Multiplexed CRISPR/Cas9 Editing of Tumor Suppressor Genes in the Mouse Endometrium Recapitulates High-Risk Endometrial Carcinoma
Maria Vidal-Sabanés, Raúl Navaridas, Núria Bonifaci, Ada Gay-Rua, Damià Ortega-Peinado, Joaquim Egea, Mario Encinas, Xavier Matias-Guiu, David Llobet-Navas, Xavier Dolcet

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
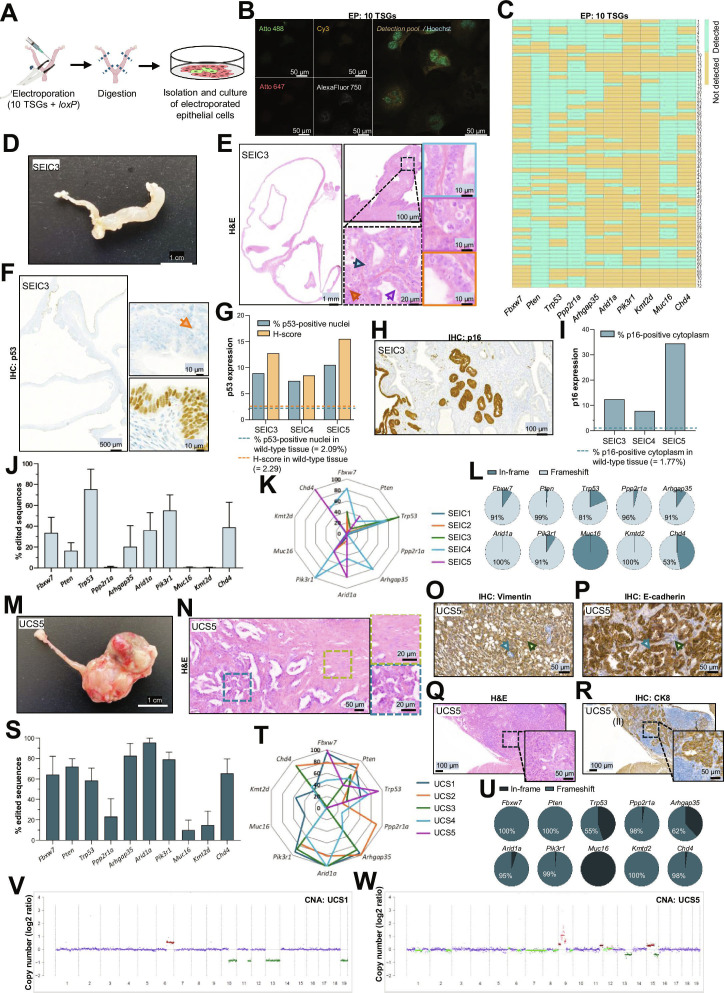 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · CRISPR and Genetic Engineering · Axon Guidance and Neuronal Signaling
Endometrial carcinoma (EC) is the fourth most common cancer in women in the Western world and the most frequent malignancy of the female genital tract, with an incidence of 10 to 20 per 100,000 women annually [1]. Traditional classifications distinguish Type I endometrioid EC from Type II non-endometrioid EC [2], while histological schemes categorize tumors into endometrioid endometrial carcinoma, clear cell carcinoma (CCC), serous endometrial carcinoma (SEC), and uterine carcinosarcoma (UCS). More recently, the Cancer Genome Atlas proposed a molecular classification defining ultramutated with polymerase ε mutations, hypermutated with microsatellite instability, copy-number low, and copy-number high (CNH) serous-like subtypes [3]. High-risk tumors, particularly UCS and SEC, are strongly associated with tumor protein 53 (TP53) mutations, extensive genomic alterations, poor prognosis, and disproportionately high mortality rates [4]. Despite these insights, the contribution of individual and combined tumor suppressor gene (TSG) mutations to EC progression remains incompletely understood, and faithful animal models of aggressive EC subtypes are scarce.
To address this, we applied an in vivo multiplex clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 strategy [5] targeting the 10 most frequently mutated TSGs in high-risk EC. Detailed materials and methods are provided in the Supplementary Materials. For gene selection, we analyzed genomic data from 1,133 EC cases across 8 studies in cBioPortal. After filtering for mutation profiles, histological grade, and survival data, we identified 334 high-risk cases (grade 3 endometrioid carcinoma, SEC, CCC, and UCS) and 479 low-risk endometrioid cases (grades 1 and 2). Comparative analysis confirmed poorer survival in the high-risk cohort and guided the selection of 10 recurrently mutated TSGs for functional modeling: TP53, protein phosphatase 2 scaffold subunit alpha (PPP2R1A), F-box and WD repeat domain containing 7 (FBXW7), chromodomain helicase DNA binding protein 4 (CHD4), rho GTPase activating protein 35 (ARHGAP35), phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1), phosphatase and tensin homolog (PTEN), AT-rich interactive domain-containing protein 1a (ARID1A), mucin 16 (MUC16), and lysine methyltransferase 2D (KMT2D) (Fig. S1A).
