The first molecular detection of equine piroplasmosis in Vietnam and genetic characterization of three co-circulating genotypes of Theileria equi
Thanh Thi Ha Dao, Tamás Szűts, Ngoc Nhu Duong, Duong Thi Quy Troung, Norbert Solymosi, Nóra Takács, Sándor Hornok, Róbert Farkas

TL;DR
This study reports the first molecular detection of equine piroplasmosis in Vietnam and identifies three genotypes of Theileria equi.
Contribution
The first molecular detection and genetic characterization of Theileria equi in Vietnam.
Findings
T. equi was detected in 15.58% of sampled horses in northern Vietnam.
Three genotypes (A, C, and E) of T. equi were identified through phylogenetic analysis.
Babesia caballi was not detected in any of the samples.
Abstract
Equine piroplasmosis, caused by Theileria equi, T. haneyi and Babesia caballi, significantly impacts on the veterinary and economic aspects of the global horse industry. However, many countries, including Vietnam, have not yet conducted epidemiological surveys to determine the prevalence of these haemoparasites. This study aimed to detect T. equi and/or B. caballi infections in horses and to identify their genotypes. Blood samples were collected from 154 apparently healthy horses in eight districts of Hanoi, Thai Nguyen, and Son La provinces located in the northern part of the country. Twenty-four horses (15.58%, 95%CI: 10.70–22.14%) were found to be infected with T. equi across six districts. Babesia caballi was not detected. No significant association was found between gender, host age, and the number of infected horses. Phylogenetic analysis of the 18S rRNA sequences from the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
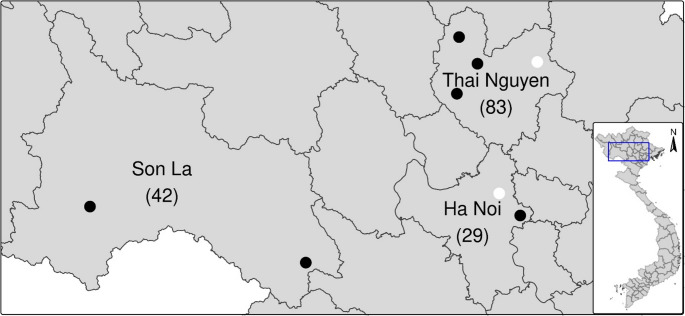 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3- —University of Veterinary Medicine
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Vector-Borne Animal Diseases
Introduction
Equine piroplasmosis is an infectious tick-borne disease caused by two main causative agents, Theileria equi (formerly Babesia equi) and Babesia caballi (Apicomplexa: Piroplasmida) (Rothschild 2013; Wise et al. 2013). Recently, Theileria haneyi has been described (Knowles et al. 2018). Preliminary studies of the small subunit ribosomal RNA gene (18S) have revealed distinct genotypes of T. equi and B. caballi (Bhoora et al. 2009; Qablan et al. 2012; Onyiche et al. 2019). The disease caused by T. equi and/or B. caballi has also been detected in donkeys, mules and zebras (Tirosh-Levy et al. 2020), as well as in non-equid species, including camels (Qablan et al. 2012), waterbucks (Githaka et al. 2014), dogs and cattle (Salim et al. 2019). While horses may clear B. caballi within a few years, T. equi persists for life unless treated (Wise et al. 2013; Scoles and Ueti 2015). Under natural and experimental conditions, more than 30 tick species belonging to various genera, including Hyalomma, Rhipicephalus and Dermacentor, can transmit T. equi and B. caballi trans-stadially (Tirosh-Levy et al. 2020). Other genera, such as Ixodes, Haemaphysalis, and Amblyomma are suspected of being capable of transmitting these hemoparasites, but this has not been confirmed (Scoles and Ueti 2015). Transovarian transmission has been documented for B. caballi, meaning that infected ticks can transmit this species over several generations without re-infection (Scoles and Ueti 2015). Transplacental transmission has been reported for T. equi, which can result in abortion or the birth of an apparently healthy foal that is a carrier (Allsopp et al. 2007; Chhabra et al. 2012). There are numerous reports of the iatrogenic transmission of both T. equi and B. caballi via surgical needles, equipment, and blood transfusions (Rothschild 2013; Wise et al. 2013). The incubation period after infection ranges between 12 and 19 days for T. equi and from 10 to 30 days for B. caballi (Knowles et al. 2018). Equine piroplasmosis is traditionally classified as peracute, acute or chronic forms. However, many infected horses remain asymptomatic or exhibit no specific clinical signs, such as high fever, tachycardia, anorexia, anaemia and jaundice. The peracute form can be life-threatening (Rothschild 2013; Wise et al. 2013; Tirosh-Levy et al. 2020). The disease caused by T. equi and/or B. caballi is endemic in several African, Asian, American and European countries (Onyiche et al. 2019; Tirosh-Levy et al. 2020), as seemingly healthy infected horses are carriers of these pathogens between endemic and non-endemic regions (Chauvin et al. 2009). Equine piroplasmosis has a significant veterinary and economic impact on the horse industry worldwide (Tamzali 2013; Tirosh-Levy et al. 2020). Accordingly, equine piroplasmosis is a reportable disease worldwide, as defined by the World Organisation for Animal Health (WOAH) (https://www.woah.org). Currently, only a few countries, including Japan, New Zealand, Iceland and Ireland, are officially considered to be free from this parasitosis (Mendoza et al. 2024).
