Multigene phylogenetic analyses and morpho-anatomical characterization revealed two new species of Xylariomycetidae from Yunnan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
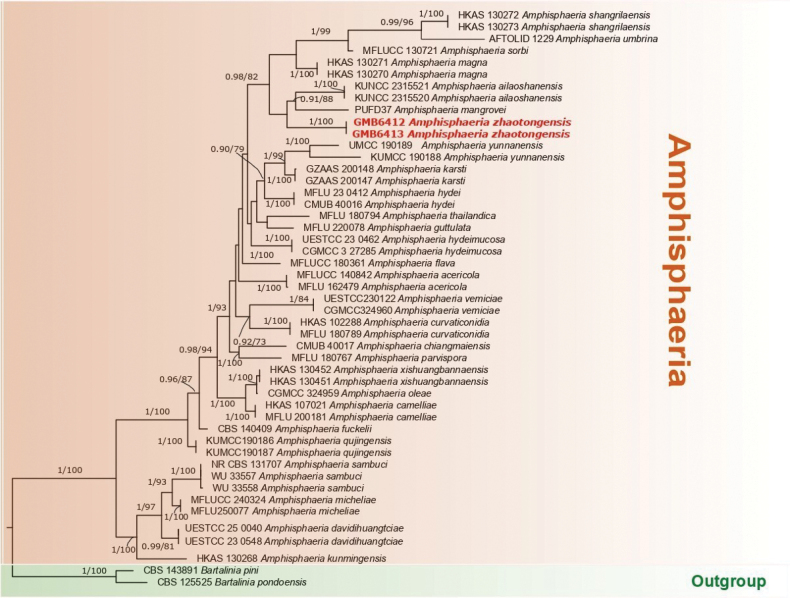 Figure 1
Figure 1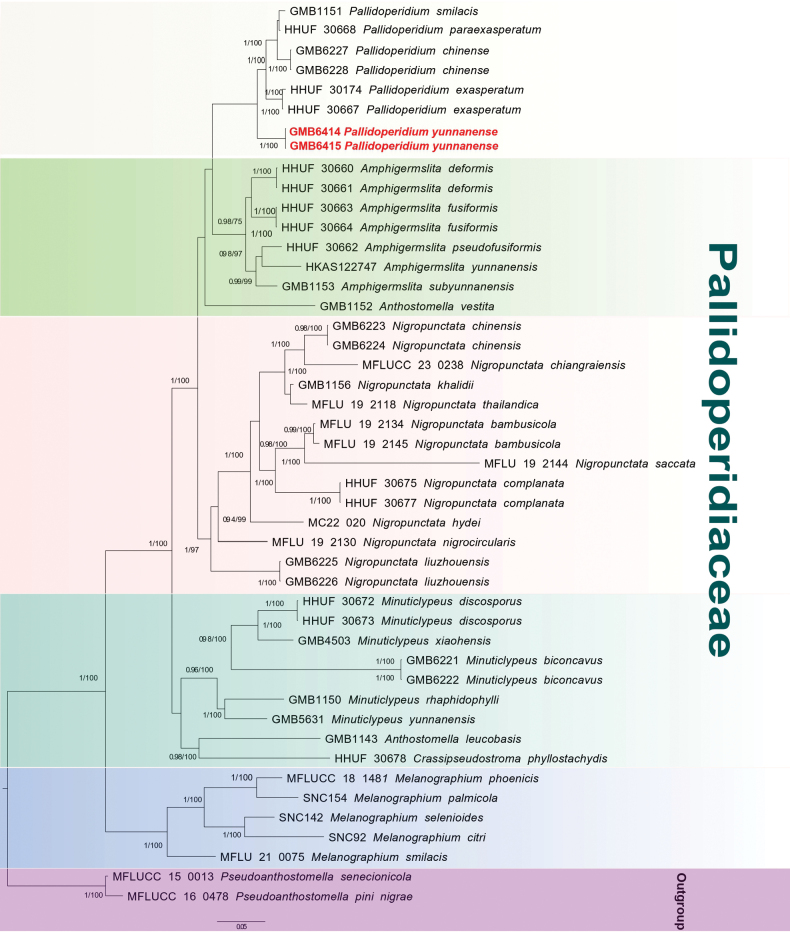 Figure 2
Figure 2 Figure 3
Figure 3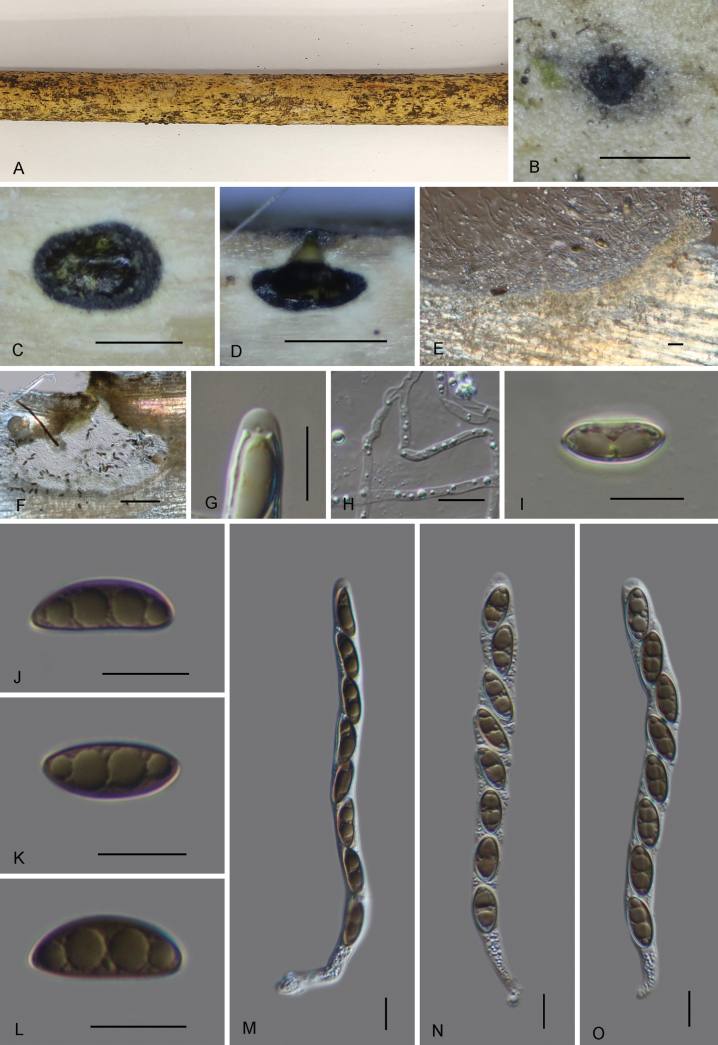 Figure 4
Figure 4| Species | Voucher | GenBank Accession Numbers | References | |||
|---|---|---|---|---|---|---|
|
|
|
|
| |||
|
| HHUF 30660T |
|
|
|
|
|
|
| HHUF 30661 |
|
|
|
|
|
|
| HHUF 30663T |
|
|
|
|
|
|
| HHUF 30664 | – |
|
|
|
|
|
| HHUF 30662T |
|
|
|
|
|
|
| GMB1153 |
|
| – | – |
|
|
| HKAS 122747 |
|
| – | – |
|
|
| MFLUCC 14-0842 |
|
| – | – |
|
|
| MFLU 16-2479 |
|
| – | – |
|
|
| KUNCC 23-15520T |
|
| – | – |
|
|
| KUNCC 23-15521 |
|
| – | – |
|
|
| HKAS 107021T |
|
|
|
|
|
|
| MFLU 20-0181 |
|
|
| – |
|
|
| CMUB 40017 |
|
|
|
|
|
|
| MFLU 18-0789T |
|
| – |
|
|
|
| HKAS 102288 |
|
|
| – |
|
|
| UESTCC 23-0548 |
|
|
| – |
|
|
| UESTCC 25-0040T |
|
|
| – |
|
|
| MFLUCC 18-0361T |
|
| – |
|
|
|
| CBS 140409T |
| – |
|
|
|
|
| MFLU 22-0078T |
|
| – | – | – |
|
| CMUB 40016 |
|
|
|
| Samarakoon (2023) |
|
| MFLU 23-0412T |
|
|
| – | Samarakoon (2023) |
|
| CGMCC 3.27285 |
|
| - |
| |
|
| UESTCC:23.0462 |
|
|
| - |
|
|
| GZAAS 20-0148T |
|
| – | – |
|
|
| HKAS 130268 |
|
| – | – |
|
|
| HKAS 130270T |
|
| – | – |
|
|
| HKAS 130271 |
|
| – | – |
|
|
| PUFD37T |
|
| – | – |
|
|
| HKAS 128843 |
|
|
| – |
|
|
| MFLU 18-0767 |
|
|
|
|
|
|
| KUMCC 19-0186 |
|
| – | – |
|
|
| KUMCC 19-0187T |
|
|
| – |
|
|
| CBS 131707T |
|
| – |
| |
|
| WU 33557T |
