ELF5 modulates casein synthesis in goat mammary epithelial cells via JAK2/STAT5 signaling pathway
Yuzhu Guo, Cunxia Ma, Tianxiang Meng, Tongtong Tu, Shuangshuang Cui, Yinghui Ling, Yunhai Zhang, Hongyu Liu, Ning Song

TL;DR
This study shows that ELF5 increases casein production in goat mammary cells by activating the JAK2/STAT5 signaling pathway, which could help improve goat milk quality.
Contribution
The study identifies the JAK2/STAT5 pathway as a novel mechanism by which ELF5 regulates casein synthesis in goat mammary epithelial cells.
Findings
ELF5 enhances cell viability and proliferation while inhibiting apoptosis in goat mammary epithelial cells.
ELF5 upregulates the expression of αS1-, αS2-, β-, and κ-casein genes.
JAK2/STAT5 signaling is essential for ELF5-mediated casein synthesis.
Abstract
Goat milk contains a high concentration of caseins which are beneficial to human health. Understanding the regulatory mechanisms of casein synthesis in goats contribute to improving milk quality. Epithelial-specific E74-like factor 5 (ELF5) is important in milk protein synthesis, although the molecular mechanism by which ELF5 regulates casein synthesis remains unclear. This study aims to investigate the regulatory roles of ELF5 on casein production in goat mammary epithelial cells (GMECs). Primary GMECs were isolated from goats and characterized using immunofluorescence and oil red O staining. The goat ELF5 gene overexpression vector and small interfering RNA were transfected into GMECs, respectively. Cell viability was accessed using cell counting kit-8 assay, and cell apoptosis was detected by flow cytometry. Genes involved in cell proliferation, apoptosis, and casein synthesis were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
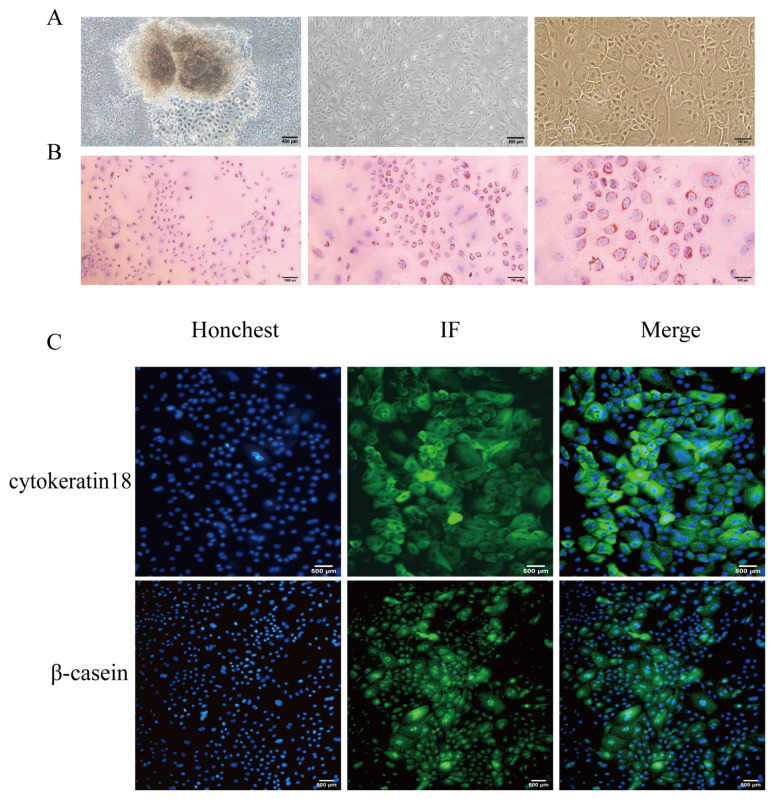 Figure 1
Figure 1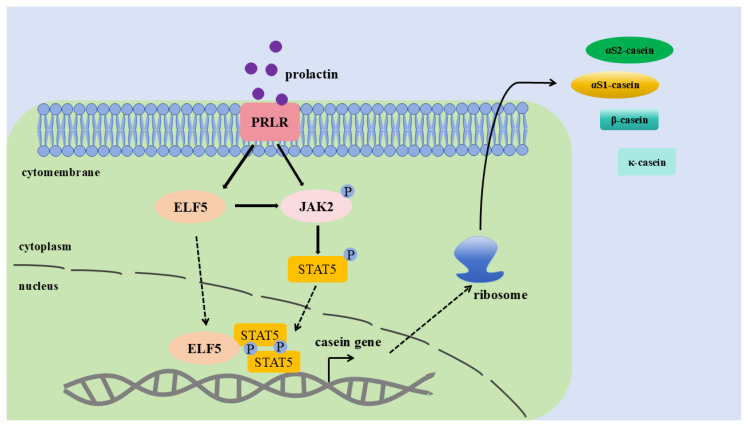 Figure 2
Figure 2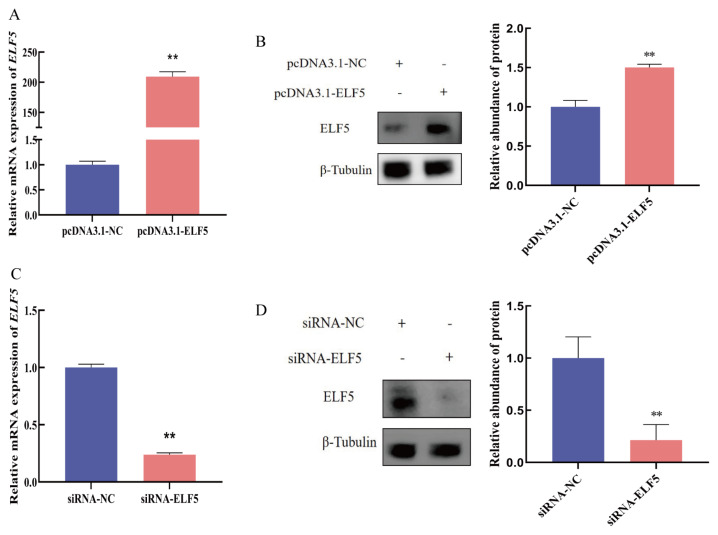 Figure 3
Figure 3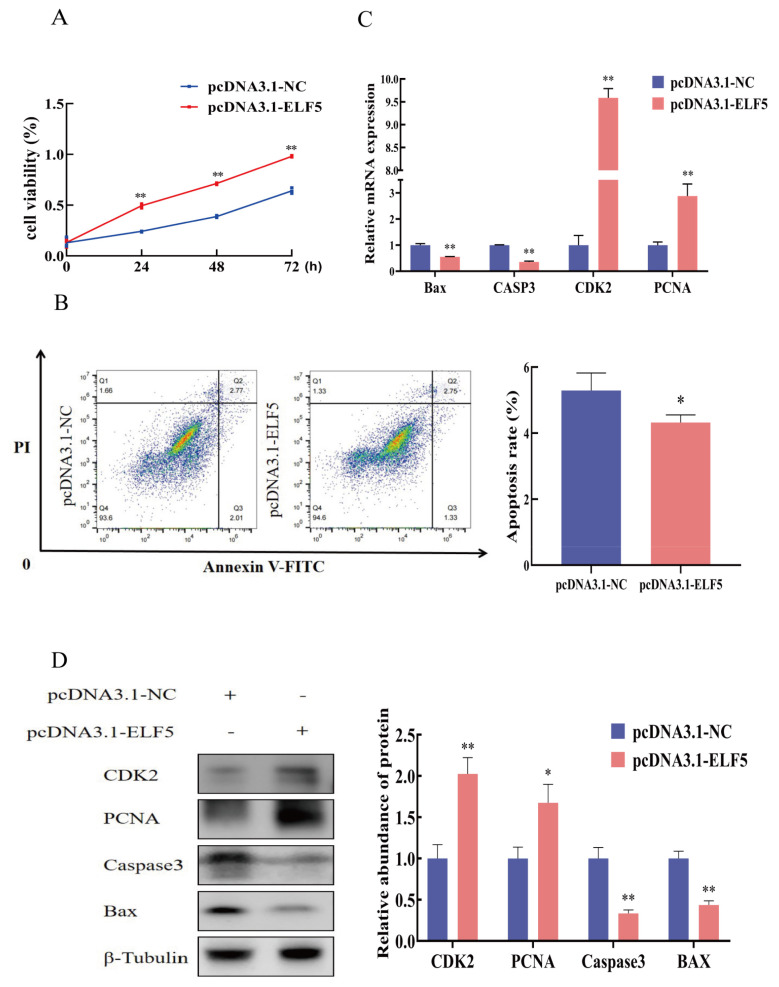 Figure 4
Figure 4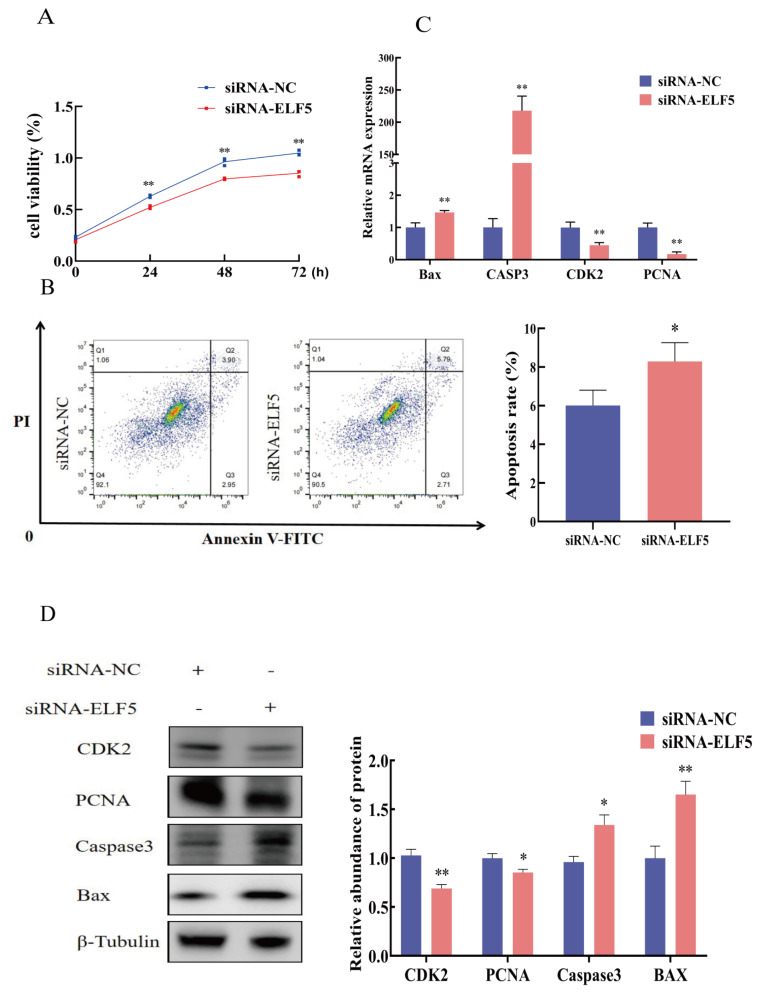 Figure 5
Figure 5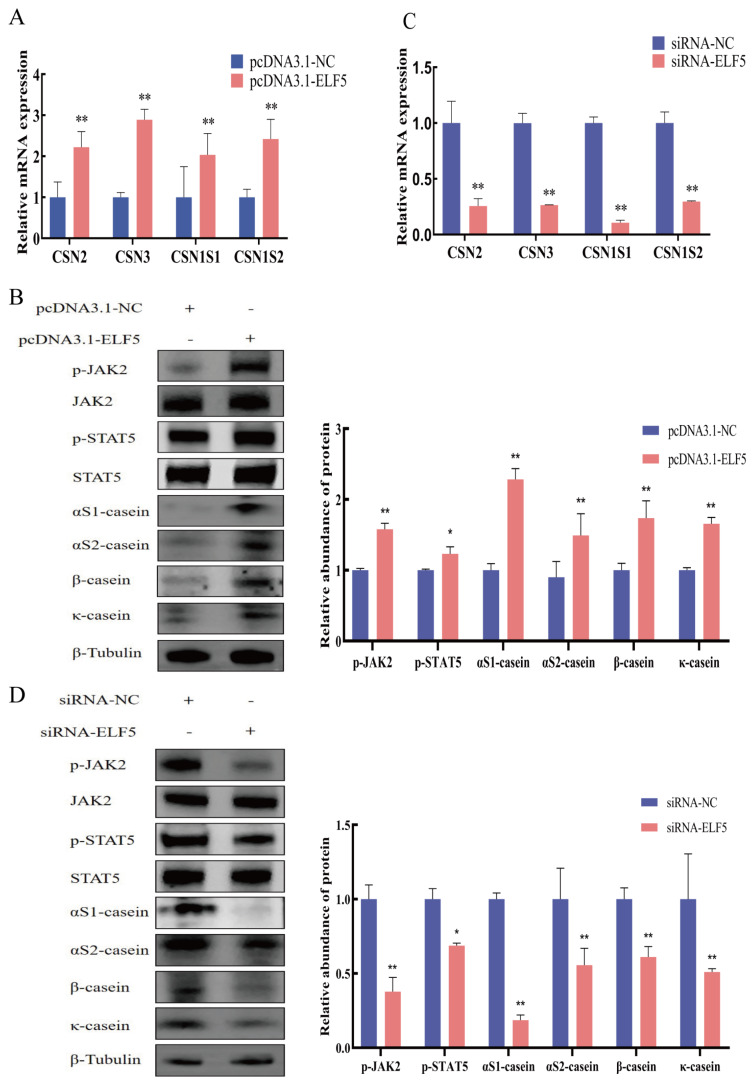 Figure 6
Figure 6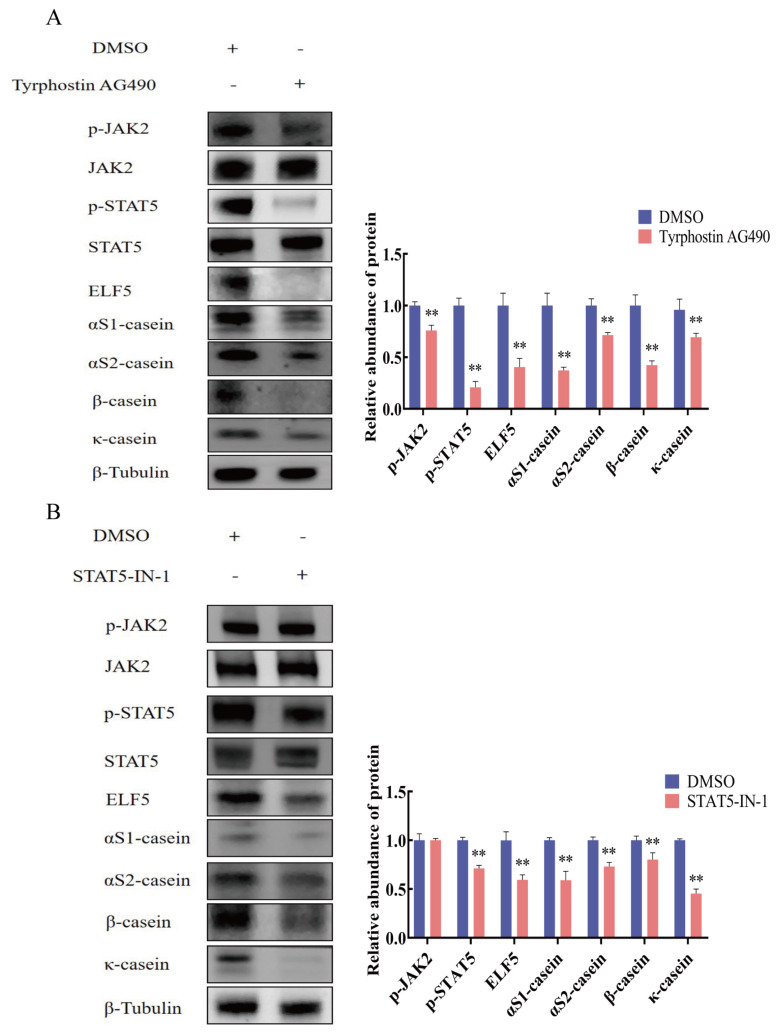 Figure 7
