15468/dl.9675az; Rockefeller: https://mushroomobserver.org/307719). The stipe/pileus colour of all of these ranged from pale Buff to white and two had an unpleasant (“plastic-like” or old rubber tyre) odour, one had a pleasant odour and the others had no odour. ITS sequences have been published in GenBank for four of the Californian specimens (MG926555, OR593569, PP975575, PV791511) that match the sequence we generated from the holotype (ITS:PX746788), one published in iNaturalist:264128202 and one in Mushroom Observer (307719), but these do not match sequences of the pale specimens from Wales.
Here, we describe a new white/pale Buff, viscid, odourless waxcap species from the UK that is very similar in appearance (except in colour) to G. irrigatus, but genetically quite distinct. We also describe two new species that resemble G. irrigatus morphologically for two of the grey-brown eastern North American clades that are phylogenetically distinct. We also show that the distributions of eDNA variants matching most of these species are wider than the distributions currently known, based on sequences obtained from basidiomes, but that G. irrigatus s.s., which we neotypify from Denmark, is restricted to Europe. Further, for two taxa restricted to north-western North America, we recombine Hygrophorus subaromaticus in Gliophorus and describe a new species, G. calunus, replacing the provisional name Gliophorus sp. ‘irrigatus-CA01’.
Materials and methods
Morphology
Macromorphology was assessed either when freshly collected or from photographs. Capitalised colour names are from Ridgway (1912) followed by Munsell (https://munsell.com/) colour annotations by Smithe (1975), while colour names in lower case are general descriptors.
Micromorphology of European collections was assessed with a Vickers M17 microscope using a 1.30 NA objective (oil immersion) with images captured using a Canon EOS system and analysed using MICAM v.1.6 (http://science4all.nl/). Micromorphology of eastern North American collections was assessed using an Olympus BH-2 microscope, an oil immersion lens (100×) and a drawing tube. A collection of G. calunus (iNaturalist:102000323) from Washington State was examined by L. Ré at 1000× with an oil immersion objective lens using a Nikon Eclipse Ci-L microscope; images captured using a Nikon Digital Sight DSFi-2. Two collections G. calunus from California HAY-F-012170 (holotype; iNaturalist:194601188) and HAY-F-000642 (iNaturalist:148430507) were examined by W. Cardimona at HAY using a Olympus BX53 microscope using an oil immersion objective lens.
DNA extraction and sequencing
DNA was extracted from voucher material using a quick extraction method (Zou et al. 2017) and amplified with the primer pairs ITS8-F (5’- AGTCGTAACAAGGTTTCCGTAGGTG-3’) / ITS6-R (5’-TTCCCGCTTCACTCGCAGT-3’) (Dentinger et al. 2010) for the ITS region or LR0R (5’-ACCCGCTGAACTTAAGC-3’) / LR5 (5’-TCCTGAGGGAAACTTCG-3’) using a Bentolab thermal cycler (https://www.bento.bio). Cycling conditions were as follows: for ITS, 95 °C for 12 min, 35 cycles [of 95 °C for 20 sec, 55 °C for 30 sec, 72 °C for 1 min] and a final extension step at 72 °C for 10 min.]; for LSU, 95 °C for 5 min, 35 cycles [of 95 °C for 30 sec, 50 °C for 30 sec, 72 °C for 1 min] and a final extension step [72 °C for 10 min]. Amplification of the ITS region from the holotype of G. subaromaticus was achieved using a new primer (SubaromF1 (5’-GAAGGATCATTAACTGAAATTTTAGGGA-3’, based on the sequence spanning the 18S/ITS1 border of GenBank:MG926555) in combination with ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) and using the same PCR cycling conditions as outlined for ITS amplification above. The resulting amplicons were sent for Sanger sequencing at the IBERS Translational Genomics Facility (Aberystwyth University). DNA of Gliophorus calunus iNaturalist:102000323 from WA, USA was extracted using a Qiagen DNeasy Plant Pro Kit, and amplified using a BioRad Thermalcycler at U. Wisconsin-LAX. DNA sequences and chromatograms were curated and assembled using Geneious Prime (https://www.geneious.com).
US samples that were sequenced by Mycota (Plymouth, Michigan, USA; https://www.mycota.com) used a MinION workflow designed by S. Russell for use with Flongle 10.4.1 flowcells and V14 Ligation chemistry (Russell 2023). A small (rice grain-sized) piece of dried basidiome was extracted with 20 µl X-Amp DNA reagent (IBI Scientific Cat. # B47441) in strip microtubes, heated for 1 hr at 80 °C in a thermocycler, followed by addition of 100 µm Molecular Biology Grade Water (IBI Scientific Cat. # IB42130). Reverse primers were uniquely tagged. Ten uniquely tagged forward (ITS1F) primers were used together with a set of 96 uniquely tagged reverse (ITS4) primers (Eurofins Genomics), designed for 96-well plates. Resulting reads were basecalled with Dorado (v.0 9.1), demultiplexed with Minibar (v.0.24) and consensus sequences were formed with NGSpeciesID (v.0.1.2.1). The reliability and accuracy of nanopore sequencing for amplicon-based barcoding has been broadly demonstrated (e.g. Koblmüller et al. (2024); Vasiljevic et al. (2021). It relies on consensus building as an error correction mechanism, producing results that, in many cases, match or even exceed Sanger accuracy (Srivathsan et al. 2021). Nanopore sequencing is now routinely employed in large-scale barcoding and biodiversity initiatives worldwide (Hebert et al. 2023).
Assembling ITS datasets
Sequences from GenBank and UNITE databases were initially identified via BLAST searches. Additional searches were undertaken for potentially related ITS1 or ITS2 sequences derived from soil eDNA metabarcoding studies via the GlobalFungi database (https://globalfungi.com/; Větrovský et al. (2020); Baldrian et al. (2021)) using release 5.0 (16.11.2023) of the database. Taxonomy for this version of the GlobalFungi database is based on UNITE version 10.0 (04.04.2024) (Abarenkov et al. 2024). For structured GlobalFungi searches, the following method was used: The “BLAST – group results (input 1 sequence; ITS1 or ITS2)” Search type was selected, to interrogate both the “Nonsingletons (all variants)” and “Singletons (annotated variants only)” databases. The search was selected to return the top 500 BLAST (sequence variant) results. The database was searched using either ITS1 or ITS2 sequences of the holo/neotypes of the six species considered in the present study. Thresholds for sequence identify were set from 93% to 100% identity and geographical distributions visualised via the ‘Map” tab (Table 1).
Additional eDNA sequences in GenBank were obtained from soil and litter collected at NEON and Long-Term Ecological Research sites and a few neighbouring parks in the USA (NCBI SRA under PRJNA1327291, USA National Science Foundation Grant DEB-2106130). For North America, a search of the GlobalFungi database, https://MycoBLAST.org and new eDNA data from US National Science Foundation Macrosystems Biology and NEON-Enabled Science grant (DEB-2106130), using American G. irrigatus-like reference sequences (MH979262 from Wisconsin and KF291086 from North Carolina) yielded 8221 eDNA sequences with > 97% identity to G. fumosus. These were used in mapping eDNA distributions. A 97% similarity for species identity was found to be useful for distinguishing species of Gliophorus, Lichenomphalia and Hygrocybe in the Hygrophoraceae (Lodge et al. 2014). Hughes et al. (2009) found that heterozygocity led to up to 3% divergence between the parental nuclear ribosomal ITS sequences in dikaryotic hyphae from a single basidiome. Hughes et al. (2013) found in several families of Agaricales that hybridisation between species in which viable progeny were produced was limited above 3%, but rarely occurred at up to 6% divergence between species.
Sequence alignment and phylogenetic analyses
Details of the sequences used for phylogenetic reconstruction in this study are listed in Suppl. material 1. The dataset containing the selected sequences was partitioned into ITS1, 5.8S and ITS2 regions. Each partition was aligned with MAFFT v.7.520 (Katoh and Standley 2013), using the G–INS–i algorithm. The final alignments were curated manually with AliView v.1.5 (Larsson 2014).