Equine piroplasmosis is known to be distributed worldwide (Tirosh-Levy et al. 2020), including many Asian countries (Kamyingkird et al. 2014; Nugraha et al. 2018; Wang et al. 2019; Kumar et al. 2020; Khaing et al. 2025). However, this important parasitic disease has not previously been reported in Vietnam. This study aimed to investigate the occurrence of piroplasmosis in Vietnamese horses using a PCR-based method, which is much more sensitive and specific than traditional smear examinations or serology (Mendoza et al. 2024).
Materials and methods
Study areas and sample collection
A total of 154 horses were examined in eight districts of Hanoi, Thai Nguyen, and Son La provinces located in the northern part of Vietnam. The animals were born in Vietnam and used by their owners to carry wood or work in rice fields. They were kept in communal pastures together with cattle, water buffalo, and small ruminants almost year-round where hard tick species were present, exposing them frequently to tick bites.
Five ml blood samples were collected from the jugular vein of each horse in 2022 and 2023 into EDTA-coated vacutainer tubes, then transferred to the laboratory in iceboxes. This study`s protocol followed the QCVN 01–83:2011/BNN&PTNT (https://cucthuy.gov.vn/en/tieu-chuan-quy-chuan/-/standards/detail/77582) and was approved by the Proposal Committee of the Ministry of Science and Technology of Vietnam (No. 02/2022/HD-NDT). All samples were taken from apparently healthy horses. Furthermore, a questionnaire was used to gather information about the main characteristics of each farm and sampled horse (see Supporting Information).
DNA extraction, polymerase chain reaction (PCR), and sequence analysis
Genomic DNA was extracted from 200 µl of each blood sample according to the manufacturer’s instructions (QIAGEN DNA Blood Mini-Kit, Germany) in the Department of Parasitology, National Institute of Veterinary Research, Hanoi. The DNA samples were stored at − 20^◦^C until use.
Conventional PCR was performed using the forward primer BJ1 5’- GTC TTG TAA TTG GAA TGA TGG– 3’ and the reward primer BN2 5’ –TAG TTT ATG GTT AGG ACT ACG − 3’ to amplify a fragment of the 18S rRNA gene approximately 500 bp in length (Casati et al. 2006). Five µl of extracted DNA were added to 20 µl of reaction mixture containing 1.0 U HotStart Taq Plus DNA Polymerase (5 U/µl) (QIAGEN, Hilden, Germany), 0.5 µl dNTP Mix (10mM), 0.5 µl of each primer (50 µM), 2.5 µl of 10x Coral Load PCR buffer (15mM MgCl_2_ included), and 15.8 µl double-distilled water.
After an initial denaturation step at 95 °C for 10 min, 40 cycles of denaturation at 95 °C for 30 s, annealing at 54 °C for 30 s and extension at 72 °C for 40 s were performed. A final extension step was then performed at 72 °C for five min., after which the sample was kept at 4 °C. DNA of Babesia sp. was used as a positive control. The PCR products were then electrophoresed in 1.5% agarose gel (100 V, 50 min), stained with ethidium-bromide and visualised under ultraviolet light.
The purification and sequencing of the positive PCR products were performed by Eurofins Biomi Ltd. (Gödöllő, Hungary). All sequences were aligned with references using NCBI BLAST, at the National Institutes of Health, USA (http://www.ncbi.nlm.nih.gov). The T. equi 18S rRNA sequences obtained in this study have been deposited in GenBank under the accession numbers PV688145–PV688154 and PX369340–PX369353.
Phylogenetic analysis
To test the placement of the Vietnamese 18S rRNA samples, we used the sequence list and clade assortment from Tirosh-Levy et al. (2020) (see Supporting Information). To provide a robust phylogenetic background to our diagnostic sequences, which are ~ 500 bp long, we selected sequences from GenBank that are over 1500 bp long.