Figure 7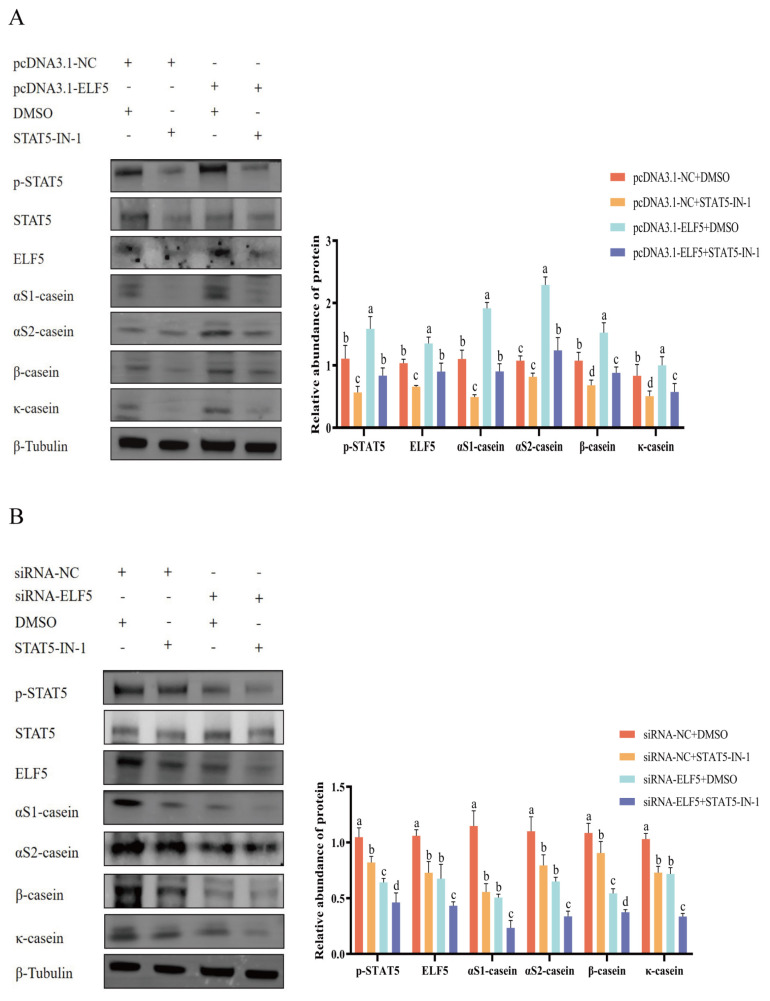 Figure 8
Figure 8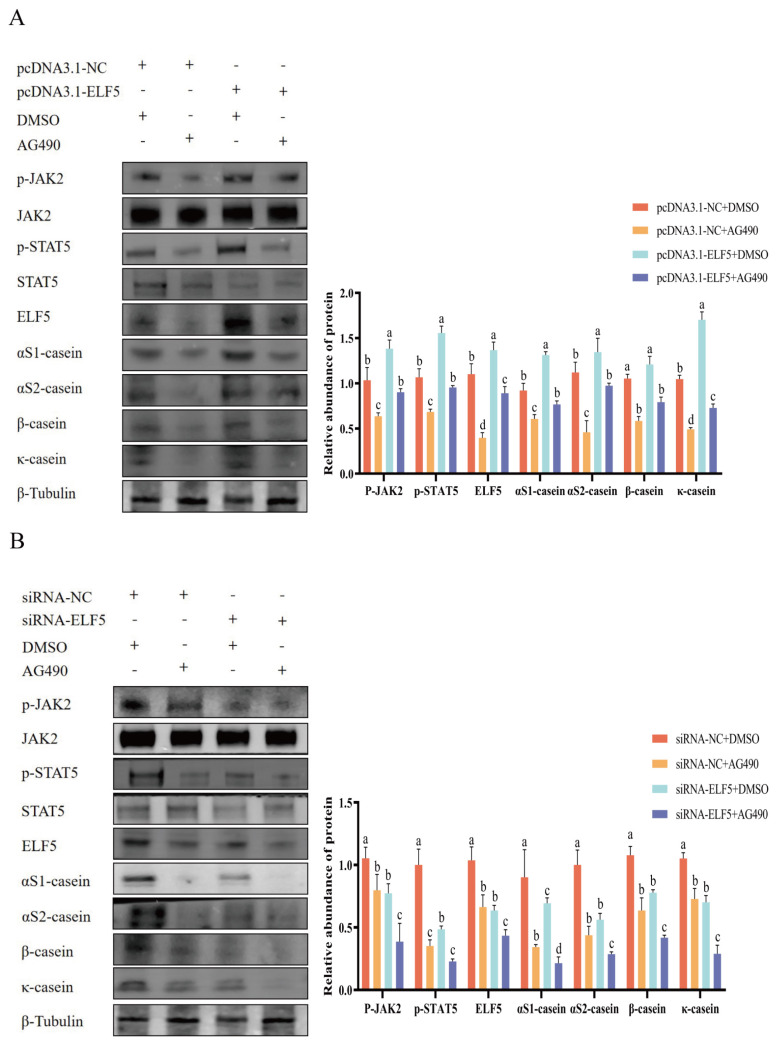 Figure 9
Figure 9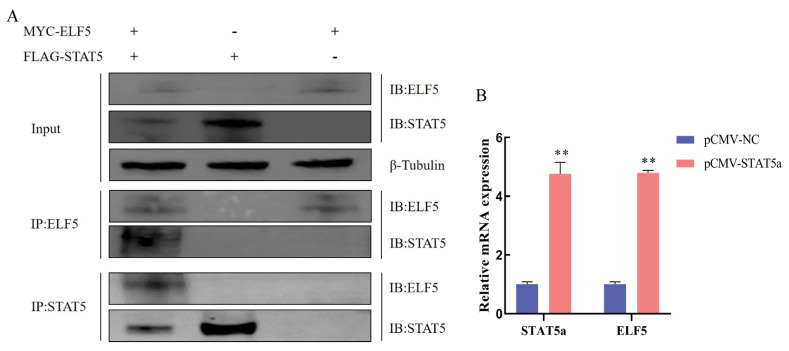 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIL-33, ST2, and ILC Pathways
INTRODUCTION
Goat milk protein contains essential amino acids and is an important source of dietary protein. Milk protein consist of casein and whey protein, with casein accounting for 80% of the total protein in goat milk. Casein is divided into αS1-casein (CSN1S1), αS2-casein (CSN1S2), β-casein (CSN2), and κ-casein (CSN3) [1]. αS1-Casein maintains the casein stability by binding to αS2-casein [2]. Bioactive peptides derived from β-casein contribute to the health of human cardiovascular, bone and immune system [3]. κ-Casein forms complexes with calcium ions, which enhances the nutritional value of dairy products [4]. These four types of casein furnish humans with high-quality protein sources. Therefore, to improve casein content in goat milk, it is necessary to analyze the molecular mechanism of casein synthesis in goat mammary epithelial cells (GMECs).
The synthesis of casein is a complex procedure that encompasses multiple signaling pathways and transcription factors in mammals. Janus kinase 2/signal transducer and activator of transcription 5 (JAK2/STAT5) and mechanistic target of rapamycin (mTOR) signaling pathways are crucial regulatory routes for governing the synthesis of casein [5]. After binding to membrane receptors of mammary epithelial cells, hormones such as prolactin activate JAK2 and catalyze downstream STAT5, then phosphorylated STAT5 activates the transcription of casein genes [6]. Amino acids activate mTOR pathway and initiate translation of casein genes [7]. E74-like factor 5 (ELF5), a member of the epithelial development-specific transcription factor, is associated with the regulation of casein synthesis [8]. The conserved deoxyribonucleic acid (DNA)-binding domain of ELF5 recognizes the promoter or enhancer regions to regulate target gene expression [9]. ELF5 participates in mammary gland development, placenta formation, maintenance of epithelial cell function, and milk protein synthesis [10].