A Maximum-Likelihood (ML) phylogeny was inferred using IQ-TREE v.3.0.1. ModelFinder with greedy partition merging (-m MFP+MERGE) used to select the best-fit substitution model for each partition in the final scheme. Branch lengths followed the edge-linked proportional scheme (-p), i.e. partitions shared a single set of relative branch lengths. while each had its own partition-specific rate multiplier. Clade support was assessed with ultrafast bootstrap (UFBoot; 3,000 replicates with gene resampling (—sampling GENE) (Hoang et al. 2018) (Suppl. material 2).
Bayesian Inference (BI) was performed using MrBayes v.3.2.7 (Ronquist et al. 2012) using two independent Markov Chain Monte Carlo (MCMC) runs (four chains each, starting from random trees). The most appropriate evolutionary models and partitioning scheme were selected from IQ-TREE’s ModelFinder results. The chains were run for 10 million generations, sampling trees every 1000 generations. After discarding the first 25% of samples as burn-in, the remaining trees were summarised into a 50% majority-rule consensus and Bayesian posterior probabilities (BPP) were mapped to nodes. Convergence and mixing were evaluated by inspecting trace plots and effective sample sizes (ESS > 200) in Tracer v.1.7 (Rambaut et al. 2018) and by MrBayes diagnostics (potential scale reduction factors approx. 1.0). Nodes were considered strongly supported when SH-aLRT ≥ 80%, BPP ≥ 0.95 and/or UFBoot ≥ 95%.
Distribution mapping
Eurasian distributions of species in Gliophorus sect. Unguinosae were mapped using sequenced specimens of Gliophorus alboviscidus from this study and soil eDNA sequences from GlobalFungi Database matching at > 97% identity and mapped using Ultimaps (map of Eurasia with countries, cropped). ITS sequence of the neotype of G. irrigatus was used to find eDNA sequences matching at > 97% identity in the GlobalFungi Database (Větrovský et al. 2020) (Suppl. material 1) and mapped using Ultimaps (map of Europe with countries). Specimen-based records of preserved G. irrigatus having GIS coordinates were mapped using GBIF, with symbols added by hand indicating sequenced specimen records from UNITE, including GenBank records and the neotype. North American species in Gliophorus sect. Unguinosae were mapped, based on sequenced specimens reported in iNaturalist, Mushroom Observer and Mycoportal plus matching eDNA records at > 97% identity to soil sequences from US Long-Term Ecological Research sites (NCBI SRA under PRJNA1327291, USA National Science Foundation CLIMUSH grant), GenBank and soil eDNA sequence records from the GlobalFungi Database (Větrovský et al. 2020).
Results
Sample collection from the UK and morphological examination
In October 2021, a group of viscid, pale Buff waxcaps (tinted 1.0Y 6.0/7.0; Fig. 1A, B) was discovered on soil in unfertilised ancient grassland at Hundleton, Pembrokeshire, Wales. The morphology suggested the collection might be an albino form of a known species or an undescribed taxon. The viscid pale pileus was campanulate with Buff tints and a white margin and a white viscid stipe. The stipe was visibly marbled, a common feature of Gliophorus spp. The lamellae were decurrent and lacked a gelatinised edge. There was no perceptible odour. Microscopically, the specimen was consistent with the description of G. irrigatus. Thus, the macro- and micro-morphological characters were consistent with the concept of Gliophorus section Unguinosae Herink.
Macromorphology of Gliophorus alboviscidus sp. nov. basidiomes. DJH21 from Hundleton, Pembrokeshire (A, B) and CMB54 from Henfield, West Sussex (C, D).
Enquiries with members of the UK field mycology community revealed a report of a similar collection in 2019 from West Sussex (Fig. 1C, D), slightly larger and with a more strongly Buff-coloured pileus/stipe, which was also microscopically consistent with the description of G. irrigatus. The viscid pileus and stipe and marbled appearance of the stipe clearly placed this specimen in the genus Gliophorus.
Phylogenetic reconstruction
DNA sequences for the full ITS and D1/D2 domains LSU loci were obtained for both specimens (Suppl. material 1). Both were identical to each other and quite distinct from other sequences of G. irrigatus from both Europe and North America (< 91% ID across ITS region), thus excluding the possibility that they were simple albino variants.
Phylogenetic reconstruction, using sequence KY807663 (G. “sciophanus”) as the outgroup, placed these pale UK samples with high confidence in a distinct clade (Fig. 2). European specimens of G. irrigatus formed a well-supported clade, quite distinct from greyish-brown eastern North American specimens, as had long been suspected (D. Jean Lodge, pers. comm. 2021). In western North America, the Hygrophorus subaromaticus sequences with predominantly white or pale Buff specimens formed a fourth distinct clade, while a new species that included some pale Buff collections and provisionally named Gliophorus sp. ‘irrigatus-CA01’ formed a fifth clade. Thus, there are three unrelated clades with white to pale Buff basidiomes within the G. irrigatus species complex.
Maximum-likelihood tree of Gliophorus sect. Unguinosae, based on ITS sequences, with G. sciophanus as the outgroup. Support values on branches are ultrafast bootstrap (UFBoot)/Bayesian posterior probabilities (BPP) and are shown only for UFBoot ≥ 80 and BPP ≥ 0.90. Dashes (−) indicate values below the threshold. Holotypes (H) and the proposed neotype (N) for G. irrigatus s.s. are indicated. UNITE Species Hypotheses (SH) at 98.5% sequence identity are shown for each clade, except for G. alboviscidus and Gliophorus sp. ‘FL03’ for which a 3% SH threshold was applied. Scale bar: nucleotide substitutions per site.
Hygrophorus subaromaticus resembles G. alboviscidus in pileus colour, but belongs to a distant clade (Fig. 2). The holotype of Hygrophorus subaromaticus from redwood forest in north-western US was sequenced (PX746788) and this was 100% identical to four sequences in GenBank (MG926555, PV791511, OR593569 and PP975575; Fig. 9). Whereas the pileus colour of the type was described as buffy-brown on the disc and pale Olive-buff near the white margin, two of the recently sequenced collections were entirely white (MG926555, Mushroom Observer 307719 and OR593569, iNaturalist:148999043) and the others were white with a Buff disc (PP975575, iNaturalist:191417805) or Drab Grey on the disc or overall (PV791511, iNaturalist:256214946. No environmental sequences close to H. subaromaticus clade were detected either via GlobalFungi or UNITE (Fig. 5B), suggesting that this species is restricted to the northwest coast of North America.
Seven sequenced collections from redwood forest in northern California resemble H. subaromaticus with pileus colours ranging from nearly white to Buff or pale greyish-brown with a white margin, but the former comprises a distant clade provisionally named Gliophorus sp. ‘irrigatus-CA01’ and described below as Gliophorus calunus (Fig. 6). Five additional sequenced collections were identified from Washington State under other conifer species in coastal forests. Gliophorus calunus can be distinguished by the blue fluorescent lamellae under UVf365 nm, whereas H. subaromaticus lamellae remain white (refer to the key). The European clade of G. irrigatus s.s. is absent from North America, with most sequences derived from soils in Europe (Bahram et al. 2020; Praeg et al. 2020) (Fig. 3B).
Distribution of species in Gliophorus sect. Unguinosae in Eurasia. A. Sequenced specimens of Gliophorus alboviscidus from this study and soil eDNA sequences from GlobalFungi Database matching at > 97% identity; B. Previously sequenced specimen designated as the neotype of G. irrigatus and eDNA sequences from GlobalFungi Database (Větrovský et al. 2020) matching at > 97% identity; C. Specimen-based records of preserved G. irrigatus with GIS coordinates (GBIF.org; 11 Sep 2025) together with sequenced specimen records from UNITE, including GenBank records. A single occurrence of G. irrigatus in eastern Canada (Nova Scotia) was also detected via GlobalFungi, but in not mapped on Fig. 3B.
Taxonomy and distributions by region
The Gliophorus species named or renamed in this publication all belong to sect. Unguinosae Herink.
Eurasian species and distributions
Sequences falling into the G. alboviscidus clade (Fig. 2) were retrieved via GlobalFungi searches (ITS2 > 97% identity; none with ITS1 search) from two studies in China (Sun et al. 2015; Zhang et al. 2017) (Table 1; Fig. 3A). The G. alboviscidus clade is geographically structured, being comprised of branches with sequences from the UK and China. G. irrigatus s.s. was widely detected across Europe, as far east as Georgia, with ITS1 searches also detecting this species at low abundance in Newfoundland, Canada, the Faroe Islands, Denmark and the Canary Islands, Spain.
