Silencing of the Nicotiana benthamiana phytoendesaturase gene by root treatment of exogenous dsRNA
Т.S. Golubeva, V.A. Cherenko, E.A. Filipenko, I.V. Zhirnov, A.A. Ivanov, A.V. Kochetov

TL;DR
This paper explores using exogenous dsRNA to silence a gene in plants without genetic modification, offering a potential alternative to chemical pesticides.
Contribution
The study demonstrates a non-GMO method for gene silencing in plants using exogenous dsRNA delivered through root treatment.
Findings
Exogenous dsRNA can effectively silence the phytoene desaturase gene in Nicotiana benthamiana.
Root treatment with dsRNA caused leaf bleaching, indicating successful gene knockdown.
The method avoids genetic modification and could be used for agricultural plant protection.
Abstract
RNA interference (RNAi) is a powerful tool for gene silencing. It has recently been used to design promising plant protection strategies against pests such as viruses, insects, etc. This generally requires modifying the plant genome to achieve in planta synthesis of the double-stranded RNA (dsRNA), which guides the cellular RNA interference machinery to silence the genes of interest. However, given Russian legislation, the approach in which dsRNA is synthesized by the plant itself remains unavailable for crop protection. The use of exogenously produced dsRNA appears to be a promising alternative, allowing researchers to avoid genetic modification of plants, making it possible to implement potential results in agriculture. Furthermore, exogenous dsRNAs are superior to chemical pesticides (fungicides, insecticides, etc.), which are widely used to control various plant diseases. The dsRNA…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
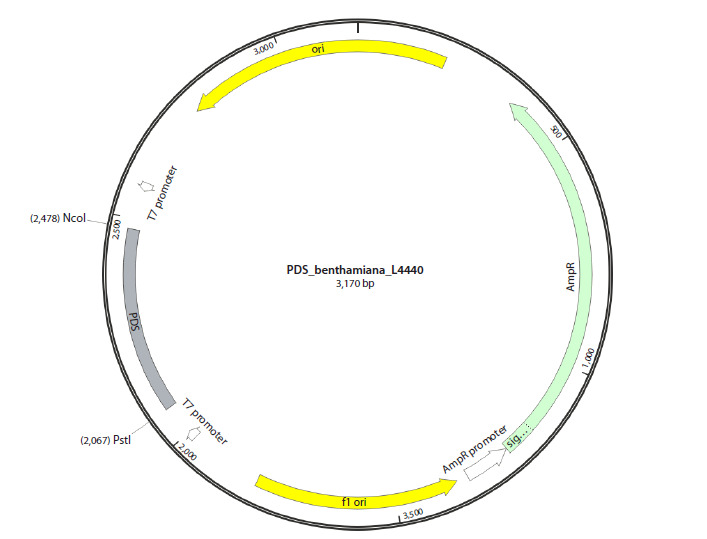 Fig. 1
Fig. 1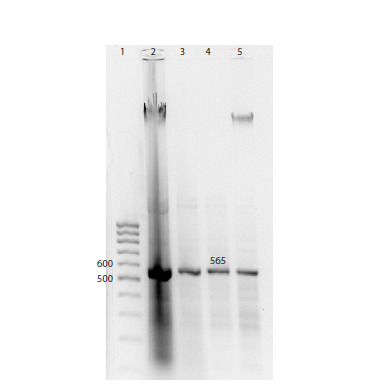 Fig. 2
Fig. 2 Fig. 3
Fig. 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Virus Research Studies · Plant Reproductive Biology · Insect Resistance and Genetics
Introduction
In order to protect plants against pathogens and pests, agriculture relies on the widespread use of chemical pesticides that are applied to the environment in large amounts. These intense applications of chemical pesticides pose potential risks for human health, beneficial organisms, and the environment (Niehl et al., 2018). Therefore, it is imperative that new alternative methods of controlling plant diseases be developed. Thus, there is a need for novel tools and alternative methods to control disease epidemics. A promising new approach with strong potential for protecting plants against viruses and other pathogens involves the application of dsRNA.
dsRNA applications can be highly advantageous over chemical compounds. Whereas chemical compounds act by a structure-dependent mechanism, dsRNAs act through their specific nucleotide sequence. Hence, once engineered to affect a specific pathogen target with a homologous sequence, dsRNAs and small interfering RNA (siRNA) derivatives should act only against the targeted pathogen. It is worth noting that, unlike chemical pesticides, dsRNA agents are biocompatible and biodegradable compounds, natural and universally found inside and outside organisms as well as in food. In this way, applying dsRNA proves to be a much more flexible and environmentally friendly approach.