Cas9-ribonucleoproteins (RNPs) targeting each locus were validated in vitro using synthetic DNA substrates, confirming the efficient cleavage of all targets (Fig. S1B and C). In vivo validation in mTmG reporter mice demonstrated editing capacity, as loxP-targeting RNPs induced green fluorescent protein (GFP) expression in electroporated uterine epithelium (Fig. S1D to G). Amplicon sequencing confirmed edits at all targeted genes, though at low aggregate frequencies, consistent with the small proportion of transfected epithelial cells (Fig. S1H). Histological analysis of uteri electroporated with no RNP, Cas9 alone, or sgRNA alone showed no morphological abnormalities for up to 16 weeks after intervention (Fig. S1I). Importantly, the edited endometrial epithelial cells remained viable and could be expanded in culture, enabling the quantification of editing efficiency and assessment of functional consequences (Fig. S1J to L).
By pooling Cas9-RNPs for intrauterine electroporation, we aimed to recapitulate the genetic heterogeneity underlying high-risk EC [6]. To assess heterogeneity at single-cell resolution, we adapted multiplexed rolling circle amplification (RCA) using padlock probes targeting wild-type transcripts of the 10 genes. Loss of detectable wild-type signal indicated CRISPR editing (Fig. S2A and B). Control experiments confirmed the specificity and multiplexing capability of the system, including the successful detection of PTEN loss in knockout mice (Fig. S2C and D). RCA applied to wild-type endometrial cells (Fig. S2E to K) or cells electroporated with pooled RNPs revealed striking cell-to-cell variability: while some cells retained transcripts for all genes, others showed loss of multiple TSG mRNAs (Fig. 1A to C and Fig. S2L and M). These results confirm that multiplexed editing generates a genetically mosaic tissue, more closely resembling human tumorigenesis than uniform conditional knockouts.
We electroporated 14 mice with pooled TSG RNPs and monitored them longitudinally (Fig. S3A). Ten developed visible uterine lesions, while 4 remained macroscopically normal. Histopathology identified 2 lesion types: serous endometrial intraepithelial carcinoma (SEIC) and UCS (Fig. S3B to D). SEICs displayed classical serous features [7], including macroscopic enlargement (Fig. 1D and Fig. S4A), marked nuclear atypia, epithelial stratification, papillary growth, high mitotic activity, and apoptotic figures (Fig. 1E and Fig. S4B). Immunohistochemistry (IHC) revealed abnormal p53 staining patterns, with both null and strong nuclear phenotypes (Fig. 1F and G and Fig. S4C and D), and diffuse cytoplasmic p16 expression compared to wild-type endometrium (Fig. 1H and I and Fig. S4E and F), reflecting diagnostic characteristics of human SEIC [8]. Next-Generation Amplicon Sequencing (NGAS) confirmed indels in all targeted TSGs, with TP53 edits being particularly enriched (Fig. 1J to L).
UCS lesions exhibited distinct macroscopic features and characteristic biphasic morphology with separate carcinomatous and sarcomatous components [9] (Fig. 1M and N and Fig. S5A and B). Immunostaining confirmed epithelial–mesenchymal transition: vimentin marked sarcomatous areas while E-cadherin and cytokeratin 8 (CK8) labeled epithelial regions (Fig. 1O and P and Fig. S5C to E). All UCS lesions invaded the myometrium, and notably, one case showed lymph node metastasis, confirmed by CK8 IHC (Fig. 1Q and R). NGAS revealed heterogeneous disruptions across all 10 TSGs (Fig. 1S to U). Given that UCS is often associated with CNH serous-like tumors, we evaluated genomic stability in 2 cases by shallow whole-genome sequencing. UCS1 exhibited a largely copy-number neutral profile, with a few focal deletions and gains, consistent with a genomically stable tumor (Fig. 1V and Fig. S5F). In contrast, UCS5 displayed extensive chromosomal instability, including high-level amplifications across multiple chromosomes and pronounced subclonality, despite near-diploid ploidy (Fig. 1W and Fig. S5F and G). Interestingly, UCS1 developed within 13 weeks, whereas UCS5 required nearly 1 year to progress, suggesting that prolonged evolution may promote the accumulation of chromosomal instability. These results recapitulate the heterogeneity seen in human UCS, where some tumors arise from copy number-low molecular subtypes, while others display highly unstable genomes.
Unexpectedly, some macroscopically and histologically normal uteri harbored oncogenic mutations (Fig. S6A and B). Electroporation efficacy was confirmed by GFP staining in mTmG mice, excluding failed transfection as a possible explanation (Fig. S6C). Amplicon sequencing revealed indels in up to 5 TSGs, including TP53 and PTEN, most of which produced frameshift mutations (Fig. S6D and E). These findings demonstrate that acquisition of multiple oncogenic mutations alone is insufficient to induce malignant transformation, supporting a multistep model of EC evolution. This observation aligns with human studies identifying clonal driver mutations in histologically normal endometrium [10] and emphasizes the role of permissive cellular or epigenetic contexts in tumor initiation.
