Targeting dCas9‐SunTag to a Susceptibility Gene Promoter Is Sufficient for CRISPR Interference
Zuh‐Jyh Daniel Lin, Gabriela L. Hernandez, Myia K. Stanton, Xingguo Zheng, Kerrigan B. Gilbert, Kira M. Veley, Greg Jensen, Marisa Yoder, Suhua Feng, Basudev Ghoshal, Jason Gardiner, Ming Wang, Steven E. Jacobsen, James C. Carrington, Rebecca S. Bart

TL;DR
This paper shows that using CRISPR interference to target susceptibility genes in cassava can reduce disease caused by viruses.
Contribution
The study demonstrates that CRISPR interference, not DNA methylation, is sufficient to reduce gene expression in cassava brown streak disease.
Findings
Targeting the promoters of nCBP-1 and nCBP-2 with a dCas9-DRMcd-SunTag system reduced gene expression.
CRISPR interference, rather than DNA methylation, was found to be responsible for the observed gene silencing.
Future research will test if methylation alone can confer resistance to cassava brown streak disease.
Abstract
Cassava production in sub‐Saharan Africa is severely impacted by diseases. Most pathogens require interaction with host susceptibility factors to complete their life cycles and cause disease. Targeted DNA methylation is an epigenetic strategy to alter gene expression in plants, and we previously reported that a zinc‐finger fused to DMS3 could establish methylation at the promoter of MeSWEET10a, a bacterial susceptibility gene, and this resulted in decreased disease. Here, we attempt a similar strategy for cassava brown streak disease. This disease is caused by the ipomoviruses CBSV and UCBSV. These viruses belong to the family Potyviridae, which has been shown extensively to require host eIF4E‐family proteins to infect plants and cause disease. We previously found that cassava plants with simultaneous knockout mutations in two eIF4E genes, nCBP‐1 and nCBP‐2, resulted in decreased…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