To enhance the casein content in goat milk, it is indispensable to comprehend the regulatory function of ELF5 in casein synthesis. Previous studies showed that ELF5 facilitates the mammary gland development and milk protein content, it remains unclear whether ELF5 could regulate casein synthesis via JAK2/STAT5 signaling pathway. Our results furnish the molecular mechanism by which ELF5 regulates casein synthesis in goats, which offers a novel approach for enhancing the protein quality of ruminant milk.
MATERIALS AND METHODS
Cell culture
GMECs were isolated from three healthy Wanlin white goats at peak lactation period (2 years old, average of about 52 kg, second parturition, 45 d postpartum). The protocol for purification and cultured procedure as previously described [11]. Briefly, mammary tissue was collected using a surgical approach and washed with D-Hank’s solution. Tissue was then minced into about 1 mm^3^ cubes, plated in 60 mm culture dishes and cultured in 5% CO_2_ at 37°C. The medium was replaced every 2 days until the epithelial cells digested from the tissue block with 0.25% trypsin- ethylene diamine tetraacetic acid solution. Subsequently, fibroblasts were eliminated by differential adherence method, which adhered to culture dishes faster than GMECs. Cells were transferred to new culture dishes and purified 5 passages for subsequent experiments. The complete medium for GMECs was incorporated with 10% fetal bovine serum (11011-8615; Every Green) and 90% Dulbecco’s modified eagle medium/Ham’s F-12 (SH30023; Hyclone) medium with 10 μg/L epidermal growth factor (PHG0311; Invitrogen), 100 kU/L penicillin/streptomycin (080092569; Harbin Pharmaceutical), 5 mg/L bovine insulin (16634; Sigma-Aldrich), and 1 mg/L hydrocortisone (H0888; Sigma-Aldrich).
Identification of mammary epithelial cells
To determine the purity GMECs, immunofluorescence and oil red O staining were performed. Cells were washed with phosphate buffer solution, fixed with 4% paraformaldehyde, permeabilized with Triton, and blocked with 5% bovine serum albumin at room temperature for 1 h. Subsequently, the cells were incubated with Cytokeratin 18 (66187; Proteintech; 1:800) and β-casein antibody (bs-0466R; Bioss; 1:500) for 1 h in the dark. Thereafter, the cells were incubated with Alexa Fluor 488-conjugated goat anti-mouse immunoglobulin G (IgG) (CW0102; CWBIO; 1:5,000). To ascertain whether GMECs possess normal secretory function, cells were stained with the oil red O staining kit (G1262; Solarbio) and observed through a microscope.
Construction of overexpression vector and small interfering ribonucleic acid
Based on goat ELF5 mRNA sequence (GenBank accession no. XM_005690080.3), the forward primer and reverse primer are CGCGGATCCCTCACGGTGATGTTGGAC and GATGGAGCAGATCATAGCT, respectively. Employing cDNA samples derived from the mammary gland of goats at peak lactation. The coding sequence of the goat ELF5 gene was cloned into the BamHI/EcoRI sites of pcDNA3.1 using T4 DNA ligase (Takara), generating the pcDNA3.1-ELF5 expression vector. By utilizing Invitrogen online software, specific small interfering ribonucleic acid (siRNA) targeting goat ELF5 gene was designed, concurrently with an irrelevant negative control (NC) sequence siRNA-NC (Sangon, China). The sequence of siRNA-ELF5 is as follows: sense: GCAGAGCUCUGAGGUACUATT, antisense: UAGUACC UCAGAGCUCUGCTT. The sequence of siRNA-NC is: sense: UUCUCCGAACGUGUCACGUT, antisense: ACG UGACACGUUCGGAGAATT.
Cell transfection
For ELF5 gene overexpression assay, pcDNA3.1-NC or pcDNA3.1-ELF5 was transfected into GMECs by Lipofectamine 3000 (Invitrogen). For the knockdown of ELF5 gene, siRNA-NC or siRNA-ELF5 (100 nM) was transfected into cells by Lipofectamine RNAiMAX (Invitrogen). For the inhibition of JAK2/STAT5 pathway, JAK2 inhibitor AG490 (30 μM, MedChemExpress) or STAT5 inhibitor STAT5-IN-1 (50 μM, MedChemExpress) were treated with cells. For rescue experiment with STAT5 inhibitor and ELF5 overexpression, cells seeded in 6-well plates were grown to 80% confluence. After pre-treatment with STAT5-IN-1 (50 μM) for 1 h, transfection was performed with pcDNA3.1-NC or pcDNA3.1-ELF5 using Lipofectamine 3,000 according to the manufacturer’s protocol. Total protein was collected 48 h after transfection for Western blot analysis. Similarly, after pre-treatment with STAT5-IN-1 (50 μM) for 1 h, transfection was performed with siRNA-NC or siRNA-ELF5 (100 nM) using Lipofectamine RNAiMAX for 48 h. For rescue experiment with JAK2 inhibitor and ELF5 overexpression, cells seeded in 6-well plates were grown to 80% confluence. After pre-treatment with AG490 (30 μM) for 1 h, transfection was performed with pcDNA3.1-NC or pcDNA3.1-ELF5 using Lipofectamine 3,000 for 48 h. Similarly, after pre-treatment with AG490 (30 μM) for 1 h, transfection was performed with siRNA-NC or siRNA-ELF5 (100 nM) using Lipofectamine RNAiMAX for 48 h. Besides, for the interaction assay of ELF5 and STAT5, cells were transfected with pCMV-STAT5a or pCMV-NC. After 48 h treatment, cells were harvested to extract total RNA and protein.