| – | – | – |
|
|
| WU 33558 |
| – | – | – |
|
|
| HKAS 130272T |
|
| – | – |
|
|
| HKAS 130273 |
|
| – | – |
|
|
| MFLUCC 13-0721T |
|
| – | – |
|
|
| MFLU 18-0794T |
|
|
| – |
|
|
| HKUCC 994 |
|
| – | – | Jeewon et al. (2016); |
|
| UESTCC 23.0122 |
|
|
| – | Samarakoon (2023) |
|
| CGMCC 3.24960T |
|
|
| – | Samarakoon (2023) |
|
| HKAS 130451T |
|
| – | – |
|
|
| HKAS 130452 |
|
| – | – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| GMB1143T |
| – |
| – |
|
|
| GMB1152T |
| – | – | – |
|
|
| CBS 125525 |
|
|
|
|
|
|
| CBS 143891 |
|
|
|
|
|
|
| HHUF 30678T |
|
|
|
|
|
|
| SNC92T |
|
|
|
|
|
|
| SNC154T |
|
|
|
|
|
|
| MFLUCC 18-1481T |
|
| – |
|
|
|
| SNC142T |
|
| – |
|
|
|
| GMB6221T |
|
| – |
|
|
|
| GMB6222 |
|
| – |
|
|
|
| HHUF 30672T |
|
|
|
|
|
|
| HHUF 30673 |
|
| – |
|
|
|
| GMB1150 |
|
| – | – |
|
|
| GMB4503 |
|
| – |
|
|
|
| GMB5631T |
|
| – | – |
|
|
| MFLU 19-2134T |
|
|
|
|
|
|
| MFLU 19-2145T |
|
|
|
|
|
|
| GMB6223T |
| – |
|
|
|
|
| GMB6224 |
|
|
|
|
|
|
| MFLUCC 23-0238T |
| – |
|
|
|
|
| HHUF 30675T |
|
|
|
|
|
|
| HHUF 30677 |
|
|
|
|
|
|
| MC22-020T |
|
|
| – |
|
|
| GMB1156T |
| – | – | – |
|
|
| GMB6225T |
|
| – |
|
|
|
| GMB6226 |
|
| – |
|
|
|
| MFLU 19-2130T |
|
| – |
|
|
|
| MFLU 19-2144T |
|
|
| – |
|
|
| MFLU 19-2118T |
|
|
|
|
|
|
| GMB6227T |
|
| – |
|
|
|
| GMB6228 |
|
| – |
|
|
|
| HHUF 30174T |
|
|
|
|
|
|
| HHUF 30667 |
|
| – |
|
|
|
| HHUF 30668T |
|
| – |
|
|
|
| GMB1151 |
|
| – | – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| MFLUCC 16-0478 |
|
| – | – |
|
|
| MFLUCC 15-0013T |
|
|
|
|
|
| Character |
|
|
|
|
|
|---|---|---|---|---|---|