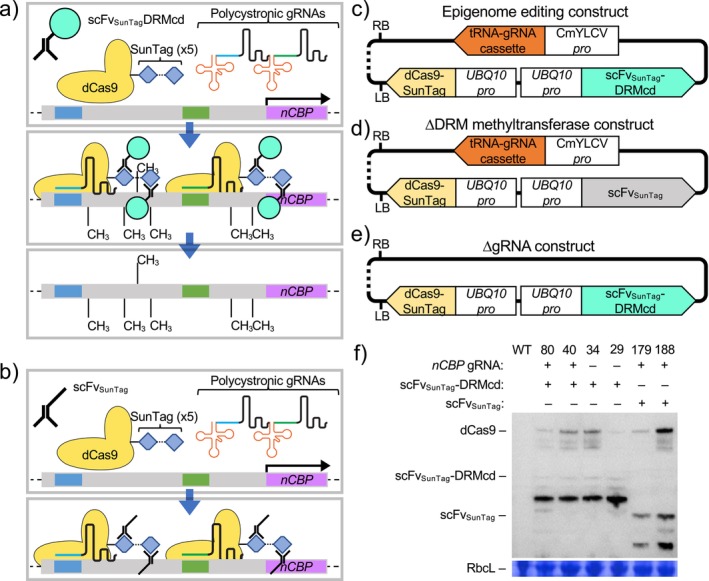 FIGURE 1
FIGURE 1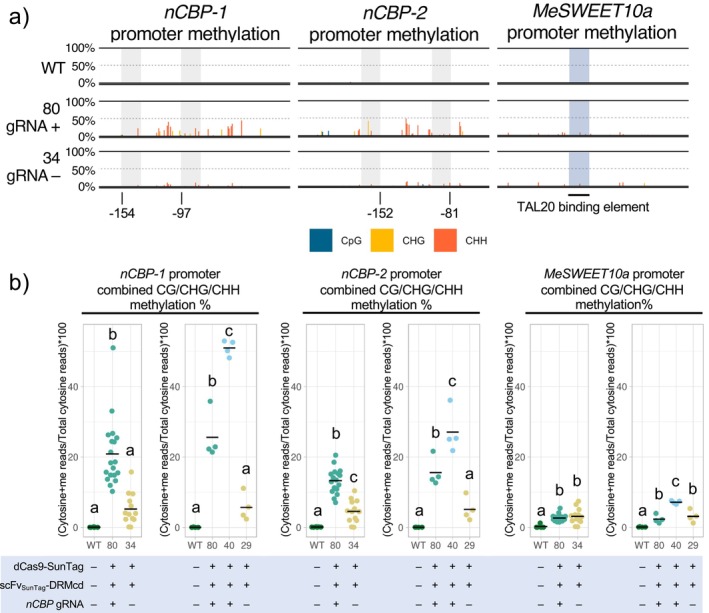 FIGURE 2
FIGURE 2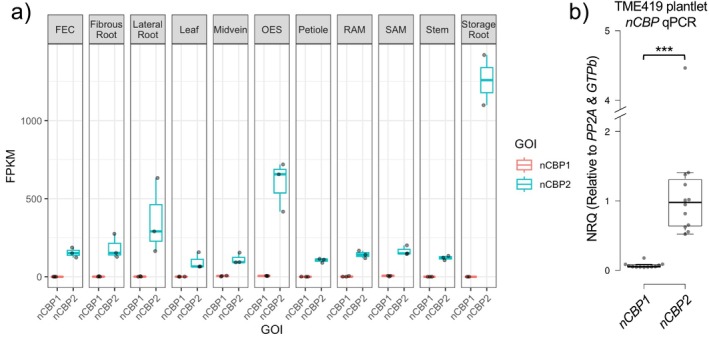 FIGURE 3
FIGURE 3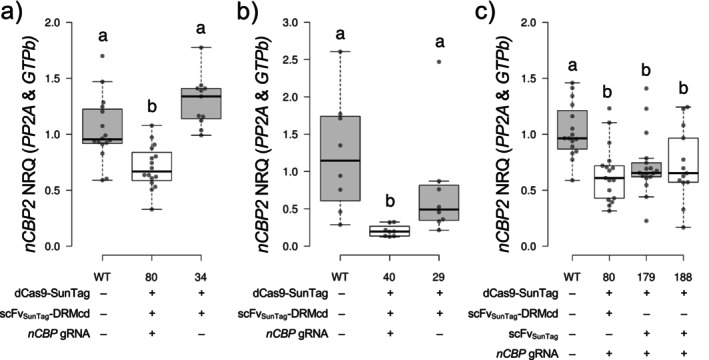 FIGURE 4
FIGURE 4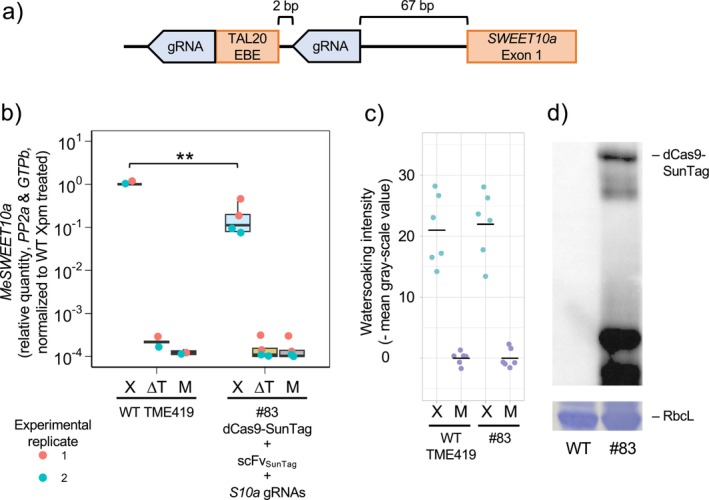 FIGURE 5
FIGURE 5- —Bill and Melinda Gates Foundation10.13039/100000865
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Virus Research Studies · CRISPR and Genetic Engineering · Cassava research and cyanide
Introduction
1
In plant–pathogen interactions, the manipulation of host processes by pathogens at the molecular level is critical for establishment of disease. These molecular interactions can involve interference with immune responses and co‐opting or positively regulating host processes for the pathogen's benefit. Host proteins that pathogens co‐opt and use to promote infection are known as susceptibility factors. Some examples of well‐studied genes encoding susceptibility (S) factors include MLO for powdery mildew (Büschges et al. 1997; Kusch and Panstruga 2017), SWEET gene products for xanthomonads (Yang et al. 2006; Cohn et al. 2014; Mormile et al. 2024), and the eIF4E‐family members for viruses of Potyviridae (Duprat et al. 2002; Lellis et al. 2002; Zaidi et al. 2018; Garcia‐Ruiz et al. 2021). Disrupting pathogen interaction with susceptibility factors can confer disease resistance. This has previously been achieved through mutant S gene alleles or by using RNA interference (RNAi) to knock down S gene expression (Bastet et al. 2017; Takakura et al. 2018; Zaidi et al. 2018; Rubio et al. 2019; Garcia‐Ruiz et al. 2021).