However, the wide application of dsRNA treatment in agriculture is hampered by the lack of efficient and costeffective methods to synthesize large quantities of dsRNA. The main approach to obtain dsRNA has been the physical annealing of two enzymatically synthesized single-stranded RNAs (ssRNA) in vitro (Laurila et al., 2002). Also, the efficiency of exogenously administered dsRNA in plants can be influenced by several factors: dsRNA concentration/dose and length/size, method of application, method of delivery, plant organ-specific activity, and stability under unfavorable environmental conditions. It is these factors that eventually determine the rate of uptake of exogenous dsRNA by plant cells for RNAi triggering. Given all these factors, it is necessary to develop an efficient method for the large-scale synthesis of dsRNA molecules and to choose the best method for their delivery (Carthew, Sontheimer, 2009).
In this work, we suggest a method for regulating the activity of the phytoene desaturase gene in tobacco (Nicotiana benthamiana) using dsRNA synthesized in Escherichia coli cells: root treatment of plants with crude bacterial lysate containing the target dsRNA led to photobleaching of plant leaves by RNA interference (RNAi).
Phytoene desaturase is a key enzyme in chlorophyll synthesis, with its silencing known to result in the phenotypic manifestation of leaf photobleaching. Thus, this gene is often used as a model for developing new approaches to regulate gene activity. Root treatment with crude lysate allows scaling up dsRNA production and minimizing delivery time and resources. The approach developed is expected to be used to protect agricultural plants by regulating gene activity by RNAi.
Materials and methods
Characterization of the bacterial strain. This study used the E. coli HT115 (DE3) strain [F-, mcrA, mcrB, IN(rrnD-rrnE)1, rnc14::Tn10(DE3 lysogen: lacUV5 promoter – T7 polymerase) (IPTG-inducible T7 polymerase) (RNAse III minus)]. This strain is deficient in RNase III and therefore can be used to generate dsRNA. The strain was cultured in standard Luria-Bertani (LB) liquid media or LB agar with tetracycline (12.5 μg/mL).
Characterization of the L4440 plasmid. Plasmid L4440 (Plasmid #1654, Addgene, USA) was used in this study. The specific feature of this plasmid (i. e. the presence of two strong T7 promoters in opposite directions) makes it suitable for the production of dsRNA fragments. Also, the plasmid carries an ampicillin resistance gene.
Selection of primers. Primers were selected using the Primer-BLAST resource (https://www.ncbi.nlm.nih.gov/tools/ primer-blast/index.cgi) on the N. benthamiana mRNA matrix. The selected primer variants were tested for the presence of hairpins, self- and heterodimers using the OligoAnalyzer tool (https://eu.idtdna.com/calc/analyzer). As a result, the following primers were synthesized: forward 5′-GGCACTC AACTTTATAAACC-3′ and reverse 5′-CTTCAGTTTTCT GTCAAACCATATATGGAC-3′ (Syntol, Russia).
Production of cDNA. mRNA was isolated using the RNeasy Plant kit (Qiagen, USA) according to the manufacturer’s protocol, with the RNA sample obtained analyzed on a Bioanalyzer Instrument 2100 RNA analyzer (SB RAS Genomics Core Facility, ICBFM SB RAS). Reverse transcription was performed with the BiolabMix R01-250 kit (BiolabMix, Russia). The HS-qPCR SYBR Blue kit (BiolabMix, Russia) was used to generate fragments, and amplification was performed in a Bio-Rad IQ instrument (Bio-Rad, USA).