| Ascomata size | 293–433 µm height × 446–700 µm diam. | 300–440 µm height × 320–390 µm diam. | 290–350 µm height × 320–340 µm diam. | 300–500 µm height × 350–600 µm diam. | 412–443 µm height × 292–443 µm diam. |
| Ascospore size | 14.4–18 × 5.5–7.3 | 15.5–22.5 × 6–7.5 | 15–19 × 5.5–7.5 | 18–35 × 5–9.5 | 16.3–18.3 × 7.7–8.4 |
| Ascospore ornamentation | Rough, narrow mucilaginous sheath | Rough, with thick mucilaginous sheath | Rough, with mucilaginous sheath | Smooth to rough, thick mucilaginous sheath | Smooth, with mucilaginous sheath |
| Apical apparatus | Inamyloid | Amyloid, thin discoid | Amyloid, thin discoid | Inamyloid | Inamyloid |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology
Introduction
Yunnan Province, located in southwestern China, is a globally recognized biodiversity hotspot. Its complex topography and range of habitats, including extensive karst systems and montane forests, support remarkably high levels of species richness and endemism (Xu et al. 2017). Mycological research in Yunnan has accelerated markedly in the last decade, with reports focusing on areas such as fungal secondary metabolites, ethno-mycology, and the taxonomy of microfungi and macrofungi using morpho-molecular approaches (Hongsanan et al. 2023). Systematic mycological surveys are essential, not only to uncover misidentified or poorly known species but also to reveal undescribed diversity. Such deeper exploration of fungal richness can also provide valuable insights into ecological interconnections and overall ecosystem health (Wijayawardene et al. 2021; Dissanayake et al. 2024; Habib et al. 2025; Liu et al. 2025).