Gliophorus
alboviscidus
Taxon classificationFungiAgaricalesHygrophoraceae
D.J. Harries & G.W. Griff. sp. nov.
5B51B33F-6E90-5380-8CEA-182CBCA6963E
Index Fungorum: IF559911
Etymology.
Refers to the white colour and viscid nature of the basidiomata.
Diagnosis.
Basidiomata resembling Gliophorus irrigatus, but lacking grey or brownish colours. ITS sequences strongly divergent (9.7%).
Holotypus.
UK • Wales, Pembrokeshire, Somerton Farm near Hundleton, 51.6610, -4.9913, in undisturbed grazed grassland, 18 Oct 2021, D.J. Harries (voucher ABS:DJH21-29, [ABS = Aberystwyth University biorepository], holotype; K-M001434165, isotype) (GenBank ITS:OP220998; nrLSU:OP221000). UNITE/PlutoF:SH0910869.10FU (1.5% threshold).
Description.
Pileus 10–40 mm diam., convex to broadly campanulate, becoming applanate to subumbonate with age, margin faintly translucently striate to one third of disc, viscid, white with pale Buff tint (1.0Y 6.0/7.0) over the centre (Fig. 1A, B). Stipe 30–50 × 3–5 mm, cylindrical, hollow, viscid, white. Lamellae subdecurrent to decurrent, white. Taste and odour indistinct.
Basidiospores broadly ellipsoid or ovoid, (5.5–)6.0–8.0 × 4.5–6.0(–6.5) μm (mean 7.0 × 5.0), Q = 1.2–1.7 (mean 1.4) (type collection, 3 sporocarps, n = 70), spore print white. Basidia 40–50 × 5–8 μm, predominantly 4-spored. Cystidia absent on lamellae. Hymenophoral trama sub-regular. Pileipellis and stipitipellis an ixotrichoderm.
Habitat.
Recorded on soil in undisturbed grassland managed through cattle-grazing and hay cropping or of low nutrient status, regularly mown, amenity grassland, unploughed for over 50 years and not subjected to any synthetic fertiliser application.
Geographical distribution.
Basidiomes hitherto only observed in the UK, but eDNA data suggest a wider distribution across north-east Asia (Fig. 3A).
Other specimens examined.
UK • England, West Sussex, Henfield, St. Peter’s Church, 50.9325, -0.2765, in mown grassland (cemetery) on soil, 27 Sep 2019, Clare M. Blencowe, ABS: CMB54; K-M001434166), morphologically near-identical to holotype (Fig. 1C, D) (ITS:OP220999).
An exact match to the ITS2 sequence of this species was detected in soil eDNA (ITS2: PX746787) from ancient grassland at Hardwick Hall, Bury St. Edmunds, Suffolk, England (52.2294,0.7023; Griffith, unpublished data; Fig. 3A).
Notes.
By way of a common English name for this new species, we suggest pearlescent waxcap (Cap Cwyr Perlaidd in Welsh).
Neotypification of Gliophorus
irrigatus
In the absence of any voucher or drawing that might constitute a holotype, we propose the following:
Gliophorus
irrigatus
Taxon classificationFungiAgaricalesHygrophoraceae
(Pers.) A.M. Ainsw. & P.M. Kirk 2013 (Index Fungorum 23: 1, 2013)
7A8F2667-2097-52A2-9690-C66413CEEFFE
Index Fungorum: IF904305
Agaricus irrigatus Pers., Syn. meth. fung. (Göttingen) 2: 361 (1801). Basionym.
Neotypus.
Denmark • Northwest Jutland, Salling, Skive, 56,070, 09.135 in dry grassland grazed by sheep, 28 Oct 2006, David Boertmann (voucher CFMR:DEN-21 [2006/70], neotype selected here) (GenBank ITS:KF291084). UNITE/PlutoF:SH0910871.10FU (1.5% threshold).
Notes.
Persoon (1801) preserved the specimen which he described as Agaricus irrigatus. He did not provide collection details, but it is likely that the original specimen(s) was/were collected near Göttingen or Leiden. Neither Persoon’s (1801) original description of A. irrigatus nor Fries’ sanctioning citation (Fries 1821) for A. irrigatus contained any illustrations that could constitute a holotype. Enquiries about the possible existence of any relevant specimens or drawings from Christian Persoon confirmed that none was extant at Leiden (Jorinde Nuytinck, pers. comm. Oct 2024). Similar enquiries were made at Uppsala, but no paintings or vouchers survive (Åsa Kruys, pers. comm.).
At the Swedish Museum of Natural History, Stockholm there is a painting of A. unguinosus by Fries (from Sunnersta ‘skog’ [forest], in Uppsala; https://herbarium.nrm.se/specimens/S0627) (Mats Wedin [NRM], pers. comm., Oct 2024). However, consistent with the opinion of the authors and others, we find no evidence that would suggest that G. irrigatus and G. unguinosus are separate species. As G. irrigatus has priority, the later painting of A. unguinosus by Fries cannot be considered a type for G. irrigatus. A photograph of the neotype from Northwest Jutland in Denmark by D. Boertmann is shown in Fig. 4A and a similar collection by D. Harries in Pembrokeshire, Wales, UK (Fig. 4B).
Typical Gliophorus irrigatus s.s. basidiomes from Europe. A. G. irrigatus basidiomes, Neotype specimen CFMR:DEN-21 [2006/70], 28 Oct 2006, Denmark, Northwest Jutland, Salling, Skive, coll. David Boertmann, 56.92, 8.62 (ITS:KF291084) (courtesy of David Boertmann); B. G. irrigatus s.s. from Pembrokeshire, Wales, UK, D. Harries (ITS:OQ065395).
The ITS sequence of this specimen was used as the representative of ‘European’ G. irrigatus in phylogenetic analyses of Hygrophoraceae by Lodge et al. (2014) and was already designated the reference sequence for this species in UNITE.
Distribution.
Europe including the UK and southern Scandinavia according to eDNA sequences from the GlobalFungi database (Fig. 3B) and specimen records from GBIF and sequenced specimens in the UNITE database (Fig. 3C). Single occurrence of eDNA (singleton sequence) in far east of Canada.
North American species and distributions
Sequences from several grey-brown G. irrigatus-like specimens, all from North America did not clearly fall in either of the clades described above. In our phylogenetic analyses, two of these collected and sequenced as part of the Great Smoky Mountains National Park All-taxa Biological Inventory (Hughes et al. 2020) were placed close to the ‘North American’ irrigatus clade, but it is apparent that this clade comprises two main distinct clusters of sequences, one of which was provisionally named Gliophorus sp. ‘irrigatus-IN01’ and named below as Gliophorus fumosus sp. nov.; it appears to be quite widespread in the eastern US (e.g. GenBank: KF291086, ON562444, PP516736, PQ469140, PQ469142) and matching eDNA sequences were recovered across western parts of the USA as well (Fig. 5D). A second clade from the eastern USA and described below as Gliophorus parafumosus sp. nov. appears to be much less commonly found (MH979262 and PX093718; Fig. 5C). These appear strongly supported as sister clades of G. subaromaticus (ML 100; BPP 0.97) (Fig. 2). Additionally, a third taxon with grey-brown basidiomes was found in NY (iNaturalist:30847408; MZ269231), while a 91.8% similar eDNA sequence from NH remains unnamed.
A fourth North American clade with basidiomes varying from white to pale Buff, pale grey or shades of Drab (light to dark greyish-brown) was reported by the California Fungal Diversity Survey (CA FUNDIS) from coastal conifer forests in northern California provisionally named Gliophorus sp. ‘irrigatus-CA01’, (e.g. iNaturalist:194601188, ITS:PQ619233; also iNaturalist:148430507, ITS:OR593565) and also found in the Puget Sound area of northern Washington State (e.g. iNaturalist:189684363). This species is described below as Gliophorus calunus sp. nov.; it has a 95.9% sequence identity to the pale Buff-coloured G. alboviscidus from the UK despite their disjunct distributions. The fifth North American clade, Hygrophorus subaromaticus that we recombine in Gliophorus below, appears to be quite rare and hitherto recorded only from coastal redwood forests in northern California (Fig. 5B).