Cell viability and apoptosis assays
Cell viability was carried out by cell counting kit-8 (CCK-8) assay in GMECs. After ELF5 gene overexpression or interference for 24, 48, and 72 h, the cells were incubated with CCK-8 reagent (5 g/L, CA1210; Solarbio) at 37°C for 2 h. A microplate reader (TECAN Spark) was employed to measure the absorbance of cell culture medium at 450 nm. Then, cell apoptosis was carried out by flow cytometry. The cells were transfected with pcDNA3.1-ELF5 or si-ELF5 for 48 h, subsequently cell apoptosis was determined by using the Annexin V-FITC/PI dual staining cell apoptosis detection kit (KGA1102-100; KeyGen) by a flow cytometer (BD Accuri C6puls).
RNA extraction and quantitative real-time polymerase chain reaction
Total RNA of GMECs was extracted by SPARKeasy (AC0-202-B; Sparkjade). The quality of RNA was evaluated by employing a NanoDrop One spectrophotometer (Thermo Fisher Scientific), where the ratio of absorbance at 260 nm to 280 nm ranged from 1.8 to 2.1. The integrity of RNA was determined through the analysis of 28S and 18S rRNA via agarose gel electrophoresis, with a ratio of 28S/18S approximately 2:1. For the quantitative analysis of cell proliferation and apoptosis marker genes as well as casein genes, the first strand cDNA was generated by utilizing purified total RNA through the PrimeScriptRT reagent kit (PC7001; Aidlab), which incorporates a gDNA eraser. The synergetic binding reagent green kit (PC3301; Aidlab) was used for quantitative real-time polymerase chain reaction (qPCR). Employing ubiquitous expression transcript (UXT) as an internal control gene, the qPCR primer sequences are presented in Table 1. All qPCR reactions were conducted on the CFX96 sequence detection system (Bio-Rad), and the data were analyzed by relative quantification (2^−ΔΔCt^) method.
Western blot
Total protein of GMECs was harvested after treated with radio immunoprecipitation assay lysis buffer (R0010; Solarbio) containing protease inhibitors (G2006-250UL; Servicebio). The protein assay kit (P0010S; Beyotime) was employed to detect protein concentration. Proteins were separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membrane (IPVHO0005; Merck) by BG-transBLOT. The membrane was blocked with 5% skim milk for 2 h (D8340; Solarbio) and subjected to incubation with the primary antibody at 4°C overnight. Then the membrane incubated with the secondary antibody. Primary antibodies contains ELF5 (sc-166653, SantaCruz; 1:1,000), αS2-casein (bs-10034R, Bioss; 1:2,000), κ-casein (bs-10031R; Bioss; 1:1,000), αS1-casein (bs-10033R; Bioss; 1:1,000), β-casein (sc-166530; SantaCruz; 1:1,000), p-JAK2 (ab32101; Abcam; 1:1,000), p-STAT5 (9351S; 1:1,000), JAK2 (3230; CST; 1:1,000), STAT5 (610191; BD; 1:1,000), proliferating cell nuclear antigen (PCNA, 10205-2-APl Proteintech, 1:5,000), cyclin dependent kinase 2 (CDK2, 10122-1-AP; Proteintech, 1:10,000), B-cell lymphoma-2-associated X (BAX, 50599-2-lg; Proteintech, 1:10,000), Caspase3 (19677-1-AP; Proteintech, 1:2,000), β-Tubulin (CW0098M; CWBIO, 1:5,000). Secondary antibody contains horseradish peroxidase-conjugated goat anti-rabbit-IgG (CW0102; CWBIO; 1:5,000) and goat anti-mouse-IgG (CW0103; CWBIO; 1:5,000). Signals were measured using an enhanced chemiluminescent Western blot system (AllianceQ9Advanced). The intensity of indicated bands was quantified by densitometry using ImageJ software (http://imagej.nih.gov/ij/). The relative abundance of proteins is normalized to β-Tubulin. The relative abundance of p-JAK2 and p-STAT5 is normalized to total JAK2 or STAT5.
Co-immunoprecipitation assay
Following 48 h co-transfection with pcDNA3.1-ELF5-MYC and PCMV-STAT5-3xFLAG vectors, GMECs were lysed for 20 min by immunoprecipitation lysis buffer. Subsequently, the mixture was centrifuged at 13,000×g for 10 min, and the supernatant was taken as the total protein. The Protein G beads were washed with pre-cooled phosphate buffer solution and diluted to a 50% suspension. Subsequently, Protein G beads (L-1202A; Biolinkedin) pre-washing was employed to eliminate non-specific adsorption from the cell lysate. Then centrifuged total cell protein and Protein G beads suspension were mixed and incubated at 4°C for 1 h. Centrifugation was performed at 600×g for 3 min, and the supernatant was collected. The antibody and Protein G beads were pre-incubated. The diluted solution was added Mouse-MYC-tag antibody (60003-2-Ig; Proteintech), Mouse-FLAG-tag antibody (66008-4-Ig; Proteintech) and Protein G beads suspension. The mixtures were incubated at 4°C for 1 h and centrifuged at 600×g for 3 min to retain the precipitates for three times. Subsequently, the samples were collected for Western blot detection.
Statistical analysis
The data were conducted by SPSS 22.0 (IBM) and presented as mean±standard error of the mean (SEM) of three biological replicates. When only two groups were compared, the data were analyzed through the t-test (two-tailed). For multiple comparisons, a general linear model program was utilized for ANOVA with Tukey’s test. A difference was regarded as statistically significant (* p<0.05, ** p<0.01).
RESULTS
Isolation and identification of goat mammary epithelial cells
Primary GMECs were migrating out from the goat mammary tissue, and purified cells were attained after 5 passages of purified culture (Figure 1A). Oil red O staining showed that numerous lipid droplet milk globules were dispersed around the GMECs, signifying that cells possess normal secretory function (Figure 1B). Immunofluorescence assays showed mammary epithelial cell markers cytokeratin 18 and β-casein were positive staining (Figure 1C).