In this publication, we describe two new species belonging to the subclass Xylariomycetidae, representing the families Amphisphaeriaceae and Pallidoperidiaceae. Amphisphaeriaceae was introduced by Winter (1887, as “Amphisphaerieae”) to accommodate the type genus Amphisphaeria Ces. & De Not. and related taxa. Members of Amphisphaeriaceae are predominantly saprobes on decaying wood in terrestrial, marine, and freshwater habitats, although some taxa are hemibiotrophic or necrotrophic (Wang et al. 2004; Senanayake et al. 2015, 2019Jaklitsch et al. 2016). According to the latest Outline of Fungi (Hyde et al. 2024), Amphisphaeriaceae presently contains the two genera Amphisphaeria and Labridella Brenckle. Amphisphaeria is the type genus of the family, with A. umbrina (Fr.) De Not. designated as the type species (Cesati and De Notaris 1863). Species of Amphisphaeria are known to produce both coelomycetous and hyphomycetous asexual morphs and have saprobic and endophytic lifestyles, commonly on woody substrates and some monocot hosts including grasses. (Samarakoon et al. 2020, 2022; Wang et al. 2023; Dissanayake et al. 2024). The teleomorph of the genus is characterized by solitary or aggregated ascomata under a poorly-developed clypeus or lack of clypeus; unitunicate asci with an apical apparatus that is amyloid or inamyloid, and light brown to dark brown, ellipsoid to fusiform, 1–3-septate ascospores (Cesati and De Notaris 1863; Pathirana et al. 2025; Samarakoon et al. 2020; Samarakoon et al. 2023). According to Species Fungorum (accessed 28 September 2025), the genus currently comprises 169 species. Pallidoperidiaceae was established by Sugita et al. (2024) to accommodate a distinct, monophyletic lineage of Anthostomella-like fungi associated with bamboo. Sugita et al. (2024) included five genera (Amphigermslita R. Sugita & Kaz. Tanaka, Crassipseudostroma R. Sugita & Kaz. Tanaka, Minuticlypeus R. Sugita & Kaz. Tanaka, Pallidoperidium R. Sugita & Kaz. Tanaka, and Nigropunctata M.C. Samar. & K.D. Hyde) in the family. Recently, Lu et al. (2025) included Melanographium, previously considered incertae sedis, within Pallidoperidiaceae based on multigene phylogenetic analyses and concordance in teleomorph characters. Pallidoperidiaceae is typified by the genus Pallidoperidium, of which P. exasperatum R. Sugita & Kaz. Tanaka is the type species. The genus is characterized by perithecial ascomata, immersed, solitary, with a conical to cylindrical, periphysate ostiolar neck, a thick, inconspicuous ascomatal wall composed of hyaline to pale brown cells, surrounded by pseudostromatic tissue, numerous, septate, unbranched paraphyses, unitunicate, cylindrical, 8-spored asci with a thin discoid, amyloid apical apparatus, and ellipsoid to fusiform, unicellular, brown, rough ascospores surrounded by a mucilaginous sheath and with a germ slit (Sugita et al. 2024). Pallidoperidium comprises four species, with P. chinense K. Habib et al. and P. smilacis (Fabre) K. Habib & Q.R. Li reported from China (Habib et al. 2025), and P. exasperatum R. Sugita & Kaz. Tanaka and P. paraexasperatum R. Sugita & Kaz. Tanaka from Japan (Sugita et al. 2024).
In a study focused on the diversity of Xylariomycetidae in Guizhou, China, we identified two species belonging to the genera Amphisphaeria and Pallidoperidium. Morphological comparisons, together with phylogenetic analyses based on ITS, LSU, rpb2, and tef1 loci, showed that these taxa did not correspond to any previously described species within their respective genera. Based on this, we propose two new species, for which we provide brief diagnoses, detailed descriptions, illustrations, and phylogenetic placements.
Materials and methods
Sample collection and morphological study
The specimens were collected during surveys conducted in Yunnan province between 2023 and 2024, with all relevant habitat information recorded in a detailed manner. The photos of the collected materials were taken using a Canon G15 camera (Canon Corporation, Tokyo, Japan). Materials were placed in paper bags and taken to the lab for morphological characterization and isolation. To remove excess humidity, they were dried in the shade at room temperature. The dried specimens were carefully labeled and stored until further processing. All specimens were deposited at the Herbarium of Guizhou Medical University (GMB) and the Herbarium of Cryptogams, Kunming Institute of Botany, and the Chinese Academy of Sciences (KUN), while living cultures were deposited at the Guizhou Medical University Culture Collection (GMBC).
Morphological characterization and isolation
Macroscopic features of the specimens were examined using an Olympus SZ61 stereomicroscope and photographed using a Canon 700D digital camera. Microscopic morphological features (ascomata, peridium, paraphyses, asci, ascospore), were observed using DIC and photographed using a Canon 700D digital camera attached. Melzer’s iodine reagent was used to test the apical apparatus structures for amyloid reaction. Asci and ascospores of the samples were measured using Tarosoft Image Framework (v. 0.9.0.7). Images were processed using Adobe Photoshop CS6 (Adobe Systems, USA). Pure cultures were obtained by single-ascospore isolation (Long et al. 2019) and maintained at 25 °C for 1–5 weeks on PDA (potato dextrose agar) and oatmeal-agar (OA) medium by adding Veterinary sterptpmycin, on 100,000 units per liter.
DNA extraction, PCR amplification and sequencing
Mycelium was scraped from pure culture plates using a sterilized scalpel and used for DNA extraction using the BIOMIGA fungus genomic DNA extraction kit following the manufacturer’s instructions. For some specimens where the ascospores did not germinate, we directly extracted DNA from the contents of the perithecium. The DNA samples were kept at –20 °C. Sequences of the internal transcribed spacer (ITS), the large subunit (LSU), rpb2 and translation elongation factor (tef1α) were amplified by PCR with primers ITS1/ITS4 (White et al. 1990; Gardes and Bruns 1993), LR0R/LR5 (Vilgalys and Hester 1990), Bt2a/Bt2b (Glass and Donaldson 1995), and EF1-983F/EF1-2218R (Rehner and Buckley 2005), respectively. The components of a 25 μL volume PCR mixture were: 9.5 μL of double distilled water, 12.5 μL of PCR Master Mix, 1 μL of each primer, and 1 μL of template DNA. Qualified PCR products were checked through 1.5% agarose gel electrophoresis stained with Golden View, and were sent to Sangon Co., China, for sequencing.