Distributions of species in Gliophorus sect. Unguinosae. A. Sequenced specimens of Gliophorus calunus; B. Sequenced specimens of Gliophorus subaromaticus; C. Sequenced specimens of Gliophorus parafumosus and two eDNA records from GlobalFungi Database (Větrovský et al. 2020); D. Sequenced specimens of Gliophorus fumosus and matching eDNA sequences at > 97% identity to soil sequences from USA NEON-LTER sites (PRJNA1327291) and soil eDNA sequence records from the GlobalFungi Database. G. calunus and G. subaromaticus were not detected in GlobalFungi database searches.
Macromorphology of Gliophorus calunus from northwest USA. A, B. Holotype HAY-F-012170 iNaturalist:194601188 (ITS:PQ619233), Damon Tighe: A. In sunlight; B. In UVf365 nm light. California, Medacino Co., near Fort Bragg, 21 Dec 2023. C, D. HAY-F-00642 iNaturalist:148430507), California, Humboldt Co., near Eureka. Mandy Hackney, Feb 2023; C. In sunlight; D. In UVf365 nm light.
Searches of the GlobalFungi database (Table 1), https://MycoBLAST.org and new eDNA data from US National Science Foundation Macrosystems Biology and NEON-Enabled Science grant (DEB-2106130) (Caiafa et al. 2025), with American G. irrigatus-like reference sequences (MH979262 from Wisconsin and KF291086 from North Carolina) yielded a larger number (8221) of matching sequences mostly linked to five studies from the USA and Canada with additional eastern North American records from iNaturalist, suggesting that this clade is restricted to North America (Fig. 5C, D).
Gliophorus
calunus
Taxon classificationFungiAgaricalesHygrophoraceae
L.A. Ré, D. Tighe, W. Cardimona, M. Hackney & Leal-Dutra sp. nov.
BBD6FD83-E83B-5519-BBB7-B8B59AC5BC51
Index Fungorum: IF904063
Etymology.
Cal – for California where the holotype and most of the additional collections were found by the California Fungal Diversity Survey and unus – Latin for one, referring to the number one in the provisional name for this species.
Diagnosis.
Pileus glutinous, margin usually inrolled, Pearl Gray, Light Drab to Drab, Dark Drab, Cinnamon or Tawny Olive at centre, usually paler towards margin. Stipe viscid with horizontal bands of contrasting translucence. Resembling Gliophorus subaromaticus from the same north-western North American coast to coastal range conifer forests, but differs in pileus centre usually with Drab tones vs. Buff to Olive-buff or buffy-brown tones, lamellae bright blue vs. white under UVf365 nm, pileus margin usually inrolled vs. straight and lacking an odour vs. usually with a disagreeable odour.
Holotypus.
USA • California, Mendocino Co., Jackson Demonstration State Forest, 39.4045, -123.7537 (accuracy 12 m), ca. 140 m a.s.l., 22 Dec 2023, D. Tighe, (voucher HAY:F-012170 [California State University East Bay Fungarium], iNaturalist:194601188, holotype) (GenBank ITS:PQ619233). UNITE/PlutoF:SH0910864.10FU (1.5% threshold).
Description.
Pileus 6–24 mm diam., convex to broadly convex with an incurved margin, becoming plane or nearly so, sometimes with an umbo, colour Pearl Gray in type, Light Drab to Drab, Cinnamon or Tawny Olive, usually paler towards margin, strongly viscid to glutinous especially when young, margin often translucent-striate. Stipe 20–60 × 3–9 mm, often concolorous with the pileus, slimy viscid, horizontal bands of contrasting translucence (marbled), glabrous, equal or slightly tapered at base, some slightly curved or nodulose; context hollow. Lamellae sinuate or slightly arcuate, white, sometimes tinted pale grey or drab, 1–2(–3) lengths lamellulae inserted, bright blue under UVf365 nm. Taste and odour indistinct.
Basidiospores hyaline, ellipsoid, (6.7–)7.5–9.6(–10.8) × (4.1–)4.4–5.5(–6.5) µm, mean Q = 1.7–1.8 in the holotype and iNaturalist:148430507 from northern California, (6.1–)6.5–8.6(–9.6) × (4.1–)4.3–5.2(–5.6) µm, mean Q = 1.6 in iNaturalist:1102000323 from Olympia in northern Washington. Basidia 4-sterigmate, with typical (non-toruloid) basal clamp connections. Cystidia absent on lamellae. Hymenophoral trama subregular. Pileipellis and stipitipellis an ixotrichoderm, with slender embedded hyphae lacking toruloid clamp connections.
Habitat.
Holotype growing on a mossy sandbank amongst grasses, sedges and cattails, near Vaccinium ovatum and Notholithocarpus densiflorus. Typically found in coastal Cupressaceae-dominated forests (coastal redwood, Sequoia sempervirens and western red cedar, Thuja plicata), often mixed with hardwoods and other conifers (western hemlock and Douglas fir).
Geographical distribution.
Occurs in coastal forests of western North America, recorded from northern California and western Washington State, USA, but likely occurring throughout coastal Cupressaceae-dominated forests in western North America, especially Thuja plicata. (Fig. 5A).
Additional specimens examined.
USA • (1) California, Humboldt Co., Eureka Area, Headwaters Forest Reserve, Headwaters Forest Trail, 40.6897, -124.1309, ca. 45 m a.s.l., under redwood (Sequoia) and alder (Alnus), Feb 2023, M. Hackney, iNaturalist:148430507, ITS:OR593565 (HAY-F-000642); (2) Washington, Thurston, County, N of Olympia, W of South Bay, 47.0738, -122.9012, ca. 10–20 m a.s.l., 22 Nov 2021, L. Ré & S. Hickey (iNaturalist:102000323).
Additional sequenced specimens with photographs.
USA • (1) California, Humboldt Co., Trinidad, Strawberry Rock Trail, 41.0790, -124.1313 (187 m accuracy), ca. 190 m a.s.l., 17 Nov 2023, N. Siegel CSALVA 37, northern Franciscan (coastal) redwood forest, (NS1093, UCSC); (2) Marin Co., Bolinas, Bolinas Fairfax Rd., 37.9404, -122.6577, ca. 45 m a.s.l., under coastal redwood (Sequoia sempervirens), Tanoak (Notholithocarpus densiflorus) and Vaccinium spp., 6 Jan 2024, D. Tighe, iNaturalist:195986180, ITS:PQ619243, (HAY-F-012180); (3) ibid., Mendocino Co., Jackson Demonstration State Forest, near Caspar Orchard Rd., 39.3606, -123.7787, ca. 110 m a.s.l., under coastal redwood (Sequoia sempervirens), western hemlock (Tsuga heterophylla), evergreen huckleberry (Vaccinium ovatum) and tanoak (N. densiflorus), 19 Jan 2025, D. Lyons, iNaturalist:259023907, ITS:PV520938 (HAY-F-013708); (4) ibid, San Mateo Co., SW of Redwood City, 37.4700, -122.27 (28.4 km accuracy), Jan 2025, Y.-M. Wang, iNaturalist:260206232; (5) ibid., Santa Cruz Co., Santa Cruz, Henry Colwell Redwoods State Park, 37.0195, -122.0436, ca. 160 m a.s.l., 13 Feb 2023, D. Lyons, iNaturalist:148743563, ITS:PQ822173 (OMDL11). (6) Washington, King Co., Auburn, 188^th^ Ave. SE, 47.2675, -122.0914, ca. 45 m a.s.l., 31 Oct 2023, Y.-M. Wang, iNaturalist:189684363 (PSMS1465); (7) ibid., Mason Co., near Little Skookum Inlet and SE Lynch Rd., 47.1480, -123.0630, ca. 10–20 m a.s.l., 2 Dec 2024, S. Ness, iNaturalist:253949042 (OLY-0918); (8) ibid., Thurston, County, N of Olympia, W of South Bay, 47.06976, -122.89716, ca. 20 m a.s.l., 25 Nov 2023, B. Funkhouser, iNaturalist:192129100; (9) ibid., Boston Harbor, near Boston Harbor Rd. NE and Bromley Ln NE, 47.13150, -122.89779, 8 m accuracy, ca. 10 m a.s.l., 25 Dec 2024, M. Koons, iNaturalist:256218784.
Notes.