The tropical root crop cassava is a staple for roughly 800 million people worldwide (Food and Agriculture Organization of the United Nations 2018). Nearly half of cassava consumers reside in sub‐Saharan Africa, where three major diseases—cassava bacterial blight (CBB), cassava mosaic disease (CMD), and cassava brown streak disease (CBSD)—threaten cassava production (Bart and Taylor 2017). CMD can be mitigated through the planting of resistant cultivars, but such cultivars are not widely available for CBB or CBSD (Fanou et al. 2018; Chikoti and Tembo 2022). In cassava, MeSWEET10a and two pectate lyases are known S genes for CBB (Cohn et al. 2014, 2016). DNA polymerase delta subunit 1 was recently described as an S gene for CMD (Wu et al. 2021; Lim et al. 2022), and CBSD requires interaction with two *eIF4E‐*family genes, nCBP‐1 and nCBP‐2 (Gomez et al. 2019). We previously demonstrated that CRISPR‐mediated mutagenesis at the MeSWEET10a promoter increased resistance to CBB (Elliott et al. 2024), and simultaneous knockout of nCBP‐1 and nCBP‐2 attenuated CBSD symptoms, including storage root necrosis (Gomez et al. 2019). For CBB, a similar disease resistance outcome was achieved by directing DEFECTIVE IN MERISTEM SILENCING 3 (DMS3) to the MeSWEET10a promoter, resulting in de novo DNA methylation and suppression of pathogen‐mediated gene induction (Veley et al. 2023).
CBSD is caused by independent or simultaneous infection by two potyvirids of the genus Ipomovirus, cassava brown streak virus (CBSV) and Ugandan cassava brown streak virus (UCBSV). The disease manifests as feathery vein chlorosis in leaves, brown streaking of stems, and corky necrosis that destroys the economically important storage roots (Tomlinson et al. 2018). Although simultaneous knockout of nCBP‐1 and nCBP‐2 in the historical cultivar 60,444 resulted in decreased disease compared to controls, these lines were not fully resistant. This lack of full resistance is possibly due to *eIF4E‐*family redundancy, a hypothesis supported by protein–protein interaction data that demonstrates CBSV VPg can associate with the entire cassava complement of eIF4E‐family encoded proteins (Gomez et al. 2019). Knocking out the entire *eIF4E‐*family for CBSD resistance is infeasible due to their necessity for endogenous host protein translation. Knockdown, opposed to knockout, of susceptibility factors has been well documented to control potyviruses that exhibit eIF4E‐ or *eIF(iso)4E‐*sub clade dependency (Bastet et al. 2017; Takakura et al. 2018; Rubio et al. 2019). However, the transferability of this method towards ipomovirus control is an open question, especially given potential differences in infection strategies used by potyviruses and ipomoviruses (Dombrovsky et al. 2014).
Our previous success with directing DNA methylation to the MeSWEET10a promoter inspired us to attempt epigenetic knockdown of members of the eIF4E‐family genes in cassava. Here, we use a deactivated Cas9 protein to target the DOMAINS REARRANGED METHYLTRANSFERASE (DRM) catalytic domain to both nCBP‐1 and nCBP‐2 promoters for epigenome editing (Papikian et al. 2019). The resulting lines showed decreased target gene expression. However, control lines lacking the DNA methylation reagent showed a comparable decrease in gene expression, suggesting that dCas9 alone interferes with transcriptional machinery. This CRISPR interference (CRISPRi) effect has previously been reported (Qi et al. 2013; Gilbert et al. 2014; Gardiner et al. 2022). Thus, the important next step will be to segregate away the DNA methylation reagents through genetic crosses and assess whether methylation is heritable and, if so, sufficient for increased resistance to CBSD.
Results
2
Production of Transgenic Cassava to Knockdown Susceptibility Factor Gene Expression
2.1
We previously demonstrated that it is possible to direct methylation to a target region of the cassava genome (Veley et al. 2023). We hypothesized that targeting methylation to the nCBP‐1 and ‐2 promoters would result in gene knockdown. To test this hypothesis, we used an epigenome editing tool composed of three components, all present on a single T‐DNA: (1) deactivated Cas9 with a C‐terminally fused SunTag epitope (dCas9‐SunTag), (2) Nicotiana tabacum DRM methyltransferase catalytic domain with its N‐terminus fused to a single‐chain variable fragment antibody that binds the SunTag epitope (scFv_SunTag_‐DRMcd), and (3) a Pol II promoter driven tRNA‐gRNA array (Figure 1a,c) (Čermák et al. 2017; Papikian et al. 2019). This tool has been used to re‐establish heritable promoter methylation and subsequent repression of FWA at the fwa‐4 epiallele in Arabidopsis. Versions of this plasmid that omit either DRMcd (∆DRMcd) or the gRNAs (∆gRNA) were used as CRISPR interference (CRISPRi) and off‐target methylation controls, respectively (Figure 1d,e). The CRISPRi construct was also evaluated for gene knockdown activity (Figure 1b).
Strategy for dCas9‐mediated gene knockdown. (a,b) Hypothesized mechanism of action for transcriptional knockdown via epigenome editing (a) or CRISPRi (b). (c–e) Binary vector diagrams with color‐coded genes for dCas9‐SunTag guided de novo methylation of DNA by scFvSunTag‐DRMcd. (f) Transgene‐encoded protein accumulation in all transgenic lines used in this study. HA‐tagged dCas9‐SunTag, scFvSunTag‐DRMcd, and scFvSunTag are all detected by western blot. Rubisco large subunit (RbcL) detected using Coomassie brilliant blue staining of the same blot is presented as a loading control.