Sequence alignments and phylogenetic analyses
All the obtained sequences were deposited in GenBank. Generated sequences were compared with each other and deposited sequences available through GenBank using the BLASTn. A molecular phylogeny was inferred from a combined dataset of ITS, LSU, rpb2 and tef1 sequences. The reference sequences retrieved from open databases originated from recently published literature (Habib et al. 2025; Dissanayake et al. 2024; Du et al. 2025; Li et al. 2024 and Sugita et al. 2024) and the BLASTn results of close matches (Table 1).
Sequences were aligned using the MAFFT v.710 online program (Katoh et al. 2019) with default settings. The alignment was adjusted manually using BioEdit v.7.0.5.3 (Hall 1999) where necessary. The phylogeny was inferred using maximum likelihood (ML) analysis as implemented in RAxML v.8.22 using the GTRGAMMA substitution model with 1,000 bootstrap replicates (Stamatakis 2014). A second phylogenetic analysis was performed following Bayesian methodology in MrBayes v. 3.2 (Ronquist et al. 2012) online. A Markov Chain Monte Carlo (MCMC) sampling in MrBayes v.3.2.2 (Ronquist et al. 2012) was used to determine the posterior probabilities (PP). Six simultaneous Markov chains were run for 1,000,000 generations, and trees were sampled every 1,000^th^ generation. The phylogenetic tree was visualized in FIGTREE v.4.4 (Rambaut 2018). All analyses were run on the CIPRES Science Gateway v 3.3 webportal (Miller et al. 2010).
Results
Phylogenetic Analysis of Amphisphaeria
The combined sequence matrix of Amphisphaeria comprised 3061 (ITS: 1–501, LSU: 501–1386, rpb2: 1386–2206 and tef1: 2206–3061) characters, after exclusion of ambiguously aligned regions and long gaps. The concatenated alignment of ITS, LSU, tef1-α and rpb2 comprised 3061 characters, of which 979 were parsimony-informative. Bartalinia pondoensis Marinc., Gryzenh. & M.J. Wingf (CBS 125525) and B. pini F. Liu, L. Cai & Crous (CBS 143891) were chosen as the outgroup taxa (Marincowitz et al. 2010; Liu et al. 2019). A BLASTn query of the newly generated ITS sequence (GMB6412) showed 90.4% nucleotide similarity with Amphisphaeria ailaoshanensis (PP584673).
The sequences of our collection Amphisphaeria zhaotongensis formed a clade, sister to A. ailaoshanensis and A. mangrovei. Although this relationship received low statistical support, it consistently appeared in both ML and Bayesian analyses. (Fig. 1).
Phylogram of the best ML tree (-lnL = 18843.27) revealed by RAxML from an analysis of the combined ITS, LSU, rpb2 and tef-1 sequence matrix of Amphisphaeria. Maximum Likelihood (ML) bootstrap support values higher than 70% and Bayesian posterior probabilities (BPP) higher than 0.90 are displayed above or below the respective branches at first and second position, respectively. The newly described species is formatted in red.
Phylogenetic Analysis of Pallidoperidium
The ITS sequences extracted from our collection showed 90.4% similarity with Pallidoperidium smilacis (PP153387) when subjected to BLASTn query in GenBank. The combined sequence matrix of Pallidoperidiaceae (Fig. 2) comprised 3106 (ITS: 1-514 bp/198 PI, LSU: 514-1370 bp/138 PI, rpb2: 1370-2239 bp/240 PI and tef1: 2239-3003 bp/217 PI), after exclusion of ambiguously aligned regions and long gaps. Pseudoanthostomella senecionicola Daranag., Camporesi & K.D. Hyde (MFLUCC 150013) and P. pini-nigrae Daranag., Camporesi & K.D. Hyde (MFLUCC 160478) were chosen as the outgroup taxa (Daranagama et al. (2016).
Phylogram of the best ML tree (-lnL = –20567.83) revealed by RAxML from an analysis of the combined ITS, LSU, rpb2 and tef-1 sequence matrix of Pallidoperidiaceae. Maximum Likelihood (ML) bootstrap support values higher than 70% and Bayesian posterior probabilities (BPP) higher than 0.90 are displayed above or below the respective branches at first and second position, respectively. The newly described species is formatted in red.
In the phylogram (Fig. 2), the sequences of our Pallidoperidium collections (GMB6414 and GMB6415) formed a well-supported, distinct clade in a basal position within Pallidoperidium, supporting the recognition of a new species, described below as Pallidoperidium yunnanense.
Taxonomy
Amphisphaeria
zhaotongensis
Taxon classificationFungiAmphisphaerialesAmphisphaeriaceae
XY Luo & Q. R. Li sp. nov.
5541E9C7-CC68-5DEE-8254-D6A322C1790F
861335
Etymology.
The specific epithet “zhaotongensis” refers to Zhaotong city, where the holotype specimen was collected.
Type.