Cloning of the pds gene fragment. The pds gene was cloned in two steps. Initially, the gene was cloned into the pCR2.1 intermediate T-vector (Invitrogen, USA). Bacterial colonies containing the gene fragment were first selected using the blue-white screening. The final selection of clones was carried out based on the presence of the complete gene fragment in the restriction spectra after treatment with EcoRI endonuclease.
Cloning into the final vector L4440 was performed using the ligase-restriction method with the restriction endonucleases PstI and NcoI. For this purpose, the corresponding restriction sites were introduced into the pds gene fragment during its amplification (the same sites are present in the structure of the L4440 vector). Recombinant E. coli clones were selected on a selective medium containing ampicillin. To verify the presence of the pds gene fragment in the L4440 vector, PCR was performed using primers specific for the pds gene. The final presence of the pds gene fragment and its nucleotide sequence were confirmed by Sanger sequencing.
Transformation of the E. coli strain HT115 (DE3) with plasmid L4440. Competent cells were prepared according to the Nishimura protocol (Nishimura et al., 1990). For transformation, a 0.1 ml aliquot of bacterial suspension was mixed with 5 μl of L4440 plasmid dissolved in TE buffer (100 pg) and incubated on ice for 30 min. Next, the cells were placed in an incubator and maintained at 42 °C for 60 s, then incubated again on ice for 2 min and diluted 10-fold with pre-warmed to 37 °C LB broth. The suspension was incubated at 37 °C for 1 h to develop ampicillin resistance. Then 100 μl of the suspension was rubbed into LB agar with ampicillin (50 μg/ ml), and the cups were incubated at 37 °C for 24 h. The grown colonies were formed by transformed bacteria.
The presence of an integrated fragment of the pds gene was verified by PCR of selected colonies as well as by Sanger sequencing (SB RAS Genomics Core Facility, ICBFM SB RAS).
VIGS positive control. A model plant, N. benthamiana, was used in VIGS experiments. Seeds of N. benthamiana were germinated and then the seedlings were planted in plastic pots (10 cm in diameter) containing a mixture of universal potting soil (TerraVita, Russia), perlite and vermiculite (8:1:1, v/v), and cultivated in a growth chamber under continuous lighting at 24 °C.
Vector construction. The VIGS vectors pTRV1 (pYL192) and pTRV2 (pYL279) obtained from the Arabidopsis Biological Resource Center (ABRC, USA) were described previously (Liu et al., 2002; Burch-Smith et al., 2006). Total RNA was extracted from leaf tissues of Nicotiana tabacum cv. SR1 (the phytoendesaturase gene sequence in this species is identical to that in N. benthamiana) using Trizol reagent (Invitrogen, USA), and first-strand cDNA was synthesized using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, USA) according to the manufacturer’s instructions. The cDNA pool was used to amplify the NtPDS gene fragment sequence (GenBank accession number: AJ616742.1) by PCR using high-fidelity Phusion polymerase (New England Biolabs, USA) as per the manufacturer’s instruction. The selected primers were checked for hairpins, self-dimers, and heterodimers using the OligoAnalyzer tool. As a result, the following primers were synthesized: forward 5′-CACCGGCACTC AACTTTATAAACC-3′ and reverse 5′-CTTCAGTTTTCT GTCAAACCATATATGGAC-3′ (Syntol, Russia).