Two guide RNAs were used for each gene, nCBP‐1 and nCBP‐2. Guide RNAs were designed to bind on either side of the putative first nucleotide of the respective 5′ UTR (based on Phytozome Version 6) (shaded boxes in Figure 2a) (Bredeson et al. 2016). Following transformation into farmer‐preferred cassava cultivar TME419, six relevant transgenic lines were recovered: ∆DRMcd lines #179 and #188; ∆gRNA lines #29 and #34; and lines containing the experimental epigenome‐editing construct, #40 and #80. These lines were specifically selected for further study as the transgene‐encoded proteins expressed in control lines are at similar or greater levels than seen in experimental lines (Figure 1f).
Targeting DRMcd to nCBP promoters produces modest levels of de novo CHH and CHG methylation. (a) Amplicon bisulfite sequencing of roughly 140–190 base pair windows in the nCBP‐1, nCBP‐2, and SWEET10a promoters. These windows encompass the transcriptional start site, and the right most nucleotide positions are −13, −69, and +1 base pair(s) relative to the first nucleotide of exon one, respectively. gRNA locations are shaded gray with the 5′ most nucleotide position relative to the start codon annotated. The TAL20 binding element of the SWEET10a promoter is shaded blue. Line #80 and #34 both express dCas9‐SunTag and scFvSunTag‐DRMcd, but only #80 also expresses a gRNA cassette targeting both nCBP‐1 and nCBP‐2. (b) Comparison of binned methylated‐cytosine (cytosine + me) read percentage [100 × (methylated cytosine reads/total cytosine reads)] from within the BS‐PCR seq window. Horizontal bar denotes mean methylation percentage. Statistical differences were detected by one‐way ANOVA and post hoc Tukey's HSD, ⍺ = 0.05.
Targeted Methylation to the nCBP Promoters
2.2
Following identification of cassava transgenic lines carrying either the experimental or control vectors, we used amplicon‐based bisulfite sequencing (ampBS‐seq) to characterize the methylation patterns within the nCBP promoter regions. Wild‐type TME419 plants exhibited no methylation within the window of interest, whereas de novo methylation in the CHH and CHG contexts was observed in transgenic lines expressing dCas9‐SunTag with the DRM catalytic domain (Figures 2a,b, S1, and S2). Specifically, in lines #40 and #80, which contain gRNAs to target the dCas9‐based machinery to the nCBP promoters, we observed significantly higher levels of cytosine methylation than in the control lines lacking gRNAs (lines #29 and #34) (Figures 2a,b, S1, and S2).
To further confirm that the de novo methylation in lines #40 and #80 was specifically targeted to the nCBP regions, we characterized a third locus, the promoter of MeSWEET10a, a susceptibility factor for cassava bacterial blight disease. MeSWEET10a was chosen as we had previously characterized its promoter‐methylation status using ampBS‐seq (Veley et al. 2023). The MeSWEET10a promoter displayed weak levels of de novo methylation in all transgenic lines (Figure 2b and S3), indicating that although the presence of ectopic DRM catalytic domain is sufficient for low levels of nonspecific de novo methylation, these levels are lower than when the enzyme is specifically targeted to a locus via gRNA‐based recruitment of dCas9‐SunTag.
dCas9‐SunTag Occupancy of nCBP‐2 Promoter Results in Transcriptional Knockdown
2.3
As promoter methylation is usually associated with downregulation of gene expression, classically demonstrated at the FWA locus in Arabidopsis (Soppe et al. 2000; Bewick and Schmitz 2017; Zhang et al. 2018), we investigated if targeting de novo methylation to the nCBP‐1 and nCBP‐2 promoters could downregulate these susceptibility genes. However, we experienced difficulty in detecting nCBP‐1 in wild‐type plants via qRT‐PCR (Figure S4a). Closer inspection of the nCBP‐1 gene model revealed that it was merged with the gene immediately downstream, Manes.09G140200 (Wilson et al. 2017; Gomez et al. 2019). Taking the correct, un‐merged gene model into account, nCBP‐1 expression is low or undetectable in all analyzed tissues, including cassava storage roots, stems, and leaves (Figure 3a). In these tissues where nCBP‐1 transcripts were previously detected by RNAseq, nCBP‐2 is at least 30‐fold more abundant. To confirm the expression of nCBP‐2 relative to nCBP‐1, tissue culture plantlet leaves were analyzed by qPCR, revealing that mean nCBP‐2 expression was 16‐fold greater than that of nCBP‐1 (Figure 3b). Furthermore, 75% of nCBP‐1 cycle quantification values observed over multiple experiments are above 30, a consistent signature of low abundance transcripts that are difficult to quantify via qPCR (Taylor et al. 2019) (Figure S4). Given this result, we focused subsequent expression analyses on nCBP‐2 only.