China • Yunnan Province, Zhaotong city, Yongshan county, Wumengshan National Nature Reserve, 28°19'29.81"N, 104°00'05.54"E, altitude: 1367 m, on dead branch of an unidentified plant, June 2024, collected by Xingyu Luo, WMS162 (GMB6412, holotype; KUN-HKAS 151613, isotype; GMBC6412 ex-holotype culture).
Amphisphaeria zhaotongensis (GMB6412, holotype). A. Dead corticated host branch with immersed ascomata; B–D. Surface view of ascomata; E. Cross section of ascoma; F, G. Vertical section of ascomata; H. Peridium; I, J. Amyloid ascal apical apparatus (stained in Melzer’s reagent); K. Paraphyses; L–O. Ascospores; P–R. Asci. Scale bars: 0.5 mm (D–F); 100 μm (G); 10 μm (H–R)
Paratype. China • Yunnan Province, Zhaotong city, Yongshan county, Wumengshan National Nature Reserve, 28°19'29.70"N, 104°00'11.04"E, altitude: 1322 m, on dead branch of an unidentified plant, June 2024, collected by Xingyu Luo, WMS613 (GMB6413, paratype; GMBC6413, ex-paratype culture).
Description.
Saprobic on the surface of a dead unknown plant. Teleomorph: Ascomata 350–610 µm high × 138–315 µm diam. (x̄ = 511 × 195 µm, n = 5), perithecial, immersed in sometimes slightly uneven bark, only visible as minute dark dots, scattered or aggregated in small groups, depressed globose, dark. Peridium 32–40 µm thick, composed of several layers of pseudoparenchymatous cells, cells small to medium-sized, thin- to slightly thick-walled, brown externally and hyaline internally. Paraphyses 2.2–4.7 µm wide, hyaline, simple, multiguttulate, septate. Asci 118.5–166 × 6.9–9.3 µm (x̄ = 138 × 7.8 µm, n = 30), 8-spored, unitunicate, cylindrical, with a short stipe, with apical apparatus 1.1–1.7 × 0.3–0.5 µm (x̄ = 1.4 × 0.4 µm, n = 10), amyloid, bluing in Melzer’s reagent. Ascospores 17.7–21 × 4.6–6.6 µm (x̄ = 19 × 5.9 µm, n = 30), L/W = 3.2, uniseriate, oblong or narrowly fusiform, hyaline, 1-septate at the center, slightly constricted at the septum, smooth, straight to slightly curved, thick-walled, without a mucilaginous sheath. Anamorph: Not observed.
Notes.
Phylogenetically, Amphisphaeria zhaotongensis is closely related to A. ailaoshanensis L.S. Dissan., K.D. Hyde & J.C. Kang and A. mangrovei Devadatha & V.V. Sarma. In BLASTn search, the ITS sequence of A. zhaotongensis showed highest similarity to A. ailaoshanensis (KUNCC 2315520, KUNCC 2315521) with 95.59%, followed by A. mangrovei (PUFD37) with 90.84%. The LSU sequences of A. zhaotongensis showed 97.51% similarity to A. ailaoshanensis (KUNCC 2315520, KUNCC 2315521) and 98.39% to A. mangrovei (PUFD37). Morphologically, A. zhaotongensis resembles A. ailaoshanensis in ascospore size but differs by having significantly larger ascomata (350–610 × 138–315 µm vs. 100–140 × 250–350 µm), longer asci (x̄ = 138 × 7.8 µm vs. 87 × 8 µm), an amyloid apical apparatus (vs. inamyloid), and ascospores with narrow ends (vs. rounded) (Dissanayake et al. 2024).
Amphisphaeria mangrovei can be easily distinguished from A. zhaotongensis by its smaller ascomata (150–280 µm high × 140–250 µm diam vs. 350–610 µm high × 138–315 µm diam. and smaller ascospores (12–15 µm in length vs. 17.7–21 µm in length) (Phookamsak et al. 2019).
Amphisphaeria zhaotongensis is morphologically similar to the type species Amphisphaeria umbrina in having immersed, globose ascomata, cylindrical asci an amyloid apical apparatus, and 1-septate ascospores. However, it differs from A. umbrina in having hyaline and slightly smaller ascospores (17.7–21 × 4.6–6.6 µm), whereas A. umbrina has brown and slightly larger ascospores (20–25 × 6–8 µm).
Pallidoperidium
yunnanense
Taxon classificationFungiXylarialesPallidoperidiaceae
Q. R. Li sp. nov.
E6184B00-4533-5F20-A3E0-045C54004D1C
861336
Etymology.
The specific epithet “yunnanense” refers to the Yunnan Province, where the holotype specimen was collected.
Pallidoperidium yunnanense (GMB6414, holotype). A. Dead corticated host culm with immersed ascomata; B. Surface view of ascomata; C. Cross section of ascoma; D, F. Vertical section of ascoma; E. Peridium; G. Inamyloid ascal apical apparatus (stained in Melzer’s reagent); H. Paraphyses; I–L. Ascospores; M–O. Asci. Scale bars: 0.5 mm (B–D); 100 μm (F); 10 μm (E, G–O)
Type.