The resulting 413-bp PCR fragment was cloned into the pENTR/D-TOPO vector (Invitrogen, USA), verified by sequencing, and then recombined into the pTRV2 vector by carrying out an LR recombination reaction using the Gateway system (Invitrogen). The generated vector pTRV2::NtPDS was transformed by heat shock into the Agrobacterium tumefaciens GV2260 strain and used in VIGS experiments
The A. tumefaciens GV2260 strain, carrying the pTRV1, pTRV2, and pTRV2::NtPDS vectors, was separately inoculated into Luria-Bertani liquid media containing kanamycin (100 μg/mL) and rifampicin (25 μg/mL). The cultures were incubated overnight with shaking at 28 °C. The cells were harvested from the overnight cultures, re-suspended in the induction buffer (10 mM MES; 10 mM MgCl2; 250 μM acetosyringone; adjusted to pH 5.5 with 1 M KOH), and incubated for 6 h at room temperature in a shaker. After incubation, the cells were harvested from the induced cultures and resuspended in the infiltration buffer (10 mM MES, adjusted to pH 5.5 with 1 M KOH) with dilution to a final absorbance OD600 = 1.0
For leaf infiltration, A. tumefaciens strain GV2260 cultures containing pTRV1 and pTRV2 or pTRV2::NtPDS were mixed in a 1:1 (v/v) ratio and infiltrated into lower leaves of 21‐days‐old N. benthamiana plants using a 1-ml needleless syringe (Ratcliff et al., 2001; Liu et al., 2002). Infiltrated plants were maintained under constant light for 12 h at a 20 °C temperature for effective Agrobacterium T-DNA insertion (Brigneti et al., 2004).
Root treatment of N. benthamiana with a crude lysate of E. coli HT115 (DE3) containing dsRNA of phytoendesaturase. After inducing dsRNA synthesis, crude lysates of bacterial suspensions can be used as a source of exogenous dsRNA. Root treatments are also used to deliver target molecules for PTGS in plants (Jiang et al., 2014; Li et al., 2015; Dubrovina, Kiselev, 2019).
Crude lysates of the E. coli HT115 strain were prepared for root treatment, transformed with the L4440 plasmid with an integrated fragment of the pds gene. Bacterial lysates without plasmid, with incorporation of the native plasmid L4440, buffer, and water were used as controls.
The crude lysate was prepared in the following manner (Gan et al., 2010). An overnight culture of the bacteria was grown for 16 h with shaking at 37 °C in standard LB broth, with ampicillin at a concentration of 50 μg/mL added for strains with the L4440 plasmid insertion. The bacterial suspension was then diluted to OD595 = 0.5. Next, dsRNA synthesis was induced by adding IPTG at a final concentration of 0.6 mM, and the bacteria were incubated at 37 °C for 4 h. After the required time, the suspension was centrifuged to obtain a precipitate (1,500g, 15 min, 4 °C), which was then resuspended in ice buffer (50 mM Tris·HCl, 10 mM EDTA, pH 7.5) taken in 1/50 of the original volume. The tubes were then placed in ice and treated with ultrasound (20 kHz, 15 min). The resul ting lysate was centrifuged (9,000 rpm, 20 min, 4 °C), and the supernatant was used for plant treatments (2 ml per plant).
Four plants were used for each type of root treatment: water, Tris·HCl/EDTA buffer, E. coli HT115 without plasmid, E. coli HT115 with the L4440 plasmid, E. coli HT115 with the L4440 plasmid with an integrated pds fragment. All plants were grown in a sterile mixture of universal soil (TerraVita, Russia), perlite and vermiculite (8:1:1, v/v). The plants were treated three times a week (Monday, Wednesday, Friday) for a period of four weeks. Root treatment was carried out by watering the plants with supernatant resuspended in 2 ml of Tris-EDTA buffer
Extraction of dsRNA. 1 ml of the bacterial suspension was centrifuged for 5 min at 10,000 rcf, and the supernatant was removed. The precipitate was dissolved in 200 μl of a mixture of 1 M CH3COONH4 and 10 mM isoamyl alcohol and extracted with an equivalent volume of isopropanol:phenol:isoamyl alcohol in the ratio (25:24:1). Then it was incubated for 15 min at 65 °C and centrifuged for 15 min at 10,000 rcf. The supernatant was withdrawn and an equivalent volume of isopropanol was added, followed by incubation for 12 hours at –20 °C. After incubation, it was centrifuged for 30 min at 12,000 rcf. The liquid was removed carefully. The precipitate was washed twice with 70 % ethanol, with the precipitate resuspended in 10 μl of RNase-free water, and DNAase buffer was added. Next, the incubation was done for 30 min, followed by the addition of 20 μl of RNase A and 20 μl of DNase. Then, the incubation was performed for 1 hour at 37 °C. 100 μl of a isopropanol:phenol:isoamyl alcohol mixture in the ratio (25:24:1) was extracted. The mixture was stirred vigorously and centrifuged at 12,000 rcf for 15 min. The supernatant was removed, and the precipitate was washed with 200 μl of 70 % ethanol, dried at room temperature, and dissolved with 1× TE buffer. The presence and integrity of dsRNA were checked by gel electrophoresis in 1 % agarose gel.