nCBP‐1 is weakly or not expressed in most tissue types. (a) nCBP‐1 and nCBP‐2 expression data extracted from the Bart Lab Cassava Atlas RNAseq browser. Gene expression levels are reported as fragments per kilobase of exon per million reads mapped (FPKM). (b) qRT‐PCR was used to detect nCBP‐1 and nCBP‐2 normalized relative quantities (NRQs), relative to PP2A and GTPb, in plantlet leaves. The NRQs shown are relative to the geometric mean of nCBP‐2 measurements. Statistical differences were detected by Mann–Whitney U test, ɑ = 0.05, *** denotes p ≤ 0.001.
Expression of nCBP‐2 was significantly lower in epigenome edited lines #40 and #80 when compared to wild type and ∆gRNA lines (#29, #34) (Figure 4a,b). Therefore, *nCBP‐*2 knockdown is not due to off‐target methylation. However, nCBP‐2 expression was also repressed in the CRISPRi control lines #179 and #188, which contain dCas9‐SunTag, scFv_SunTag_, and the nCBP‐2 gRNAs but lack the DRM methylation domain, suggesting that nCBP‐2 knockdown in the epigenome edited lines may have been due to transcriptional interference (Figure 4c). Transcriptional repression due to steric hindrance by dCas9 of endogenous transcriptional machinery when targeted near transcriptional start sites has been previously observed in bacteria and human cells (Qi et al. 2013; Gilbert et al. 2014). Similarly, in plants, dCas9 has been fused to a transcriptional repressor domain for dCas9‐based, targeted knockdown of gene expression (Pan et al. 2021). Both approaches are a form of CRISPR interference (CRISPRi) (Qi et al. 2013; Pan et al. 2021). Our results indicate that gRNA‐based targeting of dCas9‐SunTag to a promoter region, even without fusion to a transcriptional repressor, is sufficient for significant knockdown of gene expression.
nCBP‐2 knockdown can be mediated by dCas9‐SunTag without DRMcd. nCBP‐2 expression levels in transgenic lines with and without gRNAs for targeted methylation (a,b) or with or without the DRMcd methyltransferase (c). qRT‐PCR was used to detect nCBP‐2 normalized relative quantities (NRQs) relative to PP2A and GTPb. The NRQs shown are relative to the geometric mean of wild‐type measurements. The Kruskal–Wallis H test and post hoc Dunn's test using a Bonferroni‐corrected ɑ were used in (a–c) to detect significantly different mean ranks between paired groups.
dCas9‐SunTag Mediates CRISPRi at MeSWEET10a
2.4
To assess the applicability of dCas9‐SunTag for CRISPRi at other loci, we tested if this system could inhibit induction of MeSWEET10a by the bacterial pathogen Xanthomonas phaseoli pv. manihotis (Xpm). Xpm secretes the effector transcription activator‐like20 (TAL20) into host cells to bind the MeSWEET10a promoter and activate its expression. This Xpm‐mediated induction of MeSWEET10a is necessary for development of watersoaking symptoms. We were able to produce one transgenic cassava line, #83, expressing dCas9‐SunTag/scFV_SunTag_ and two guide RNAs targeted to sequences immediately flanking the TAL20 effector binding element (Figure 5). qPCR analysis of MeSWEET10a in this line after inoculation with Xpm revealed a roughly 10‐fold reduction in MeSWEET10a expression relative to wild‐type plants (Figure 5b). This level of expression in line #83 was still several orders of magnitude greater than seen in plants inoculated with Xpm lacking TAL20 or mock inoculated with water. We proceeded to examine the effect of this on Xpm‐induced watersoaking and found no difference between wild type and line #83 plants (Figure 5c).
dCas9‐SunTag/scFvSunTag targeted to the TAL20 EBE inhibits Xpm‐mediated induction of MeSWEET10a. (a) Diagram for guide RNAs targeting dCas9‐SunTag/scFvSunTag to the MeSWEET10a promoter. (b) qPCR analysis of MeSWEET10a induction in wild‐type TME419 and a transgenic line (#83) expressing dCas9‐SunTag and scFVSunTag targeted to the TAL20 effector binding element in the MeSWEET10a promoter. Plants were treated with Xanthomonas phaseoli pv. manihotis (Xpm, X), Xpm with the TAL20 effector deleted (∆T), or mock (M). Results plotted are from two separate experiments. MeSWEET10a is quantified relative to PP2A and GTPb, then expressed relative to the Xpm‐treated wild‐type sample. Statistical differences were detected using a one sample T‐test, mu = 1. ** denotes a p value < 0.01. (c) Watersoaking severity analysis after treatment with Xpm (X) or mock (M) inoculation. Watersoaking intensity is measured as the mean negative grayscale value of an inoculated area normalized to the average of mock. (d) Representative western blot showing accumulation of HA‐tagged dCas9‐SunTag in line #83.