China • Yunnan Province, Zhaotong city, Yongshan county, Wumengshan National Nature Reserve, 28°19'29.72"N, 104°00'11.55"E, altitude: 1228 m, on dead bamboo, June 2024, collected by Xingyu Luo, WMS149 (GMB6414, holotype; KUN-HKAS 151614, isotype; GMBC6414 ex-holotype culture).
Paratype. • Yunnan Province, Zhaotong city, Yongshan county, Wumengshan National Nature Reserve, 28°19'29.36"N, 104°00'10.28"E, altitude: 1300 m, on dead bamboo, June 2024, collected by Xingyu Luo, WMS614 (GMB6415, paratype; GMBC6415, ex paratype culture).
Description.
Saprobic on dead bamboo. Teleomorph: Ascomata 293–433 µm high, 446–700 µm diam., perithecial, deeply immersed in host tissue, solitary, subglobose. Ostiolar neck conical to cylindrical, 175–233 µm diam., periphysate. Ascomatal wall 15-56 µm thick, composed of 3-5 layers of elongate cells of 5–8.8 × 2–3 µm, hyaline at the inside, pale brown towards the outside. Paraphyses 1.7–3 µm wide, numerous, septate, unbranched, cylindrical, hyaline. Asci 118–151 × 6.7–10.3 µm (x̄ = 130 × 8.5 µm, n = 20), 8-spored, unitunicate, cylindrical, with a short stipe, with apical apparatus not bluing in Melzer’s reagent. Ascospores 14.4–18 × 5.5–7.3 µm (x̄ = 16 × 6.4 µm, n = 30), l/w = 2.5, uniseriate, ellipsoid to fusiform, unicellular, brown, surrounded by a narrow mucilaginous sheath. Anamorph: Not observed.
Habit and habitat.
Scattered on dead bamboo.
Notes.
Morphologically, Pallidoperidium yunnanense resembles P. exasperatum R. Sugita & Kaz. Tanaka and P. paraexasperatum R. Sugita & Kaz. Tanaka, in having immersed perithecial ascomata, cylindrical unitunicate asci with a discoid apical apparatus, and brown, ellipsoid to fusiform ascospores. However, P. yunnanense differs from P. exasperatum by having markedly broader ascomata (446–700 µm vs. 320–390 µm wide) and ascospores with a narrow mucilaginous sheath (vs. ascospores with a thick mucilaginous sheath) (Sugita et al. 2024).
P. yunnanense is distinguished from the additional two species in the genus by its smaller, narrower ascospores (14.4–18 × 5.5–7.3 µm) compared to P. chinense (18–35 × 5–9.5 µm) and P. smilacis (16.3–18.3 × 7.7–8.4 µm). In addition, P. chinense and P. smilacis have smooth to roughened ascospores with a thick mucilaginous sheath, while mucilaginous sheath narrow in P. yunnanense (Habib et al. 2025).
Phylogenetically, Pallidoperidium yunnanense forms an independent clade in a basal position within the genus. Although phylogenetically distinct, its morphology is consistent with the generic concept of Pallidoperidium, including subglobose, deeply immersed perithecial ascomata, discoid apical apparatus, and ellipsoid to fusiform brown ascospores (Table 2).
Discussion
Southwestern China is considered a hotspot for diverse fungal taxa, especially for wood-inhabiting fungi which contribute considerably to nutrient cycling and ecosystem balance (Hyde et al. 2020; Samarakoon et al. 2022). Here, we describe two novel taxa from this unique habitat, namely, Amphisphaeria zhaotongensis and Pallidoperidium yunnanense, characterized and described by using both a morphological and multi-gene phylogenetic approach. These discoveries underscore the still hidden fungal diversity of this region and provide insights into phylogenetic relationships within Xylariomycetidae.
Species of Amphisphaeria are usually saprobes on woody branches of different hosts, distributed in diverse ecological zones, such as grasses in terrestrial, mangrove, and freshwater habitats (Senanayake et al. 2019; Samarakoon et al. 2020; Samarakoon et al. 2022; Dissanayake et al. 2020). However, A. orixae was isolated as an endophyte from the medicinal plant O. japonica (Wang et al. 2023).
Species of Amphisphaeria are morphologically distinguished by immersed to erumpent ascomata, unitunicate asci with an apical apparatus that is amyloid or inamyloid, and often hyaline to brown ascospores (Samarakoon et al. 2020; Wang et al. 2023; Dissanayake et al. 2024); however, these characteristics frequently overlap across species. Due to significant morphological overlap among species and by its historical ambiguity with the genus Lepteutypa, which was previously distinguished primarily on stromatal form and ascospore septation but which cannot be phylogenetically separated from Amphisphaeria, the genus Amphisphaeria has undergone numerous taxonomic rearrangements (Samarakoon et al. 2020; Senanayake et al. 2015).