Overexpression and interference of ELF5 gene
To explore the regulatory mechanism of ELF5 on casein synthesis, overexpression vector and siRNA of ELF5 gene were performed. qPCR and Western blot results indicated that pcDNA3.1-ELF5 vector could significantly enhance the mRNA and protein expression of ELF5 compared with the pcDNA3.1-NC group (p<0.001, Figures 2A, 2B, Supplement 1). Similarly, compared with the siRNA-NC group, siRNA-ELF5 markedly decreased the mRNA (p<0.001) and protein expression (p = 0.005) of ELF5 (Figures 2C, 2D, Supplement 2).
Cell proliferation and apoptosis after ELF5 gene overexpression
The CCK-8 results showed that cell viability was significantly enhanced after ELF5 gene overexpression for 24, 48, and 72 h (p<0.01, Figure 3A). Flow cytometry assay revealed cell apoptosis rate was decreased by overexpressing ELF5 gene (p = 0.043, Figure 3B). The results of qPCR and Western blot showed that ELF5 gene overexpression markedly increased the mRNA and protein expression of proliferation marker genes PCNA and CDK2, and conspicuously decreased the expression of apoptosis marker genes Bax and Caspase3 (p< 0.05, Figures 3C, 3D, Supplement 3).
Cell proliferation and apoptosis after ELF5 gene interference
The CCK-8 results showed that cell viability was significantly decreased after ELF5 gene interference for 24, 48, and 72 h (p<0.01, Figure 4A). Flow cytometry assay revealed cell apoptosis rate was increased by knocking down ELF5 gene (p = 0.035, Figure 4B). qPCR and Western blot results showed that ELF5 gene interference markedly decreased the mRNA and protein expression of PCNA and CDK2, and increased the expression of Bax and Caspase3 (p<0.05, Figures 4C, 4D, Supplement 4).
Casein synthesis by ELF5 gene overexpression and interference
In comparison with control group, ELF5 gene overexpression conspicuously elevated the mRNA and protein expression of αS1-casein, αS2-casein, β-casein, and κ-casein as well as the activity of phosphorylated JAK2 and STAT5 (p<0.05, Figures 5A, 5B, Supplement 5). In addition, ELF5 gene interference significantly decreased the expression of αS1-, αS2-, β-, and κ-casein, and reduced the activity of JAK2/STAT5 signaling pathway (p<0.05, Figures 5C, 5D; Supplement 6).
Casein synthesis by ELF5 mediated JAK2/STAT5 signaling pathway
Regulation of casein synthesis by JAK2/STAT5 signaling pathway was confirmed by Western blot assay. The protein abundance of p-JAK2, p-STAT5, αS1-casein, αS2-casein, β-casein, and κ-casein were reduced after JAK2 inhibitor AG490 treatment (p<0.01, Figure 6A; Supplement 7). Then, the protein abundance of p-STAT5 and four caseins were significantly reduced in STAT5 inhibitor STAT5-IN-1 group (p<0.01, Figure 6B, Supplement 8).
Moreover, to validate whether ELF5 promotes casein synthesis via JAK2/STAT5 signaling pathway, the rescue experiments were carried out. Overexpression of ELF5 gene upregulated p-STAT5 activity and casein expression, while co-treatment with STAT5 inhibitor abrogated the increase of p-STAT5 activity and casein expression (Figure 7A, Supplement 9). Simultaneously, knockdown of ELF5 gene downregulated p-STAT5 activity and casein expression, whereas co-treatment with STAT5 inhibitor further reduced activity of p-STAT5 and abundance of caseins (Figure 7B, Supplement 10). Furthermore, co-treatment with ELF5 gene overexpression and JAK2 inhibitor abrogated the increase of JAK2/STAT5 activity and casein expression (Figure 8A, Supplement 11). Besides, co-treatment with ELF5 gene interference and JAK2 inhibitor further decreased JAK2/STAT5 activity and casein expression (Figure 8B, Supplement 12).
Immunoprecipitation of ELF5 and STAT5
To verify whether ELF5 interacts with STAT5, an immunoprecipitation assay was carried out. We demonstrated that ELF5 interacted with STAT5 in GMECs by Western blot assay (Figure 9A). Moreover, qPCR result showed that the mRNA level of ELF5 was highly elevated after STAT5a gene overexpression (p<0.001, Figure 9B, Supplement 13). These results further validate that ELF5 could regulate casein synthesis via JAK2/STAT5 signaling pathway by interacting with STAT5 (Figure 10).
DISCUSSION
ELF5 plays crucial roles in cell proliferation, differentiation, development, cell apoptosis and milk synthesis of mammary gland [12]. In human mammary epithelial cell lines, prolactin upregulated the expression of ELF5, and milk protein synthesis was blocked in the absence of prolactin, while overexpression of ELF5 restored milk protein synthesis [13]. In the ELF5 gene knocked out mouse model, there was a notable stagnation of mammary development, encompassing a decrease in milk protein synthesis [14]. ELF5 gene overexpression could increase the milk protein content in an inducible transgenic model [15]. In lactating dairy goats, ELF5 binds to the lactalbumin gene promoter sequence to promote the synthesis of α-lactalbumin and β-lactoglobulin [16]. In our previous study, ELF5 and CSN2 gene exhibit higher expression levels during the peak and mid-lactation periods compared to dry lactation period of goats [17]. Our study indicated that ELF5 promotes the synthesis of caseins in GMECs by overexpression and interference of ELF5 gene. We investigated the mechanism of ELF5 on casein synthesis and refined the function of ELF5 in mammary lactation of goats.
During pregnancy, ELF5 regulates the lactation process in mammary gland, facilitating the differentiation of mammary stem cells into secretory cell types [18]. Furthermore, recombinant ELF5 activates the promoter of whey proteins, facilitating the development of mouse mammary epithelial cells [19]. Moreover, ELF5 regulates the proliferation and apoptosis of epithelial cells in the respiratory system, such as secretory cells and alveolar type II cells [20]. In breast cancer cells, ELF5 binds to CD24 gene promoter region, thereby regulating CD24 expression to inhibit cell proliferation and invasion [21,22]. In the present study, in addition to enhancing casein synthesis, ELF5 also promotes cell proliferation and inhibits cell apoptosis in GMECs. which predicts a tight link between casein synthesis and cell activity.