Discussion
3
We previously reported evidence that CBSV may recruit all cassava eIF4E‐family proteins for disease. However, knocking out all eIF4E‐family members simultaneously is not a viable strategy as these are critical host factors. We hypothesized that susceptibility factor knockdown may allow for the balancing of plant viability and disease severity. Although susceptibility factor knockdown has been demonstrated to control disease caused by many potyviruses, this strategy had yet to be demonstrated for an ipomovirus with such broad eIF4E‐family recruitment (Bastet et al. 2017; Takakura et al. 2018; Rubio et al. 2019). Here, we attempt to use targeted DNA methylation to knockdown expression of nCBP‐1 and *nCBP‐*2. Although we successfully achieved methylation at the target regions of the cassava genome, key control lines revealed that CRISPRi from the dCas9‐SunTag fusion protein is sufficient to cause gene expression knockdown. Whether the level of nCBP promoter methylation achieved in this study is sufficient for heritable epigenetic knockdown will be further investigated by segregating away the epigenome editing tools.
An epigenome editing method is attractive as it may allow for transgene‐free heritable knockdown of susceptibility factors. Thus far successful heritable epigenome editing has only been reliably accomplished at the fwa‐4 epiallele in Arabidopsis thaliana (Papikian et al. 2019; Dubois and Roudier 2021; Ghoshal et al. 2021). Although we were able to specifically enhance methylation at the nCBP promoters over wild‐type and ∆gRNA genetic backgrounds, the highest level of methylation observed at any cytosine for either nCBP promoter was 70% in the CHH context (Figure S1). Of the three possible methylation contexts, only CHH and CHG were observed. This is notable as CG methylation is thought to play a significant role in gene silencing and mediating inheritance of epigenetic marks (Ghoshal et al. 2021). The lack of CG methylation at the nCBP promoters in lines #40 and #80 is surprising given that our epigenome tool utilizes the catalytic domain of DRM, a de novo methyltransferase that methylates all three contexts (Law and Jacobsen 2010; Papikian et al. 2019). In contrast, our previous work targeting DMS3, a non‐catalytic component of RNA‐directed de novo methylation machinery, to the MeSWEET10a promoter resulted in near 100% methylation of CG residues within the DNA motif required for MeSWEET10a upregulation by X. phaseoli pv. manihotis (Veley et al. 2023). Promoter DNA methylation is not the only epigenetic mark associated with repression of gene expression; acetylation and methylation of certain lysine residues of histone 3 also control gene expression and have a complex interplay with DNA methylation (Du et al. 2015; Zhang et al. 2018). It is possible that existing histone epigenetic marks at the nCBP promoters antagonize DNA methylation, whereas those at the MeSWEET10a promoter are permissive of DNA methylation.
The majority of CRISPRi studies in plants have focused on the development of dCas9‐transcriptional repressor fusions (Lowder et al. 2015; Piatek et al. 2015; Vazquez‐Vilar et al. 2016; Pan et al. 2021; Gardiner et al. 2022; Bhuyan et al. 2023; Wang et al. 2024). dCas9 alone has been shown to function as a transcriptional repressor in Agrobacterium‐mediated transient assays in Nicotiana benthamiana but has yet to be demonstrated in stable transgenic plants (Piatek et al. 2015; Vazquez‐Vilar et al. 2016; Wang et al. 2024). Our approach differs from these transcription factor free fusions in that we use dCas9 fused to 10 copies of the SunTag (GCN4) epitope that recruits 10 copies of a single‐chain variable fragment antibody fused to GFP (scFv_SunTag_). Furthermore, we simultaneously utilize two gRNAs in our target promoters. Our data clearly demonstrate that dCas9‐SunTag/scFv_SunTag_ targeted to the transcriptional start site of nCBP‐2 can downregulate its expression (Figures 1b and 4c). Furthermore, we also demonstrate that this tool can interfere with TAL effector mediated induction of the MeSWEET10a susceptibility gene (Figure S5). These effects are likely due to CRISPRi. It is worth noting, however, that the transcriptional inhibition mechanism for each of these two examples may not be the same; one case involves endogenous regulation of gene expression, and the other involves interfering with DNA binding by a pathogen‐derived transcription factor. Additional work will be necessary to establish the general utility of this tool at other loci across plant systems.