Results
Production of exogenous dsRNA using an RNase-deficient E. coli HT115 strain transformed with the L4440 plasmid
The E. coli HT115 (DE3) strain was used to produce dsRNA because it is deficient in RNase III, an enzyme that normally hydrolyzes dsRNA in the bacterial cell. The L4440 vector was used for the transformation. Due to having two strong T7 promoters in opposite directions, this vector can be used to produce target dsRNA fragments.
A 404 bp fragment of the tobacco phytoene desaturase pds gene was cloned into the L4440 vector by sticky ends using restriction endonucleases PstI and NcoI, with the presence of the required fragment in the vector confirmed by sequencing. A map of the resulting plasmid is shown in Figure 1.
Map of vector L4440 containing a fragment of the pds gene of N. benthamiana (PDS_benthamiana_L4440).
The presence of the target fragments of the correct length was confirmed by electrophoretic analysis (Fig. 2).
Electrophoretic analysis of dsRNA fragments synthesized in E. coli HT115 (DE3), transformed with the PDS_benthamiana_L4440 vector.1 – 100 bp DNA marker; 2 – total nucleic acid fraction; 3 – total nucleic acid fraction treated with DNase; 4 – total nucleic acid fraction treated with RNase A and DNase; 5 – total nucleic acid fraction treated with RNase A.
Preparation of a crude bacterial lysate containing dsRNA fragments for silencing phytoene desaturase N. benthamiana
A strain in E. coli HT115 (DE3) transfected with the PDS_benthamiana_ L4440 vector was used to produce target dsRNA fragments for silencing pds genes. Induction of dsRNA synthesis was performed using IPTG. Next, a crude lysate was obtained from the bacterial suspension (Gan et al., 2010) containing dsRNA. 2-ml aliquots of the crude lysate were prepared for treatment of N. benthamiana plants.
Root treatment of N. benthamiana with a crude bacterial lysate containing dsRNA for silencing the pds gene
In this study, root treatment of tobacco with a crude bacterial lysate containing exogenous dsRNA was performed for the first time. Bentham tobacco (N. benthamiana) was used as a model plant for the experiment. Experimental plants were treated with the E. coli HT115 (DE3) crude lysate with the PDS_benthamiana_L4440 plasmid. After four weeks of root treatment, N. benthamiana leaves treated with the bacterial lysate with insertion of a phytoene desaturase gene fragment showed photobleaching phenotypes of young leaves typical for pds gene silencing (Fig. 3). The leaves of N. benthamiana plants from the negative control did not change their phenotype throughout the entire treatment period.
Photobleaching of N. benthamiana leaves after induced silencing of the pds gene using exogenous dsRNA.1 – water; 2 – Tris-EDTA; 3 – E. coli HT115 crude lysate without plasmid; 4 – E. coli HT115 crude lysate with the native L4440 plasmid; 5 – VIGS; 6 – crude lysate of E. coli HT115 with the L4440 plasmid containing a fragment of the pds gene; 7 – comparison of a negative control with an experimental plant with pds silencing. Controls in the experiment: treatments 1–4 serve as negative controls, treatment 5 serves as a positive control. Characteristics of leaf photobleaching: L – lightness of color in relative units (the higher the value, the lighter the shade, from 0 to 100); a – green-red channel in relative units (the lower the value, the greener the shade, from –127 to 0).
Discussion
Currently, there are numerous studies indicating the possibility of turning off or reducing the expression of certain genes to regulate resistance, growth, and other properties of plants by inducing RNA interference (Kamthan et al., 2015; Tiwari et al., 2017). Yet this approach involves the stage of obtaining a transgenic plant or using constructs based on attenuated plant viruses.