Our aim was to select 10–10 sequences from the five genotypes reported in T. equi (Tirosh-Levy et al. 2020). However, only two 1500-bp-long sequences (AB515310 and EU642507) were available for genotype B from horses. Thus, we supplemented our selection with four sequences (KF597073, KF597077, KF597078 and KF597081) from waterbuck samples, raising the total number of sequences for genotype B to six. In addition to the T. equi samples, we used five Theileria parva sequences (HQ895973–HQ895975; HQ895984–HQ895985) as outgroups. Five T. haneyi sequences (KU647704–KU647708) were used to confirm the presence of T. haneyi in our sequences. A total of 80 terminals were analysed in the analysis. The full list of GenBank accession numbers for the sequences, together with their distribution data and assigned clade (by Tirosh-Levy et al. 2020), can be found in the Supporting Information. The sequences were aligned using MAFFT (Katoh and Toh 2008) and curated in BGME (Criscuolo and Gribaldo 2010). The substitution model was selected using SMS (Lefort et al. 2017) and the tree was analysed using maximum likelihood in PhyML (Guindon et al. 2010) on the NGPhylogeny server (Lemoine et al. 2019). Bootstrap values were calculated according to the method of Lemoine et al. (2018), and the tree was visualised and edited in iTol (Letunic and Bork 2024).
Statistical analysis
The Fisher exact test (Agresti 2002) was applied as independence test on frequency tables within R-environment (v4.5.2, R Core Team 2025).
Results
The occurrence of horse piroplasmosis
Targeting the 18S rRNA gene with specific PCR on 154 examined horses, 24 were found to be positive in six districts of the three provinces (Fig. 1).Fig. 1. The map of Vietnam illustrates the three provinces with the number of samples. Black dots indicate districts where Theileria infected horses were detected, while white dots indicate districts where they were not
The overall prevalence was 15.58% (95%CI: 10.70–22.14.70.14) (Table 1). Based on the sequence analysis, 21 samples were identified as T. equi and three as T. haneyi. Theileria equi was detected in six districts across three provinces (Fig. 1). The prevalence of infection was very similar in Hanoi and Son La provinces. Babesia caballi did not occur. There were no significant differences in the frequency of PCR-positive animals among the provinces (p = 0.4684).Table 1. Prevalence of Theileria equi infection in horses, as detected by molecular methods in three provinces of VietnamProvinceNumber examinedNumber infectedPrevalence (%)95%CIHa Noi29620.699.85–38.39Son La42921.4311.71–35.94Thai Nguyen83910.845.81–19.34Overall1542415.5810.70–22.14
Males had slightly higher odds of T. equi infection than females, but this association was not significant (OR: 1.02, 95%CI: 0.38–2.67, p = 1). There was no significant association between age group (1–3 years old: positive = 7, negative = 48; 3–6 years old: positive = 14, negative = 57; over six years old: positive = 3, negative = 25) and number of horses infected with T. equi (p = 0.4684).
Sequences analysis of T. equi
Based on the NCBI BLAST results, the newly obtained T. equi 18S rRNA sequences (PV688145–PV688154 and PX369340–PX369353) shared 99.09–100% identity to each other and to T. equi sequences in GenBank. The results of the maximum likelihood analysis are shown in Fig. 2. Our results were consistent with those published by Tirosh-Levy et al. (2020), although the placement of genotype A with B + E was not supported. Twelve samples (PV688149–PV688151, PV688153, PX369342–PX369343, PX369345, PX369347–PX369351) formed two distinct groups within genotype A; three samples (PX369340, PX369341, PX369344) formed a distinct group within genotype C; and the remaining nine samples (PV688145–PV688148, PV688152, PV688154, PX369353, PX369352, PX369346) formed a distinct group within genotype E (Fig. 2). Genotypes A and E were detected in 12 and nine horses living in three provinces, respectively. Genotype C was found in two horses in Hanoi province and one horse in Thai Nguyen province. Genotype C was found in two horses in the province Hanoi and one horse in the province Thai Nguyen.Fig. 2. The inferred Maximum likelihood tree of *T. equi *18S rRNA sequences. The tree included 80 sequences and 1548 positions. TN93 + G + I substitution models were used, bootstrap values below 85 are not shown. Five Theileria parva sequences were used as outgroups, five Theileria haneyi sequences were used to identify the species’ clade
Discussion
Since the beginning of the 20th century when Laveran (1901) gave the name Piroplasma equi to the intraerythrocytic parasite found in the blood of horses, equine piroplasmosis has been reported from many European, American and African countries (Onyiche et al. 2019; Tirosh-Levy et al. 2020). While it was hypothesised over a hundred years ago that piroplasmosis had occurred in a herd of mules in French Indochina (Schein 1917), there was no information available for many decades regarding the occurrence of this economically significant parasitic disease in Asia. However, since the 2010 s serological and/or molecular studies have revealed the presence of equine piroplasmosis in various Asian countries, with one or both haemoparasite species being detected (Munkhjargal et al. 2013; Kamyingkird et al. 2014; Nugraha et al. 2018; Ybañez et al. 2018; Wang et al. 2019; Zhao et al. 2020; Kumar et al. 2020; Khaing et al. 2025). To the best of our knowledge, no serological or molecular examinations of this tick-borne disease had been carried out in Vietnam prior to this survey.