It also remains to be seen whether nCBP‐2 knockdown is sufficient for attenuating CBSD symptoms. However, a knockdown strategy for CBSD resistance may provide some advantages compared to a traditional genome editing approach; the eIF(iso)4E and nCBP clades in cassava each consist of two homologs, whereas eIF4E has just one. In Arabidopsis, eIF4E and eIF(iso)4E are individually dispensable for plant viability, but simultaneous mutations in both are lethal (Patrick et al. 2014). Assuming one intact eIF4E or eIF(iso)4E allele is required for viability, combining additional eIF4E and eIF(iso)4E mutations with those in the nCBPs for resistance requires several simultaneous CRISPR induced frameshift mutations all while preserving at least one eIF4E/eIF(iso)4E allele. Such a scenario may be difficult to obtain as insertion and deletion mutations induced by CRISPR are not always frameshifting, homozygous, or bi‐allelic. Expanding our CRISPRi approach by targeting dCas9‐SunTag or a dCas9‐transcriptional repressor to additional eIF4E‐family member promoters of interest would be an alternative method. For other purposes, CRISPRi and/or promoter methylation may prove advantageous compared to traditional RNAi‐mediated knockdown in that dCas9 is targeted upstream of coding sequences, thus allowing for specific targeting of genes with close homologs sharing high sequence similarity. If CRISPRi were to be pursued further as an experimental or disease control strategy, future work might compare dCas9, dCas9‐SunTag/scFv_SunTag_, and dCas9‐repressor efficacy across multiple loci and also in the context of gRNA positional and dosage associated effects. As for targeted methylation, whereas this work clearly demonstrates feasibility of targeting specific loci with methylation, future work will need to understand heritability of de novo methylation and impact on gene expression in the absence of the DNA methylation reagents.
Methods/Experimental Procedures
4
Plant Transformation, Growth Conditions, and Disease Assays
4.1
Transformation of cassava cultivar TME419 was performed as previously described (Veley et al. 2023). Transgenic lines were then maintained as in vitro plantlets (Taylor et al. 2012). For greenhouse growth, in vitro plantlets were moved to soil and allowed to establish on a misting bench before transfer to greenhouse conditions (Veley et al. 2023). Watersoaking assays were carried out as described in (Veley et al. 2023).
Plasmid Construction
4.2
gRNAs targeting the nBCP promoters (nCBP‐1: CCGATGAATAAGAGCGCTAG, CGCTCTCAACTGTACTTCAT, nCBP‐2: CACTCGATTGCAGATTTTTG, AGACGATGAAAGAGCCGAAG) were designed using CRISPOR (Concordet and Haeussler 2018). gRNAs were assembled into pDIRECT_21C as described by Čermák et al. (2017). The CmYLCVpro::tRNA‐gRNA array was then amplified by PCR and cloned into the KpnI site of the dCas9‐SunTag/scFv_SunTag_‐DRMcd epigenome editing binary vector. The dCas9‐SunTag/scFv_SunTag_ constructs were derived from dCas9‐SunTag/scFv_SunTag_‐DRMcd by utilizing BsiWI sites immediately flanking the DRMcd sequence.
Methylation Analysis
4.3
Cassava genomic DNA was extracted using the DNeasy Pro Mini Kit (Qiagen) and then prepared and processed for ampBS‐seq as previously described (Veley et al. 2023). Primer sequences are provided in Table S1. Raw paired sequencing reads were merged and aligned to the following nCBP promoter sequences in CLC Genomics Workbench (Qiagen): nCBP‐1, CCGATGAATAAGAGCGCTAGAGGTGAACGGTTTTCACCTGTCATCCTCTCTGGTTTCCGCTCTCAACTGTACTTCATAGGAAGCTATACTCGAATACAGAGACCTCCTATTAGAGCTTTTAGGATTTGGGGTTCAGCAAAG; nCBP‐2, CCGAGTGCGGGAAATTTTATCACCGTTTCCAAACAGGCCACAAAAATCTGCAATCGAGTGAAATAAATGAAGCGTGTGTAGCTTTCCACTTCCTTTTCTCTCCGGTTTCCTCTTCGGCTCTTTCATCGTCTTCGCTGGACCT. Methylated cytosine reads were then called using the CLC Genomics Workbench epigenomic analysis plug‐in.
nCBP and MeSWEET10a Expression Analysis
4.4
In vitro plantlet or greenhouse cassava leaf tissue was used for qPCR analysis of nCBP‐1 and nCBP‐2 expression. For in vitro plantlets, RNA was extracted using TRIzol (Invitrogen) in conjunction with the Direct‐zol RNA Miniprep Kit (Zymo). For greenhouse grown plants, RNA was extracted using the Spectrum Plant Total RNA Kit (Sigma). cDNA was synthesized using the SuperScript III First‐Strand Synthesis System (Invitrogen). qPCR was performed with SYBR Select Master Mix (Thermo Fisher) on CFX384 Touch (Bio‐Rad) and QuantStudio 5 (Applied Biosystems) Real‐Time PCR Systems. nCBP and MeSWEET10a normalized relative quantities were calculated and analyzed as described in Gomez et al. (2019) and Veley et al. (2023). Primers used for qPCR are described in Table S1.