Gliophorus calunus resembles G. subaromaticus, but differs in pileus centre usually with Drab tones vs. Buff to Olive-buff or buffy-brown tones, lamellae bright blue vs. white under UVf365 nm, pileus margin usually inrolled vs. straight and lacking an odour vs. usually with a disagreeable odour. Both species occur in north-western North American coastal forests dominated by Cupressaceae, with G. calunus growing from soil beneath both Sequoia sempervirens and Thuja plicata, whereas G. subaromaticus has hitherto only been recorded under S. sempervirens. G. calunus appears to be a more widespread and abundant species (Fig. 5A, B). The pileus of G. fumosus, hitherto only collected in eastern North America, but with eDNA sequences extending into western North America (Fig. 5D), is usually tinted more greyish-brown and differs in having a straight margin. Coastal conifer forests where G. calunus likely occurs in north-western North America are currently threatened by heavy logging. We suggest California Slimy Waxcap as the common English name for this new species.
Gliophorus
fumosus
Taxon classificationFungiAgaricalesHygrophoraceae
D.J. Lodge, S.D. Russell & Leal-Dutra sp. nov.
549E343B-7E9A-5B36-A0D0-1452B8CBF290
Index Fungorum: IF902428
Etymology.
Fumosus = smoke, referring to the type locality in the Great Smoky Mountains National Park and to the smoky colour of the stipe.
Diagnosis.
Basidiomata resembling Gliophorus irrigatus but basidiospores shorter (5.6–7.4 vs. 6.5–9) and hitherto recorded only in North America.
Holotypus.
USA • North Carolina, Haywood Co., Great Smoky Mt. National Park, Cataloochie Cove, near Rough Fork & Big Fork Ridge trails, 35.6164 -83.1208, in forest, 12 Aug 2005, E.B. Lickey & D.J. Lodge (voucher TENN-F-061868 [DJL05NC50], holotype) (GenBank ITS:KF291086). UNITE/PlutoF:SH0910862.10FU (1.5% threshold).
Macromorphology of Gliophorus fumosus basidiomes. A, B. Holotype of G. fumosus, DJL05NC50 from USA: North Carolina, Haywood County, Great Smoky Mountain National Park, Cataloochie, 12 Aug 2005, E.B. Lickey & D.J. Lodge (ITS:KF291086); C. USA, New York, Rome, 27 Sept 2022, D. McCheyne, iNaturalist:136883620 (ITS:DMc055); D. USA, Wisconsin, La Pointe, Big Bay State Park, 19 Sep 2023, Kyle Canan iNaturalist:183983484, (ITS:PP516736); E. Canada: New Brunswick, Queens Co., Canning, 8 Sep 2022, A. Justo, iNaturalist:134364174 (ITS:PQ469140).
Description.
Pileus 10–35 mm diam., shape variable amongst and within collections, narrowly to broadly campanulate with a mammalate umbo when young in the type and iNaturalist:145165934 and iNaturalist:178924890, often parabolic-umbonate or convex-hemispheric with a low umbo, becoming broadly parabolic or broadly convex with age with or without a low umbo, margin straight, translucently striate one third to two thirds to centre, glutinous or viscid, Drab (9.0 YR 5.5/2.5) or greyish-brown with darker striae and Dark Greyish-Brown (6.0 R 2.0/1.0) centre (Fig. 7). Stipe 30–60 × 2–5 mm, cylindrical, hollow, often tapered at base and flared at apex, rarely caespitose; surface Smoke Grey (2.5 Y 6.0/2.4 to 5.0 Y 7.0/2.0), drab grey or Drab (9.0 YR 5.5/2.5), glutinous to viscid, hyphae with small clamps. Lamellae 5–10 mm broad, sinuate with a small to large decurrent tooth, white, pale grey towards pileus, regular, 1–2 lengths of lamellulae inserted, edges concolorous, slightly wavy. Lamellae of iNaturalist:290749159 emitted an intense blue light under UVf365 nm. Taste and odour indistinct. Basidiospores broadly ellipsoid or ellipsoid, 5.6–7.2(–7.4) × 4–5.6 µm (mean 6.7 × 4.6), Q = 1.3–1.6(–1.7), (mean 1.5) (type collection n = 20). Basidia 35–82 × 7.2–15.2 µm, 4-spored with medallion clamps at base. Cystidia absent on lamellae. Hymenophoral trama subregular, comprised of cells 19.2–82 × 8.8–15.2 µm, gelatinisation absent, conducting elements abundant in context at the lamellar edge. Pileipellis and stipitipellis an ixotrichoderm.
Habitat.
Recorded on forest soil, with one eDNA record detected in Fagaceae roots (Arkansas, USA). iNaturalist:290749159 was collected between a bog and pond under Quercus sp. and Nyssa sylvatica, with abundant Vaccinieae in the understorey. Habitats covered in eDNA studies include prairie and alpine habitats extending to western USA in addition to eastern forests.
Geographical distribution.
Basidiomes known only from eastern North America, but eDNA data suggests distribution as far west as Arizona, Colorado and Oregon (Fig. 5D).
Other specimens examined.
USA • (1) Indiana, Porter Co., North Mineral Springs, Indiana Dunes National Lakeshore, Dune Acres, Cowles Bog Trail, 41.6495, -87.0744, 4 Jul 2017, S.D. Russell, MM6069 (PUL 00035147; F 20317), (ITS:ON562444); (2) Ohio, Summit Co., Green, Nimisila Reservoir Metro Park, 40.9369, -81.5155, 18 Jun 2025, Jessica Williams, 2025SMP20, iNaturalist:290749159 (ITS:PX103270), KEF015 as “Gliophorus ‘irrigatus-IN01’.
Additional records based on photographs and ITS sequences.
Sequenced photographic records posted on iNaturalist are all < 1% divergent from the holotype sequence (all within SH0910862.10FU), provisionally named ‘irrigatus-IN01’.
Canada • New Brunswick, Queens, Jemseg Grand Lake Watershed (45.9458, -66.1106, 8 Sep 2022, A. Justo [voucher at NBM]), iNaturalist:134364174, (ITS:PQ469140).
USA • (1) Wisconsin, Ashland Co., Apostle Islands LTA, Madeline Island, 46.7922, -90.6656, 19 Sep 2023, K. Canan [voucher OMDL01567]), iNaturalist:183983484, (ITS:PP516736); (2) Pennsylvania, Huntingdon Co., Alan Seeger Natural Area, 40.6944, -77.7552, 18 Aug 2023, J. Plischke, voucher JP23-0492, iNaturalist:193184758 (ITS:PQ469141); (3) Pennsylvania, Somerset Co., Sequanota Lutheran Conference Center and Camp Estate (40.17824, -79.10461, 2 Sep 2023, J. Plischke [voucher N23-1072]), iNaturalist:181314311 (ITS:PQ469142); (4) New York, Oswego Co., Pulaski, 43.5510, -76.2047, 17 Aug 2023, P. DeSanto, voucher MYCO-1000032 (CM23-80169), iNaturalist:178924890 (ITS:CM23-80169); (5) New York, Oneida Co., Rome, Sand Plains Unique Area (43.23238, -75.56605, 27 Sep 2022, D. McCheyne, voucher: MYCO-1000031, iNaturalist:136883620 (ITS:DMc055).
Additional environmental sequences.
USA: Arkansas, clone OTU_368, from (ectomycorrhizal) Fagaceae roots, (ITS2:MF665108). DNA of this species was detected in a range of woodland, prairie and alpine habitats in North America (Fig. 5D; 617 sequences deposited at NCBI SRA under PRJNA1327291).
Notes.
The macroscopic characters are not diagnostic for this species. Macroscopic characters such as the presence of an umbo varies amongst collections and amongst specimens within a collection and the microscopic characters do not vary distinctly amongst species in this complex. The basidiospores are slightly shorter than those of G. irrigatus s.s. from Europe as noted in the diagnosis, but slightly longer than spores of G. parafumosus. Microphotographs of basidia and basidiospores of J. Williams 2025SMP20 are posted on iNaturalist:290749159 (ITS:PX103270). ITS sequences, however, are diagnostic and differ by about 10% between the G. fumosus and G. parafumosus clades. The bright blue appearance of lamellae under UVf365 nm illumination is distinctive, but appearance of other eastern species under UV light is lacking. We suggest Smoky Waxcap as the common English name for this new species.
Gliophorus
parafumosus
Taxon classificationFungiAgaricalesHygrophoraceae
D.J. Lodge, S.D. Russell & G.W. Griff. sp. nov.