The results revealed that 24 out of 154 blood samples collected in Vietnam contained the DNA of T. equi, which is the predominant species in some Asian countries (Tirosh-Levy et al. 2020; Kumar et al. 2020). Conversely, a higher prevalence of B. caballi than T. equi was reported in Mongolia (Munkhjargal et al. 2013). Neither B. caballi nor dual infections with the two protozoan species were identified in Vietnam. There are two possible explanations for the absence of B. caballi. Either this species does not occur among horses in the provinces studied, or it was present but efficiently eliminated by the host immune system. Another possibility is that using a single-round PCR rather than a nested PCR did not allow B. caballi to be detected, given that its parasitemia is generally extremely low. This is contrasts with the lifelong persistence of T. equi in untreated horses (Brüning 1996).
When we compared our results (15.58%) with those of other studies in Asia, we found that a higher prevalence was reported in China (Wang et al. 2019; Zhao et al. 2020), Iran (Kalantari et al. 2022) and the Philippines (Ybañez et al. 2018), while a lower prevalence was found in India (Kumar et al. 2020), and Myanmar (Khaing et al. 2025). This variation in equine theileriosis prevalence across countries may be attributed to differences in competent vector occurrence and abundance, tick environmental factors, the application of various methods, and the efficacy of tick control measures (Onyiche et al. 2019). No significant correlation was found between the PCR results and the age of the horses. However, Ruegg et al. (2007) reported that a positive correlation was found between host age and T. equi infection in the surveyed provinces. As in a previous study (Steinman et al. 2012; gender was not found to be a risk factor for T. equi infection in this study. However, other authors (Ruegg et al. 2007; Moretti et al. 2010) did find a correlation between gender and infection. No horses were found to be infested with ticks during the sampling period. Therefore, we could not obtain any relevant data on tick species that could carry T. equi. However, most of the suspected tick genera (Hyalomma, Dermacentor, and Haemaphysalis), that may act as vectors (De Waal 1992; Scoles and Ueti 2015), are present in Vietnam (Hornok et al. 2024; Ngoc et al. 2025) as are stable flies, one of the most important mechanical vectors of Theileria spp. including T. equi (Hornok et al. 2020) which are commonly found on pastures. Furthermore, iatrogenic transmission cannot be ruled out, because contaminated needles and other equipment have been reported by several authors (Rothschild 2013; Wise et al. 2013) as a source of such infections.
Preliminary studies have revealed five 18S rRNA genotypes of T. equi (A, B, C, D and E) (Bhoora et al. 2009; Qablan et al. 2012; Onyiche et al. 2019). The genetic diversity of T. equi may be important for the transmission of this pathogen (Manna et al. 2018; Tirosh-Levy et al. 2020). However, more molecular epidemiological data are needed to confirm this. This study provides the first data on the genetic diversity of T. equi in Vietnam. Three genotypes (A, C and E) were identified through sequence analysis of T. equi samples taken from 24 horses. The most prevalent genotype was A, which has been reported in many countries and is more commonly associated with clinical theileriosis than other genotypes (Tirosh-Levy et al. 2021). Genotype E was found in nine animals in the present study and has primarily been reported in China (Chen et al. 2022) and Mongolia (Otgonsuren et al. 2024). Genotype C was found in three horses. A comparison of the phylogenetic tree from the present study with that from Knowles et al. (2018) suggests that a genetically homogeneous group within genotype C most likely corresponds to T. haneyi. This is corroborated by sequences EU642511 and EU888905 from South Africa, as well as the T. haneyi reference sequences KU647704–KU647708 included in our study (Fig. 2). We also included EU888903, which Knowles et al. (2018) identified as T. equi, thus marking the species limits. As T. equi has been assigned to genotype C rather than T. haneyi (Tirosh-Levy et al. 2021), the taxonomy of T. haneyi remains uncertain. If it is accepted as valid, then our records will be the first from Vietnam.
The time and place of the arrival of T. equi, the agent of theileriosis in horses, in Vietnam is unknown. However, based on reports by Laveran (1901) and Schein (1917), it is evident that this haemoparasite species was present in northern Vietnam at the beginning of the 20th century, resulting in clinical symptoms and death in mules in the Haiphong area.
Conclusion
In conclusion, the results of the first molecular survey suggest that T. equi, the agent that causes equine piroplasmosis, is present in northern Vietnam, albeit in a subclinical form. Further studies are needed to assess the risk posed by T. equi and B. caballi in other parts of the country. These studies should be accompanied by tick vector surveillance and educational programmes for veterinarians about this notifiable disease.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary File 1 (JPG 105 KB)
Supplementary File 2 (PDF 306 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1R Core Team (2025) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org