Author Contributions
Conceptualization: J.C.C., S.E.J., and R.S.B. Methodology: Z.D.L., X.Z., K.V., S.F., B.G., J.G., M.W., J.C.C., S.E.J., and R.S.B. Investigation: Z.D.L., G.L.H., M.K.S., X.Z., K.V., G.J., M.Y., S.F., B.G., and J.G. Visualization: Z.D.L., K.B.G., and M.K.S. Writing – original draft: Z.D.L.; Writing – review and editing: Z.D.L., G.L.H., M.K.S., X.Z., K.B.G., S.F., B.G., J.G., M.W., J.C.C., S.E.J., and R.S.B.
Funding
This work was funded by the Bill and Melinda Gates Foundation (OPP1125410 to R.B.S., S.E.J., and J.C.C.).
Conflicts of Interest
The authors declare no conflicts of interest.
Accession Numbers
nCBP‐1, Manes.09G140300; nCBP‐2, Manes.08G145200; GTPb, Manes.09G086600; PP2a, Manes.09G039900; MeSWEET10a, Manes.06G123400.
Supporting information
Figure S1: nCBP‐1 promoter cytosine methylation landscape. Percent methylated cytosines across the nCBP‐1 promoter ampBS‐seq window for epigenome edited transgenic line #80 and the ΔgRNA control line #34. Each row within a genotype corresponds to an independent biological replicate. Figure S2: nCBP‐2 promoter cytosine methylation landscape. Percent methylated cytosines across the nCBP‐2 promoter ampBS‐seq window for epigenome edited transgenic line #80 and the ΔgRNA control line #34. Each row within a genotype corresponds to an independent biological replicate. Figure S3: MeSWEET10a promoter cytosine methylation landscape. Percent methylated cytosines across the MeSWEET10a promoter ampBS‐seq window for epigenome edited transgenic line #80 and the ΔgRNA control line #34. Each row within a genotype corresponds to an independent biological replicate. Figure S4: (a) Observed cycle quantitation (Cq) values for nCBP1 and nCBP2 across from across five qPCR experiments examining tissue culture leaf, greenhouse leaf, and CBSV‐infected storage root material. Threshold values for Cq selection were chosen based on those automatically selected by qPCR software for nCBP‐2 and then applied to nCBP‐1. (b) Differences in nCBP‐1 and nCBP‐2 Cq from each tissue sample used in (a).
Table S1: Primers used in this study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bart, R. S. , and N. J. Taylor . 2017. “New Opportunities and Challenges to Engineer Disease Resistance in Cassava, a Staple Food of African Small‐Holder Farmers.” P Lo S Pathogens 13: e 1006287.28493983 10.1371/journal.ppat.1006287 PMC 5426740 · doi ↗ · pubmed ↗
- 2Bastet, A. , C. Robaglia , and J.‐L. Gallois . 2017. “e IF 4E Resistance: Natural Variation Should Guide Gene Editing.” Trends in Plant Science 22: 411–419.28258958 10.1016/j.tplants.2017.01.008 · doi ↗ · pubmed ↗
- 3Bewick, A. J. , and R. J. Schmitz . 2017. “Gene Body DNA Methylation in Plants.” Current Opinion in Plant Biology 36: 103–110.28258985 10.1016/j.pbi.2016.12.007PMC 5413422 · doi ↗ · pubmed ↗
- 4Bhuyan, S. J. , M. Kumar , P. Ramrao Devde , et al. 2023. “Progress in Gene Editing Tools, Implications and Success in Plants: A Review.” Frontiers in Genome Editing 5: 1272678.38144710 10.3389/fgeed.2023.1272678 PMC 10744593 · doi ↗ · pubmed ↗
- 5Bredeson, J. V. , J. B. Lyons , S. E. Prochnik , et al. 2016. “Sequencing Wild and Cultivated Cassava and Related Species Reveals Extensive Interspecific Hybridization and Genetic Diversity.” Nature Biotechnology 34: 562–570.10.1038/nbt.353527088722 · doi ↗ · pubmed ↗
- 6Büschges, R. , K. Hollricher , R. Panstruga , et al. 1997. “The Barley Mlo Gene: A Novel Control Element of Plant Pathogen Resistance.” Cell 88: 695–705.9054509 10.1016/s 0092-8674(00)81912-1 · doi ↗ · pubmed ↗
- 7Čermák, T. , S. J. Curtin , J. Gil‐Humanes , et al. 2017. “A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants.” Plant Cell 29: 1196–1217.28522548 10.1105/tpc.16.00922 PMC 5502448 · doi ↗ · pubmed ↗
- 8Chikoti, P. C. , and M. Tembo . 2022. “Expansion and Impact of Cassava Brown Streak and Cassava Mosaic Diseases in Africa: A Review.” Frontiers in Sustainable Food Systems 6: 1076364. 10.3389/fsufs.2022.1076364/full. · doi ↗