2757D973-FE15-57F7-B635-D27D8BFFAA59
Index Fungorum: IF904176
Etymology.
Parafumosus; para= close by and fumosus = smoke, referring to its phylogenetic proximity, as well as its similarity in both morphology and distribution to G. fumosus.
Holotypus.
USA • Wisconsin, Bayfield, Big Bay State Park, 46.7976, -90.6697, 8 Sep 2017, S. Wunderle, (voucher F:C0348314F [NAMA 2017-248] [F = Field Museum of Natural History biorepository]) (GenBank ITS:MH979262). UNITE/PlutoF:SH0910870.10FU (1.5% threshold).
Diagnosis.
Basidiomata similar macro- and microscopically to Gliophorus fumosus and having the same geographic range, but differing in ITS sequence and by having a darker pileus umbo (Burnt Umber to Fuscous vs. Warm Sepia to Dark Drab) and an inrolled rather than straight pileus margin.
Macromorphology of Gliophorus parafumosus basidiomes. A. G. parafumosus NC-91 (ITS:PX093718). USA: North Carolina, Swaine County, Great Smoky Mountain National Park, Mingas Mill, near Graveyard, (Lat/long: 35.5219, -83.3096; 360 m a.s.l.), 15 Aug 2005, D.J. Lodge; B. G. parafumosus holotype NAMA 2017-248 (ITS:MH979262). USA: Wisconsin, Bayfield, Big Bay State Park (Lat/long: 46.7976,-90.6697), 8 Sep 2017, S. Wunderle, Mycportal:C0348314F.
Description.
Pileus 12–35 mm diam., parabolic when young, becoming broadly parabolic with age, margin inrolled and slightly sulcate when young and less so with age, surface strongly viscid, colour Burnt Umber in centre when young, Fuscous with age, Drab on margin when young and overall with age. Stipe 25–75 × 3–8 mm, broader in lower 1/3, some compressed, hollow, surface strongly viscid, Smoke Grey (2.5 Y 6.0/2.4 to 5.0 Y 7.0/2.0) or tinted drab grey. Lamellae White or pale Cream Colour, sinuate or adnate with a decurrent tooth, 4–8 mm broad, margin wavy, one length of lamellulae inserted. Taste and odour indistinct. Basidiospores ovoid, hyaline, (5.2–)5.6–6.4(–8) × 4–5.2 µm, mean 6.4 +/- 0.76 × 4.6 +/- 0.39 µm, with or without guttulate contents, inamyloid. Basidia 4-sterigmate, 28–37 × 4.8–8 µm, with a small basal clamp. Subhymenium 10–12 µm deep, of interwoven hyphae 2.4–4.8 µm wide. Lamellar context subregular, comprised of swollen elements 30–80 × 8.8–18.5 µm rarely with small clamp connections; lamellar edge not gelatinised. Cystidia absent on lamellae. Pileus context of swollen hyphae 80–120 × 8–20 µm, with pale brown contents in potassium hydroxide (KOH) and small, rare clamp connections. Pileipellis an ixocutis, hyphae embedded in gelatinous matrix 1.2–2.4 µm, rarely with small clamps. Stipe context hyphae parallel, 5.6–16 µm diam. with pale brown contents, slightly constricted at septa, lacking clamp connections. Stipitipellis and ixotrichoderm, hyphae embedded in gelatinous matrix 1.2–3 µm diam. with few small clamps.
Habitat and distribution.
Hitherto recorded only in North America. Growing in moss in moist, mixed deciduous forests of eastern North America (Fig. 5C).
Other specimens examined.
USA • (1) North Carolina, Swaine County, Great Smoky Mountain National Park, Mingas Mill, near Graveyard, (35.5219404, -83.3096265) 360 m a.s.l., 15 Aug. 2005, D.J. Lodge, NC-91, (ITS:PX093718).
Additional environmental sequence.
Canada • Halifax, UNITE soil sample TUE002679, UDB03115324, 8 Nov 2019 (Tedersoo et al. 2014).
Notes.
Differs from G. fumosus in having a pileus margin that is distinctly inrolled and sulcate-striate, in contrast to G. fumosus (and G. irrigatus) has/have a straight or rarely slightly in-rolled gluten on the margin and is either not striate or only translucent-striate – not sulcate. Mean basidiospore length of G. parafumosus is slightly smaller than in G. fumosus (6.4 vs. 6.7 µm). Based on numbers of sequence-confirmed vouchers, G. parafumosus appears to be less common that G. fumosus. The microscopic observations were from the type collection deposited at F. The collection NC-91 was not annotated because it was immature and the remaining piece was not deposited at TENN after removing part for sequencing. We suggest False Smoky Waxcap as the common English name for this new species.
Gliophorus
subaromaticus
Taxon classificationFungiAgaricalesHygrophoraceae
(A.H. Sm. & Hesler) Rockefeller, G.W. Griff. & D.J. Harries comb. nov.
8242633D-FB51-58D3-9541-BEB49163B772
Index Fungorum: IF901873
Gliophorus subaromaticus (A.H. Sm. & Hesler) Rockefeller, G.W. Griff. & D.J. Harries, comb. nov.Hygrophorus unguinosus var. subaromaticus A.H. Sm. & Hesler, Lloydia 5(1): 81 (1942). Basionym.Hygrophorus subaromaticus (A.H. Sm. & Hesler) Largent, The Agaricales (Gilled Fungi) of California, 5. Hygrophoraceae (California): 106 (1985). Synonym.
Holotypus.
USA • California, Orick, Prairie Creek State Park, Nov 28 1937, (voucher MICH:10963 [Smith 9167]). Augmented here with ITS sequence (PX746788) of holotype: UNITE/ PlutoF:SH0910861.10FU (1.5% threshold).
Description.
Pileus 10–50 mm broad, convex to broadly convex with incurved margin, becoming plane or nearly so, sometimes with a low flattened umbo, colour “buffy-brown” on the disc, “pale olive-buff” near the whitish margin (a dull olive greyish-brown to pallid) in the type, pale Buff with or without a white margin in iNaturalist:256214946 (ITS:PV791511), white with pale Buff disc in iNaturalist:191417805 (ITS:PP975575) and pure white in both iNaturalist:9640562 (ITS:MG926555) and iNaturalist:148999043 (ITS:OR593569), glabrous slimy-viscid, margin striatulate in the type, but not in the recent collections. Context thin, very soft and fragile, whitish. Stipe 5–6 cm long, 6–10 mm thick concolorous with gills when fresh, but drying pale grey like the pileus in type, equal, hollow, fragile, slimy viscid, as in G. laetus, glabrous. Lamellae bluntly adnate with decurrent tooth in type or sinuate, white, with a faint grey cast in type, broad sub-distant, edges even; 1–2 lengths lamellulae inserted. Taste and odour faint, but disagreeable subaromatic in type, taste mild or slightly disagreeable, indistinct, similar to burnt plastic. Lamellae white (not fluorescing blue) under UVf365 nm. Basidiospores 8.1–10.2 × 5.6–6.1 µm in type description, pale yellow in Melzer’s reagent. Cuticle a thick (180–300 µm), gelatinous zone, the hyphae narrow colourless, more or less interwoven (an ixotrichodermium). Hypodermium a rather well-defined brownish zone. Pileus trama of radial hyphae. Clamp connections absent or very rare.
Habitat.
On soil under coastal redwoods (Sequoia sempervirens).
Geographical distribution.
Known only from coastal redwood forests in north-western North America (Fig. 5B).
Additional specimens examined.
USA • California: (1) Mendocino Co., Jackson State Forest, (39.3890,-123.68460, 26 Jan 2018, A. Rockefeller [voucher AR-2018a]), iNaturalist:9640562 (ITS:MG926555); (2) Humboldt Co., Eureka Area, Arcata Community Forest, near Sequoia sempervirens (40.88326, -124.06329, 16 Feb 2023, Mandy Hackney [voucher HAY-F-000661]), iNaturalist:148999043 (ITS:OR593569); (3) Humboldt Co., Eureka Area, McKay Community Forest, near Sequoia sempervirens (40.7501, -124.1446, 18 Nov 2023, Dean Lyons [voucher HAY-F-002303]), iNaturalist:191417805 (ITS:PP975575).
Notes.