The problem of RNA delivery without using vector systems and modifying the genome became particularly acute after GMOs had been banned in Russia and European countries. Recently, reports have appeared in the scientific literature indicating that exogenously applied double-stranded RNA molecules (e. g. by spraying, spraying under high pressure, using RNA molecule adhesion materials, or using transfer proteins) are capable of penetrating into the plant vascular system and directly into plant cells and then inducing RNA interference, thereby increasing plant resistance to fungal and viral infections (Numata et al., 2014; Koch et al., 2016; Mitter et al., 2017; Wang et al., 2018). There are also few studies on dsRNA delivery aimed at regulating gene function by irrigation. For example, maize resistance to sugarcane mosaic virus (SCMV) infection was enhanced by this delivery method (Gan et al., 2010).
Similarly, we obtained a crude lysate containing a dsRNA fragment of the N. benthamiana phytoene desaturase gene, and carried out root treatment of tobacco for four weeks. As a result, all experimental plants exhibited the phenotype of photobleaching of young leaves, which is not uncommon for phytoene desaturase silencing. The degree of whitening was comparable to the positive VIGS control. Thus, we suggest the approach that allows regulating the activity of plant genes without creating GMOs and using only environmentally friendly methods. We believe this approach to be highly promising for implementation in agriculture in order to improve the stress tolerance of cultivated plants.
Our work is based on research by F. Tenllado and coauthors (2003), where tobacco (N. benthamiana) was successfully protected from pepper mild mottle virus (PMMoV) by spraying the above-ground parts of the plants with a crude lysate of E. coli HT115 bacteria containing dsRNA. The authors note that this method of obtaining target dsRNA molecules is quite simple and economically advantageous compared to in vitro dsRNA synthesis. The authors also show that the use of crude lysate may be more cost-effective than the use of a purified preparation, with no significant loss of efficacy observed.
Since plant protection using RNAi methods can be an alternative to the creation of GMOs, which are prohibited by law in the Russian Federation for agricultural purposes, we decided to develop the idea of F. Tenllado and co-authors (2003) and try an even simpler method of delivering dsRNA through root treatment by watering plants with crude lysate. We have shown that the introduction of exogenously synthesized dsRNA in this way can effectively influence the phenotype of plants, confirming that the introduced dsRNA enters the plants through the roots and is transported to the above-ground part of the plant, where the activity of the target pds gene is regulated.
Conclusion
Recently, methods based on RNAi have been actively developed for plant protection, especially with the use of exogenously synthesised dsRNA, as they allow avoiding the creation of GMOs, which are prohibited by law for use in agriculture in a number of countries, including Russia. However, the use of exogenous dsRNA molecules has a number of complications. For example, the method of obtaining dsRNA must be easily scalable and cost-effective, and the delivery of the resulting molecules must be as simple and efficient as possible. In our work on dsRNA synthesis, we developed a system from the E. coli HT115 strain, transformed with the L4440 plasmid with a fragment of the Bentham tobacco phytoendesaturase gene (N. benthamiana), and for the delivery of the resulting molecules, we used root treatment – watering the plants with a coarse lysate. As a result, we were able to achieve silencing of the phytoendesaturase gene,which confirms the possibility of regulating the work of plant genes without creating GMOs. The approach we propose can also be scaled up and has potential for application in agriculture to protect plants from pathogens
Conflict of interest
The authors declare no conflict of interest.