Together, our findings establish that multiplexed in vivo CRISPR/Cas9 editing of high-risk TSGs faithfully reproduces the histological and molecular diversity of aggressive EC. Notably, all observed lesions were SEIC or UCS, recapitulating human high-risk EC subtypes, while no low-grade endometrioid carcinomas emerged. This highlights the specificity of the targeted TSGs in driving aggressive histology. The system’s ability to generate mosaic populations of mutant and wild-type cells within the same tissue more closely mirrors the clonal architecture of human tumors and provides a powerful tool for dissecting cooperative mutational interactions that enable clonal selection and drive tumor expansion.
Although multiple strategies exist for delivering CRISPR/Cas9, including viral vectors, lipid nanoparticles, extracellular vesicles, and physical transfection, our approach offers distinct advantages (Table S1) and provides a preclinical framework for testing these therapeutic strategies. Tumors generated through multiplex editing displayed heterogeneity in histology, mutational spectrum, and genomic stability, modeling the diversity observed clinically. The presence of oncogenic mutations in histologically normal tissue raises important questions regarding early detection and prevention, as it suggests a latent reservoir of genetically altered cells that may progress only under specific contexts.
In conclusion, multiplex in vivo CRISPR/Cas9 editing of 10 recurrently mutated TSGs faithfully recapitulated the molecular, morphological, and genomic hallmarks of high-risk EC, generating SEIC and UCS lesions. The model captures clonal heterogeneity, reveals oncogenic mutations in normal tissues, and reproduces chromosomal instability in human UCS. By linking genomic data to functional validation, it offers a rapid and versatile platform for mechanistic studies, therapeutic testing, and deeper insights into tumor evolution in aggressive gynecologic cancers.
Ethical Approval
All procedures performed in this study followed the National Institute of Health Guide for the Care and Use of Laboratory Animals and were compliant with the guidelines of the Universitat de Lleida (permit number: N. 02-02/19).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Makker V, Mac Kay H, Ray-Coquard I, Levine DA, Westin SN, Aoki D, Oaknin A. Endometrial cancer. Nat Rev Dis Primer. 2021;7(1):88.10.1038/s 41572-021-00324-8PMC 942194034887451 · doi ↗ · pubmed ↗
- 2Bokhman JV. Two pathogenetic types of endometrial carcinoma. Gynecol Oncol. 1983;15(1):10–17.6822361 10.1016/0090-8258(83)90111-7 · doi ↗ · pubmed ↗
- 3Cancer Genome Atlas Research Network, Kandoth C, Schultz N, Cherniack AD, Akbani R, Liu Y, Shen H, Robertson AG, Pashtan I, Shen R, et al. Integrated genomic characterization of endometrial carcinoma. Nature. 2013;497(7447):67–73.23636398 10.1038/nature 12113 PMC 3704730 · doi ↗ · pubmed ↗
- 4Bell DW, Ellenson LH. Molecular genetics of endometrial carcinoma. Annu Rev Pathol. 2019;14:339–367.30332563 10.1146/annurev-pathol-020117-043609 · doi ↗ · pubmed ↗
- 5Navaridas R, Vidal-Sabanés M, Ruiz-Mitjana A, Perramon-Güell A, Megino-Luque C, Llobet-Navas D, Matias-Guiu X, Egea J, Encinas M, Bardia L, et al. Transient and DNA-free in vivo CRISPR/Cas 9 genome editing for flexible modeling of endometrial carcinogenesis. Cancer Commun. 2023;43(5):620–624.10.1002/cac 2.12409 PMC 1017408836762520 · doi ↗ · pubmed ↗
- 6Navaridas R, Vidal-Sabanés M, Ruiz-Mitjana A, Altés G, Perramon-Güell A, Yeramian A, Egea J, Encinas M, Gatius S, Matias-Guiu X, et al. In vivo intra-uterine delivery of TAT-fused Cre recombinase and CRISPR/Cas 9 editing system in mice unveil histopathology of Pten/p 53-deficient endometrial cancers. Adv Sci Weinh Baden-Wurtt Ger. 2023;10(32): Article e 2303134.10.1002/advs.202303134 PMC 1064627737749866 · doi ↗ · pubmed ↗
- 7Bogani G, Ray-Coquard I, Concin N, Ngoi NYL, Morice P, Enomoto T, Takehara K, Denys H, Nout RA, Lorusso D, et al. Uterine serous carcinoma. Gynecol Oncol. 2021;162(1):226–234.33934848 10.1016/j.ygyno.2021.04.029PMC 9445918 · doi ↗ · pubmed ↗
- 8Gatius S, Matias-Guiu X. Practical issues in the diagnosis of serous carcinoma of the endometrium. Mod Pathol. 2016;29:S 45–S 58.26715173 10.1038/modpathol.2015.141 · doi ↗ · pubmed ↗