This species, known only from the coastal regions of north California in North America, is found in coastal redwood (Sequoia sempervirens) forest. Its key distinguishing feature is its unpleasant odour, but the odour is not always present. The lamellae appear white under UVf365 nm, distinguishing it from the bright blue fluorescence of lamellae in G. calunus. Though first recorded in 1937, most records date from 2013 or later. Reports of this species from Vancouver B.C. in Canada are a misapplied name. M. Berbee & F. Stonehouse at UBC obtained an ITS sequence of a collection by A. and U. Češka from Vancouver (UBC-F-3506) that most closely matched the type description of H. subaromaticus. The GenBank BLAST of that sequence confirmed that it had been misidentified as H. subaromaticus and instead belonged to a possibly unknown species, near Cuphophyllus sp. ‘PNW01’ and C. subaustraligus. Although an ixocutis was found on the pileus and stipe of the Vancouver collections, the specimens differed in having only a somewhat viscid rather than glutinous pileus that is typical of both G. calunus and H. subaromaticus.
Diagnostic microcharacters in Gliophorus and section Unguinosae
Species of Gliophorus in section Unguinosae differ from section Glutinosae in lacking a gelatinised lamellar edge and ixocheilocystidia embedded in the gel. In addition, toruloid/open-loop medallion clamp connections on basidioles that are characteristic in section Glutinosae (Lodge et al. 2014) were apparently absent from all the species and collections of Gliophorus in section Unguinosae which we examined despite a concerted effort to search for these. Other species of Gliophorus in section Unguinosae, however, have toruloid clamp connections at the base of basidioles (Lodge et al. 2014). Microscopic characters were generally not helpful in distinguishing amongst Gliophorus species in sect. Unguinosae (Fig. 10), except that spores of G. parafumosus were noticeably shorter than in other species in this section including G. fumosus which has an overlapping range (mean 6.4 vs. 6.7 µm).
Macromorphology of Gliophorus subaromaticus basidiomes. A. G. subaromaticus USA, California: (1) Mendocino Co., Jackson State Forest”, (39.3890,-123.68460, 26 Jan 2018, A. Rockefeller [voucher AR-2018a]), iNaturalist:9640562 (ITS:MG926555); B. Humboldt Co., Eureka Area, Arcata Community Forest, near Sequoia sempervirens (40.88326, -124.06329, 16 Feb 2023, Mandy Hackney [voucher HAY-F-000661]), iNaturalist:148999043 (GenBank ITS sequence:OR593569); C. Humboldt Co., Eureka Area, McKay Community Forest, near Sequoia sempervirens (40.7501, -124.1446, 18 Nov 2023, Dean Lyons [voucher HAY-F-002303]), iNaturalist:191417805 (ITS:PP975575). UNITE/PlutoF:SH0910861.10FU (1.5% threshold).
Micro-drawings of spores and basidia of Gliophorus species in sect. Unguinosae. A, B. European species; C–E. North American species. A. G. alboviscidus, (DJH21-29 from UK); B. G. irrigatus (DJH 21-31, from UK); C. G. irrigatus (D. Boertmann, neotype, from Denmark); D. G. calunus, holotype (Mendocino Co. California, HAY:F-012170); E. G. fumosus, holotype (North Carolina, USA, TENN-F-061868 [DJL05NC50]); F. G. parafumosus, holotype (Bayfield, WI, F:C0348314F [NAMA 2017-248); G. G. subaromaticus, holotype, (Prairie Creek St. Park, Orick, CA, MICH:10963 [Smith 9167]). Scale bars: 10 µm.
Key to Gliophorus sect. Unguinosae
**: **
Discussion
The first new species reported here, G. alboviscidus, is identical in morphology to G. irrigatus, except for its colour. Initial conclusions that it was an albino form were dispelled by ITS sequence analysis, which found that both specimens formed a distinct clade from European G. irrigatus s.s. To our knowledge, Persoon (1801) did not provide any image of G. irrigatus, nor has any voucher survived, but it is likely that the specimen described in his monograph was found near Göttingen, Germany. Therefore, we neotypify G. irrigatus, based on voucher CFMR DEN-21 (KF291084), assessed by David Boertmann in Lodge et al. (2014), from Denmark, ca. 600 km north of Göttingen.
There was historically some debate as to whether G. irrigatus and G. unguinosus were distinct species (Arnolds 1990a; Boertmann 2010). The latter species was named by Fries, who also provided a painting based on a Swedish specimen. However, there is no evidence from either vouchers or eDNA data that more than one grey or brown species within Gliophorus sect. Unguinosae exists in Europe.
Our designated neotype of G. irrigatus (ITS:KF291084) and holotype of G. alboviscidus diverge by > 9% in their ITS sequences. It is noteworthy that G. irrigatus specimens from across Europe form a distinct clade hitherto not reported beyond Europe, even when eDNA data are assessed (single exception, see above), whereas G. alboviscidus has a broader distribution, being detected in eDNA datasets from China.
G. irrigatus-like basidiomes are widely reported from North America. However, our sequence analysis found these species to be quite distinct (ca. 85% identity for ITS locus) from their morphologically very similar European relatives. Here, we describe two sibling species found across eastern North America, G. fumosus and G. parafumosus, that form two genetically distinct clades, but having only subtle differences in the morphology of the pileus margin. eDNA sequences of G. fumosus extend from eastern to western USA, whereas G. parafumosa appears to be restricted to eastern North America.
A similar white to Buff-coloured species from the G. irrigatus complex is known from the western USA (G. subaromaticus), often characterised by an unpleasant odour and recently found to have white lamellae under UVf365 nm. Sequence data from the holotype confirmed that this to belongs to Gliophorus sect. Unguinosae. Additionally, from the western US, a sixth species was found with pale or often more strongly pigmented basidiomes and bright blue lamellae under UVf365 nm, G. calunus. It is also clear from the phylogenetic analysis of sequence data that additional species exist within Gliophorus sect. Unguinosae beyond the six species named here (e.g. OU003418 from Ecuador and MZ269231 from NY). In most cases, these are represented only by eDNA sequences, one exception being a single voucher (iNaturalist:30847408/MZ269231) from north-eastern USA, which appears to be morphologically distinct and is included in our key.
The other exception is a south-eastern US clade represented by a single voucher (iNaturalist:251773868/PV223767) provisionally named Gliophorus sp. ‘FL03’ from near Orlando, Florida that formed a clade with an environmental sequence from the Duke Forest in Durham, NC (GenBank:AY969860) and an ITS2 sequence from an ectomycorrhiza in Arkansas (GenBank:MF665104) (Fig. 2). This Gliophorus sp. ‘FL03’ appears inside a strongly supported clade with G. alboviscidus from Eurasia and G. calunus from north-western North America, which suggests it is part of a lineage that was previously widespread and diverged into three Grayan disjunct species.
Lastly, the grey-brown species found in Australia and named Hygrocybe irrigatus (Young 2005) (pp 114/128) is likely to be divergent and to represent a new species. GlobalFungi search using ITS1 sequence of G. fumosus did detect eDNA (95% ID) hits from soil samples collected in New South Wales and Tasmania (Bissett et al. 2016), suggesting that a distinct species with darker pileus and most closely related to G. fumosus is indigenous to Australasia (Suppl. material 2).
It is often accepted as a rule of thumb that species whose ITS sequences diverge by > 3% represent different distinct species (Blaalid et al. 2013; Kauserud 2023), though there are examples of members of the same species having greater levels of ITS divergence, up to 6% (Hughes et al. 2013). In our study, the most closely related (named) species pair, G. alboviscidus and G. calunus diverge by ca. 4%, but there is no evidence that their geographical ranges overlap even when eDNA data are assessed.
Assessing the distribution of novel species is by definition difficult. However, the advent and by now widespread use of eDNA metabarcoding provides a means to better assess species distributions. The GlobalFungi website provides an accessible means, whereby such datasets may be interrogated and, therefore, provides an invaluable resource for assessment of species distributions; one which will become progressively more useful as eDNA metabarcoding datasets are published at an accelerating rate. One point important to note is that eDNA data cannot assess whether the sequences detected derive from mature individuals, an important element in fungal conservation (Ainsworth et al. 2013; IUCN 2024), where occurrence of basidiomes forms the basis of biological recording and conservation policy.