In mammals, endocrine hormones such as prolactin, insulin, and hydrocortisone maintain the lactation process and regulate milk protein synthesis through JAK2/STAT5 and mTOR pathways [23]. Phosphorylated STAT5 translocates into the nucleus, and binds to gamma-interferon activation sequence, thereby facilitating the transcription of casein gene promoter. JAK2/STAT5 regulate the expression of suppressor of cytokine signaling 3 and sterol regulatory element-binding protein implicated in casein synthesis [24,25]. Furthermore, phosphorylated STAT5 activates mTOR signaling pathway to facilitate the translation of casein genes [26]. In dairy cow mammary epithelial cells, STAT5 regulates the expression of amino acid transporters and promotes milk protein synthesis, and JAK2/STAT5 and mTOR pathways compensate to maintain milk protein synthesis [27]. In our study, inhibition of JAK2 or STAT5 activity decreased casein expression of goats, confirming the function of JAK2/STAT5 in milk protein synthesis. Furthermore, the overexpression and knockdown of ELF5 gene and the inhibition of JAK2/STAT5 signaling pathway were employed, which demonstrated that ELF5 mediates the regulation of casein synthesis via JAK2/STAT5 signaling pathway.
The expression of ELF5 in mammary epithelial cells is regulated by STAT5, while ELF5 concurrently enhanced STAT5 activity. The specific knockout of ELF5 in the mouse mammary gland reduced the phosphorylation activity of STAT5, and inhibited the synthesis and secretion of milk protein, while overexpression of ELF5 gene significantly promoted the production of milk protein [28]. Compared with the dry lactation stage of yaks, ELF5 was highly expressed in the lactation stage, which enhanced the phosphorylation activity of STAT5 and affected the expression of key genes in milk protein synthesis [29]. ELF5 binds to the STAT5 gene promoter sequence in the mammary tissue of dairy cows, enhance the transcription level and protein activity of STAT5, and regulate milk protein synthesis [30]. ELF5 and STAT5 modulate the differentiation of mammary epithelial cells to jointly govern the synthesis of milk proteins [31]. STAT5 could interact with ELF5 to augment the transcriptional activity of casein genes [32]. The co-immunoprecipitation experiment substantiates the interaction between ELF5 and STAT5, and the phosphorylation of STAT5 is up-regulated due to ELF5 gene overexpression, further intensifying the activation of JAK2/STAT5 pathway to facilitate milk production.
In dairy cow mammary epithelial cells, the proliferation, apoptosis, and casein synthesis were regulated by nutrition and hormone mediated JAK2/STAT5 and mTOR signaling pathway [33,34]. JAK2/STAT5 pathway promotes the expression of cell cycle-related genes cyclin D1 and cyclin E1, inhibiting the expression of cell cycle inhibitory factors p21 and p27, and apoptotic signaling pathway Caspase3 and Caspase8, thereby suppressing cell apoptosis and facilitating cell proliferation [20]. The vitality and quantity of mammary epithelial cells constitute the key indicator influencing milk protein secretion, and high vitality cells could effectively utilize nutrients for the synthesis of milk proteins [35]. In this study, ELF5 elevated the expression of cell proliferation markers CDK2 and PCNA, whilst inhibiting the expression of cell apoptosis markers Bax and Caspase3. Cell survival influences cellular metabolic activity, casein synthesis capacity, and the maintenance of lactation function; high cell survival facilitates casein synthesis in mammary epithelial cells [36–38]. While, the mechanism by which ELF5 mediates JAK2/STAT5 pathway to regulate cell activity still needs to be further studied.
CONCLUSION
ELF5 promotes cell viability and upregulated casein synthesis through JAK2/STAT5 signaling pathway in GMECs. ELF5 augments the activation of JAK2/STAT5 pathway by interacting with STAT5, thereby facilitating casein synthesis of goats. Our study provides a theoretical basis for increasing casein yield and improving milk protein quality in goats.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gillespie AL Green BD The bioactive effects of casein proteins on enteroendocrine cell health, proliferation and incretin hormone secretion Food Chem 20162111485910.1016/j.foodchem.2016.04.10227283618 · doi ↗ · pubmed ↗
- 2Hassanin AA Osman A Atallah OO Phylogenetic comparative analysis: chemical and biological features of caseins (alpha-S-1, alpha-S-2, beta- and kappa-) in domestic dairy animals Front Vet Sci 2022995231910.3389/fvets.2022.95231936187819 PMC 9519386 · doi ↗ · pubmed ↗
- 3de Gaudry DK Lohner S Bischoff K A 1- and A 2 beta-casein on health-related outcomes: a scoping review of animal studies Eur J Nutr 20226112110.1007/s 00394-021-02551-x PMC 878386034075432 · doi ↗ · pubmed ↗
- 4Li Z Bai R Zhang L Effect of electron beam irradiation on the structural characteristics and functional properties of goat's milk casein Int J Biol Macromol 20242601294263823288810.1016/j.ijbiomac.2024.129426 · doi ↗ · pubmed ↗
- 5Zhu J Yang L Deng H Hepatic-derived extracellular vesicles in late pregnancy promote mammary gland development by stimulating prolactin receptor-mediated JAK 2/STAT 5/m TOR signalling Int J Biol Macromol 202428113649810.1016/j.ijbiomac.2024.13649839393720 · doi ↗ · pubmed ↗
- 6Deacon AM Blouin R Thibault C Lacasse P Mechanism underlying the modulation of milk production by incomplete milking J Dairy Sci 2023106783913642431810.3168/jds.2022-22164 · doi ↗ · pubmed ↗
- 7Qi H Wang L Zhang M Wang Z Gao X Li M Methionine and leucine induce ARID 1A degradation to promote m TOR expression and milk synthesis in mammary epithelial cells J Nutr Biochem 202210110892410.1016/j.jnutbio.2021.10892434843932 · doi ↗ · pubmed ↗
- 8Kwon HC Jung HS Kim DH Han JH Han SG The role of progesterone in Elf 5 activation and milk component synthesis for cell-cultured milk production in MAC-T cells Animals 20241464210.3390/ani 1404064238396610 PMC 10886090 · doi ↗ · pubmed ↗