References
Brigneti G., Martín-Hernández A.M., Jin H., Chen J., Baulcombe D.C., Baker B., Jones J.D.G. Virus-induced gene silencing in Solanum species. Plant J. 2004;39(2):264-272. doi 10.1111/J.1365-313X. 2004.02122.x
Burch-Smith T.M., Schiff M., Liu Y., Dinesh-Kumar S.P. Efficient virus-induced gene silencing in Arabidopsis. Plant Physiol. 2006; 142(1):21-27. doi 10.1104/pp.106.084624
Carthew R.W., Sontheimer E.J. Origins and mechanisms of miRNAs and siRNAs. Cell. 2009;136(4):642-655. doi 10.1016/j.cell.2009. 01.035
Dubrovina A.S., Kiselev K.V. Exogenous RNAs for gene regulation and plant resistance. Int J Mol Sci. 2019;20(9):2282. doi 10.3390/ ijms20092282
Gan D., Zhang J., Jiang H., Jiang T., Zhu S., Cheng B. Bacterially expressed dsRNA protects maize against SCMV infection. Plant Cell Rep. 2010;29(11):1261-1268. doi 10.1007/s00299-010-0911-z
Jiang L., Ding L., He B., Shen J., Xu Z., Yin M., Zhang X. Systemic gene silencing in plants triggered by fluorescent nanoparticle delivered double-stranded RNA. Nanoscale. 2014;6(17):9965-9969. doi 10.1039/c4nr03481c
Kamthan A., Chaudhuri A., Kamthan M., Datta A. Small RNAs in plants: recent development and application for crop improvement. Front Plant Sci. 2015;6(APR). doi 10.3389/FPLS.2015.00208/PDF
Koch A., Biedenkopf D., Furch A., Weber L., Rossbach O., Abdellatef E., Linicus L., Johannsmeier J., Jelonek L., Goesmann A., Cardoza V., McMillan J., Mentzel T., Kogel K.H. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016;12(10):e1005901. doi 10.1371/journal.ppat.1005901
Laurila M.R.L., Makeyev E.V., Bamford D.H. Bacteriophage φ6 RNAdependent RNA polymerase. Molecular details of initiating nucleic acid synthesis without primer. J Biol Chem. 2002;277(19):17117- 17124. doi 10.1074/jbc.M111220200
Li H., Guan R., Guo H., Miao X. New insights into an RNAi approach for plant defence against piercing-sucking and stem-borer insect pests. Plant Cell Environ. 2015;38(11):2277-2285. doi 10.1111/ pce.12546
Liu Y., Schiff M., Marathe R., Dinesh-Kumar S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002;30(4):415-429. doi 10.1046/J.1365-313x.2002.01297.x
Mitter N., Worrall E.A., Robinson K.E., Li P., Jain R.G., Taochy C., Fletcher S.J., Carroll B.J., Lu G.Q., Xu Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat Plants. 2017;3:16207. doi 10.1038/nplants.2016.207
Niehl A., Soininen M., Poranen M.M., Heinlein M. Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol J. 2018;16(9):1679-1687. doi 10.1111/pbi.12904
Nishimura A., Morita M., Nishimura Y., Sugino Y. A rapid and highly efficient method for preparation of competent Escherichia coli cells. Nucleic Acids Res. 1990;18(20):6169. doi 10.1093/nar/18.20.6169
Numata K., Ohtani M., Yoshizumi T., Demura T., Kodama Y. Local gene silencing in plants via synthetic dsRNA and carrier peptide. Plant Biotechnol J. 2014;12(8):1027-1034. doi 10.1111/pbi.12208
Ratcliff F., Martin-Hernandez A.M., Baulcombe D.C. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001;25(2):237-245. doi 10.1046/J.0960-7412.2000.00942.x
Tenllado F., Martínez-García B., Vargas M., Díaz-Ruíz J.R. Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC Biotechnol. 2003;3:3. doi 10.1186/ 1472-6750-3-3
Tiwari I.M., Jesuraj A., Kamboj R., Devanna B.N., Botella J.R., Sharma T.R. Host Delivered RNAi, an efficient approach to increase rice resistance to sheath blight pathogen (Rhizoctonia solani). Sci Rep. 2017;7(1):1-14. doi 10.1038/S41598-017-07749
Wang J., Gu L., Knipple D.C. Evaluation of some potential target genes and methods for RNAi-mediated pest control of the corn earworm Helicoverpa zea. Pestic Biochem Physiol. 2018;149:67-72. doi 10.1016/j.pestbp.2018.05.012