Our phylogenetic analysis confirms the placement of A. zhaotongensis within Amphisphaeria, forming a distinct lineage among recently described Chinese species. The discovery of this species in subtropical forests contributes to our understanding of the genus’s diversity in the region and highlights the value of multilocus approaches for reliable species identification. To date, Species Fungorum recognizes approximately 169 species of the genus (accessed September 18, 2025). Prior to this study, Amphisphaeria species reported from China include A. ailaoshanensis, A. xishuangbannaensis, A. yunnanensis, A. karsti, A. shangrilaensis, A. qujingensis, A. kunmingensis, A. davidihuangtciae, A. hydeimucosa (Dissanayake et al. 2020, 2024; Du et al. 2025; Habib et al. 2025). With the addition of A. zhaotongensis, the known diversity of the genus in China is further expanded.
The family Pallidoperidiaceae was recently introduced to accommodate and resolve a longstanding ambiguity for the placement of several Anthostomella-like taxa (Sugita et al. 2024). Prior to this study, four species were recognized worldwide: P. exasperatum, P. paraexasperatum from Thailand and P. chinense, P. smilacis from China (Sugita et al. 2024; Habib et al. 2025).
All known species of Pallidoperidium, including the newly described P. yunnanense, have been collected exclusively from bamboo substrates indicating a strong host preference of this genus (Sugita et al. (2024); Habib et al. (2025). Such host specificity may indicate an ecological adaptation to the lignocellulosic composition or microenvironmental conditions of bamboo culms. Moreover, additional sampling from additional bamboo hosts and a wider geographical area is required to confirm whether Pallidoperidium is an exclusively bamboo-associated lineage or simply a group with strong ecological affinity for bamboo habitats.
The addition of Pallidoperidium yunnanense and Amphisphaeria zhaotongensis highlights the taxonomic and ecological diversity of Yunnan’s Xylariomycetidae. Resolving taxonomic boundaries within Amphisphaeriaceae and Pallidoperidiaceae will require further comprehensive studies that combine morphology with multigene phylogenetic frameworks, as many species lack molecular data and several allied species are still poorly defined.
Supplementary Material
XML Treatment for Amphisphaeria zhaotongensis
XML Treatment for Pallidoperidium yunnanense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Daranagama DA, Camporesi E, Jeewon R, Liu X, Stadler M, Lumyong S, Hyde KD (2016) Taxonomic rearrangement of Anthostomella (Xylariaceae) based on a multigene phylogeny and morphology. Cryptogamie. Mycologie 37: 509–538. 10.7872/crym/v 37.iss 4.2016.509 · doi ↗
- 2Dissanayake LS, Samarakoon MC, Mortimer PE, Lu YZ, Li QR, Hyde KD, Kang JC (2020) Morpho-molecular characterization of two novel amphisphaeriaceous species from Yunnan, China. Phytotaxa 446: 144–158. 10.11646/phytotaxa.446.2.2 · doi ↗
- 3Dissanayake LS, Samarakoon MC, Maharachchikumbura SSN, Hyde KD, Tang X, Li QR, Mortimer PE, Faraj TK, Xu JC, Kang JC, Wanasinghe DN (2024) Exploring the taxonomy and phylogeny of Sordariomycetes taxa emphasizing Xylariomycetidae in Southwestern China. Mycosphere 15(1): 1675–1793. 10.5943/mycosphere/15/1/12 · doi ↗
- 4Du HZ, Chi MF, Wu N, Dissanayake AJ, Liu NG, Cheewangkoon R, Liu JK (2025) Taxonomic and phylogenetic contributions to Dothideomycetes and Sordariomycetes associated with medicinal plants in Southwestern China. Mycosphere 16(2): 179–343. 10.5943/mycosphere/16/2/2 · doi ↗
- 5Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for Basidiomycetes: Application to the identification of mycorrhizae and rusts. Molecular Ecology 2: 113–118. 10.1111/j.1365-294X.1993.tb 00005.x 8180733 · doi ↗ · pubmed ↗
- 6Glass NL, Donaldson GC (1995) Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Applied and Environmental Microbiology 61: 1323–1330. 10.1128/AEM.61.4.1323-1330.1995 PMC 1673887747954 · doi ↗ · pubmed ↗
- 7Habib K, Li WH, Ren YL, Liu LL, Lu CT, Zhang QF, Yao ZQ, Luo XY, Zhou X, Zeng WY, Kang YQ, Shen XC, Wijayawardene NN, Elgorban AM, Al N/A Rejaie S, Li QR (2025) Exploration of ascomycetous fungi revealing novel taxa in Southwestern China. Mycosphere 16: 1412–1529. 10.5943/mycosphere/16/1/9 · doi ↗
- 8Hongsanan S, Phookamsak R, Bhat DJ, Wanasinghe DN, Hyde KD, Samarakoon MC, Karunarathna SC, Dissanayake LS, Tibpromma S, Jayawardena RS, Tang X, Xu JC, Mortimer PE (2023) Exploring ascomycete diversity in Yunnan, China I: Resolving ambiguous taxa in Phaeothecoidiellaceae and investigating conservation implications of fungi. Frontiers in Cellular and Infection Microbiology 13: 1252387. 10.3389/fcimb.2023.1252387 PMC 1051306237743866 · doi ↗ · pubmed ↗