In the case of Hymenomycetes, which are mostly outcrossing, establishment of a primary mycelial (monokaryon) from single basidiospore must be followed by mating with a compatible basidiospore or primary mycelium to permit formation of the secondary mycelium (dikaryon) and later fruiting (James 2023). However, follow-on field surveys for fruit bodies of the species detected can be geo-focused, based on eDNA data and this method has been successfully deployed in the UK (The Leasowes SSSI) to confer legal protection on sites of high grassland fungal diversity (UK_Government 2019; Detheridge and Griffith 2021).
Use of eDNA data to better understand the distributions of fungi has huge potential (Copoț et al. 2024), but it does not (and cannot) define mature (reproducing) individuals (the basis of biological recording, conservation policy and, for example, GBIF). For agaric fungi, most form highly mobile uninucleate spores which may land and infrequently form mycelia at great distance. However, these ‘lost monokaryons’ will only proceed to form mature individuals if they mate with a compatible basidiospore or mycelium, a much rarer event than the establishment of the original mycelium, if far beyond the normal range of the species. There are additional alternative explanations for eDNA being detected outside the range of agaric species known from spore-producing basidiomes.
Besides long-range dispersal via haploid basidiospores forming monokaryotic mycelia that fail to encounter an opposite mating type, as noted above, we hypothesise three additional reasons why the distributions of eDNA sequences may exceed the known ranges of Gliophorus species, based on basidome specimen records: 1) possible gaps or insufficient effort in collecting basidiomes or failing to collect basidiomes believed to be a common species (most likely explanation in our opinion); 2) spores may have dispersed beyond climatic zones that are currently or rarely conducive to fruiting; and 3) eDNA distributions exceeding the known range of basidiomes may indicate remnant populations of previously widespread distributions.
In the case of G. irrigatus s.s., basidiomes and eDNA have been intensively documented for Europe, including the UK and southern Scandinavia, regions where no other species are easily confused with it, basidiome and eDNA distributions are largely congruent (Fig. 3B, C). In contrast, the abundance of eDNA sequences of G. alboviscidus in eastern China, a species only known from basidiomes in the UK, might indicate insufficient surveys in eastern Eurasia, previous confusion of this new species with G. irrigatus from Europe or a formerly widespread distribution that has become disjunct with changes in climate. The relatively small divergence in ITS sequences between Eurasian G. alboviscidus and north-western North American G. calunus suggests disjunction of a formerly widespread range.
The new eastern North American species, G. fumosus, is abundantly documented by sequenced specimens and eDNA records in eastern North America as far west as Wisconsin, while eDNA records extend west to the northern (Montana and Colorado) and southern (New Mexico) Rocky Mountains, plus the Cascade Mountains in western Oregon. The combined eDNA and basidiome distribution of G. fumosus, omitting the Cascades, is largely concordant with the distribution of basidiomes of Chromosera lilacifolia (Peck) (Grootmyers et al. 2025).
Taken together, this suggests that the widespread distributions of these two predominantly eastern North American species of Hygrophoraceae may represent remnants of formerly more widespread distributions, but they differ in that C. lilacifolia is able to reproduce in the western part of its range, whereas G. fumosus has either failed to reproduce or more likely only reproduces rarely beyond the mid-western USA and adjacent Canada and/or has been ignored. Occurrences in the mountains of New Mexico are concordant with a southern east-west dispersal pathway via the transvolcanic belt in northern Mexico that has been documented for several species and sister species of boletes (Ortiz-Santana et al. 2007). Grayan disjunction of sister species and genera between North American and east Asian taxa occurred 3–15 (-25) MYA (Wang et al. 2004; Petersen and Hughes 2007; Yih 2012) and the uplift of the Rocky Mountains caused changes in rainfall to their east resulting in extirpations, followed by Quaternary glaciation cycles that extirpated additional populations in parts of North America and western Europe (Yih 2012). The estimated divergence times for the Hygrophoraceae and Gliophorus are early enough to be consistent with this hypothesis. The Stem Age for the Hygrophoraceae was estimated as 125 MYA and the divergence of the genus Gliophorus was dated to around 60–65 MYA (He et al. 2019).
Whilst there is evidence from eDNA that G. subaromaticus exists beyond North America, no fruit bodies have been observed beyond the far west of North America. However, the report of a subspecies of G. irrigatus (Gliophorus irrigatus f. ammoniacus) from Saarland, Germany, with “strong nitrous odour” (Schmitt 2022) suggests that G. subaromaticus may be more widely distributed. However, no sequence data nor any other details have been published for this specimen (IF/MB#842679), so potential synonymy cannot currently be resolved. eDNA studies at Aberystwyth University have detected G. subaromaticus in soil samples from upland (non-wooded) habitats in south Wales and the English Peak district (GWG unpub. data), but as noted above, these may well not represent mature individuals.
This study highlights how sustained efforts by citizen scientists — particularly through the Pembrokeshire Fungus Recording Network, with whom our first author is affiliated — can materially advance fungal systematics. Community-led observations and collections, integrated with molecular analyses, were essential to recognising the novel diversity reported here; continued support for such networks will accelerate future taxonomic discovery. The Fungal Diversity Survey (Ness 2021; Grootmyers et al. 2025) and citizen scientists collecting and sequencing specimens through The MycoMap Network, Mycota Lab, the Ohio Mushroom DNA Lab, the Counter Culture Lab in Oakland CA and sequencing of select foray specimens by the North American Mycological Association have been critically important in documenting new species, describing variation within species and extending ranges of known species in North America.
In this study, we have been careful to obtain where possible holotype samples for ITS sequencing and, where this was not possible, to neotypify species according to best practice guidelines. Failure to follow such an approach when naming new species can lead to future confusion and we advocate that fellow fungal taxonomists focus not only on naming new taxa, but also clarifying the concepts of well-established species.
Supplementary Material
XML Treatment for Gliophorus alboviscidus
XML Treatment for Gliophorus irrigatus
XML Treatment for Gliophorus calunus
XML Treatment for Gliophorus fumosus
XML Treatment for Gliophorus parafumosus
XML Treatment for Gliophorus subaromaticus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abarenkov K, Nilsson RH, Larsson K-H, Taylor AF, May TW, Frøslev TG, Pawlowska J, Lindahl B, Põldmaa K, Truong C (2024) The UNITE database for molecular identification and taxonomic communication of fungi and other eukaryotes: Sequences, taxa and classifications reconsidered. Nucleic Acids Research 52: D 791–D 797. 10.1093/nar/gkad 1039 PMC 1076797437953409 · doi ↗ · pubmed ↗
- 2Ainsworth A, Smith J, Boddy L, Dentinger B, Jordan M, Parfitt D, Rogers H, Skeates S (2013) Red List of Fungi for Great Britain: Boletaceae; A pilot conservation assessment based on national database records, fruit body morphology and DNA barcoding Species Status 14. Joint Nature Conservation Committee, Peterborough, 37 pp.
- 3Bahram M, Netherway T, Hildebrand F, Pritsch K, Drenkhan R, Loit K, Anslan S, Bork P, Tedersoo L (2020) Plant nutrient‐acquisition strategies drive topsoil microbiome structure and function. The New Phytologist 227: 1189–1199. 10.1111/nph.1659832279325 · doi ↗ · pubmed ↗
- 4Baldrian P, Větrovský T, Lepinay C, Kohout P (2021) High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Diversity: 1–9. 10.1007/s 13225-021-00472-y · doi ↗
- 5Bissett A, Fitzgerald A, Meintjes T, Mele PM, Reith F, Dennis PG, Breed MF, Brown B, Brown MV, Brugger J (2016) Introducing BASE: the Biomes of Australian Soil Environments soil microbial diversity database. Gigascience 5: s 13742-13016-10126-13745. 10.1186/s 13742-016-0126-5PMC 487075227195106 · doi ↗ · pubmed ↗
- 6Blaalid R, Kumar S, Nilsson RH, Abarenkov K, Kirk P, Kauserud H (2013) ITS 1 versus ITS 2 as DNA metabarcodes for fungi. Molecular Ecology Resources 13: 218–224. 10.1111/1755-0998.1206523350562 · doi ↗ · pubmed ↗
- 7Boertmann D (1995) The genus Hygrocybe. Fungi of Northern Europe (Vol. 1). Danish Mycological Society (1st edn.), 287 pp.
- 8Boertmann D (2010) The genus Hygrocybe. Danish Mycological Society (2nd edn.), 287 pp.