Potential role of circ_WHSC1 and miR-145-5p in breast cancer promotion
Maryam Abtin, Asghar Hosseinzadeh, Nahid Nafisi, Ramesh Omranipour, Leyla Sahebi, Mohsen Ahmadi, Soudeh Ghafouri-Fard, Abbas Shakoori

TL;DR
This study explores how circ_WHSC1 and miR-145-5p interact in breast cancer and suggests they may play a role in its development.
Contribution
The study identifies a potential molecular axis involving circ_WHSC1 and miR-145-5p in breast cancer pathogenesis.
Findings
Circ_WHSC1 levels were significantly higher in breast tumors compared to normal tissues.
MiR-145-5p levels were significantly lower in breast tumors compared to normal tissues.
A weak negative correlation was found between circ_WHSC1 and miR-145-5p levels.
Abstract
This study aims to investigate the potential interplay between circ_WHSC1 and miR-145-5p in breast cancer pathogenesis using in silico tools, assess their clinical relevance, and evaluate the diagnostic utility of circ_WHSC1 in clinical samples as a biomarker for breast cancer. This multi-component study employed a combination of bioinformatic analyses and laboratory validation. First, in silico tools were used to investigate miR-145-5p and circ_WHSC1 using public databases. Subsequently, their expression, correlation, and clinical relevance were experimentally assessed in a cohort of breast cancer patients. Circ_WHSC1 level was significantly higher in breast tumor compared with patient-matched adjacent normal tissues (4.17-fold change, p-value <0.01). Additionally, significant reduction of miR-145-5p (9.11-fold downregulation, p-value <0.01) level was detected in breast tumors…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
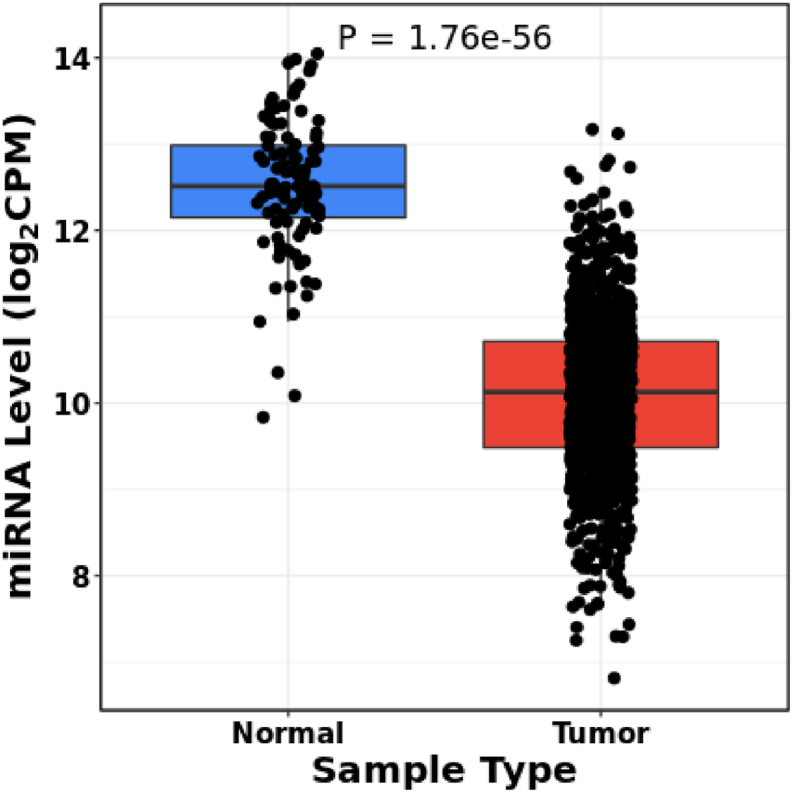 Figure 1
Figure 1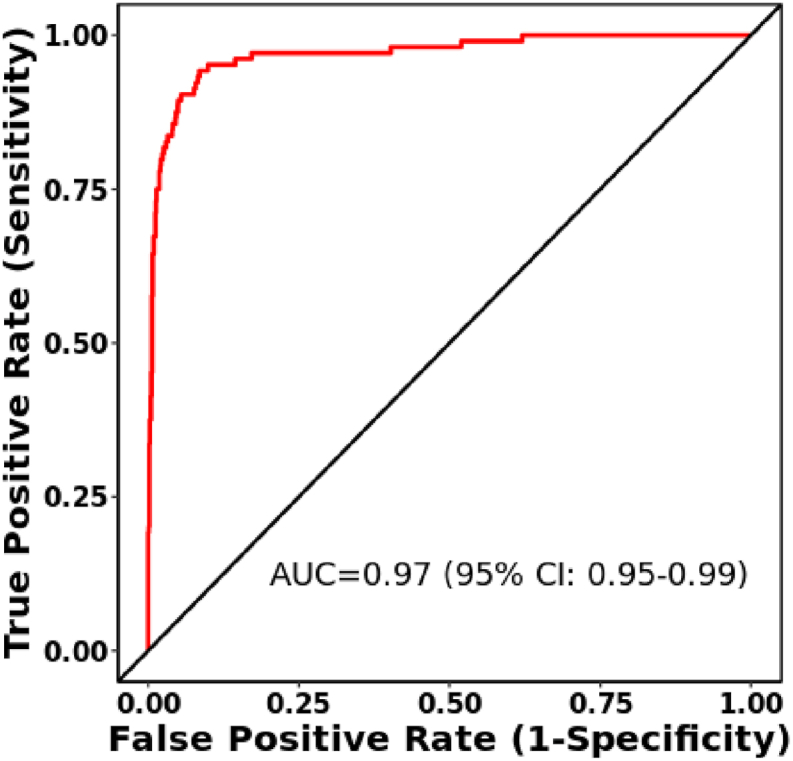 Figure 2
Figure 2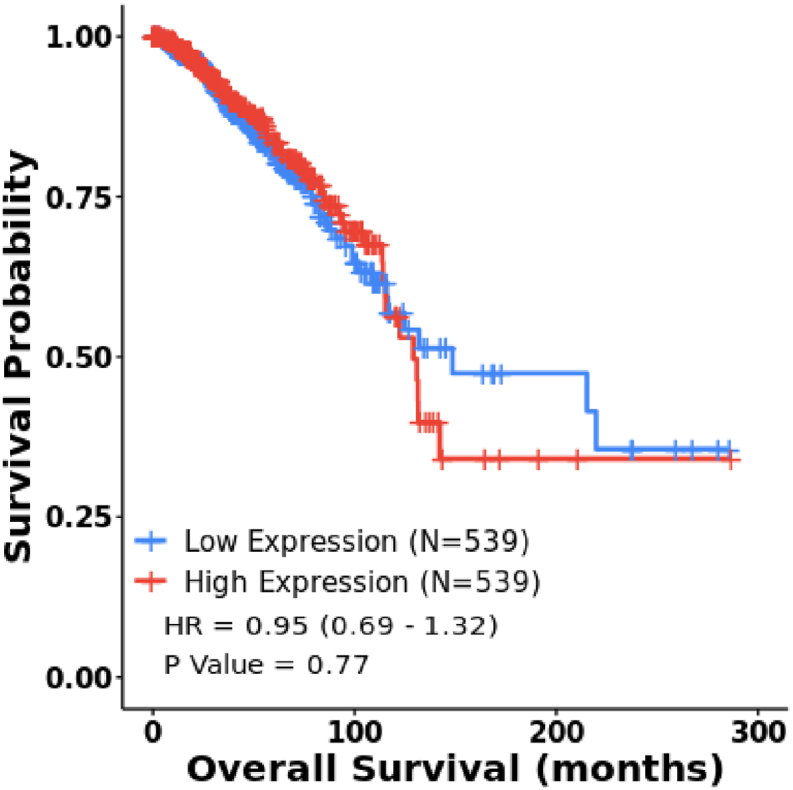 Figure 3
Figure 3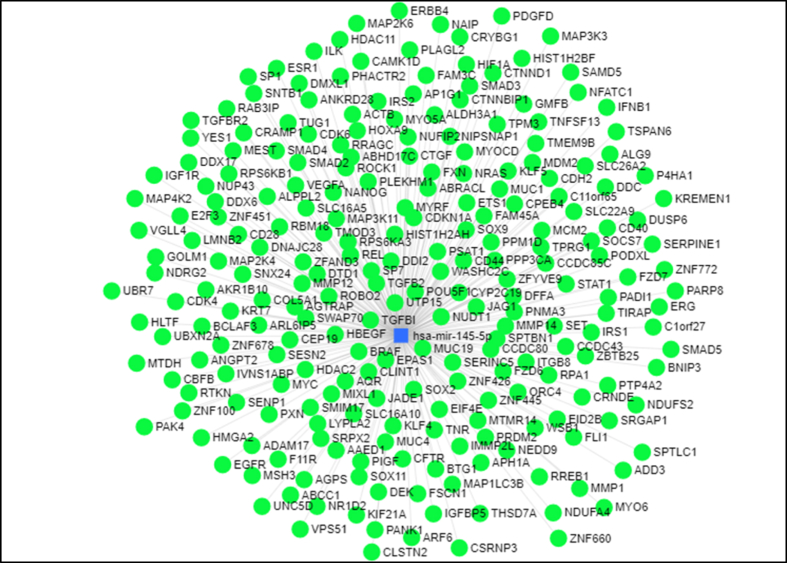 Figure 4
Figure 4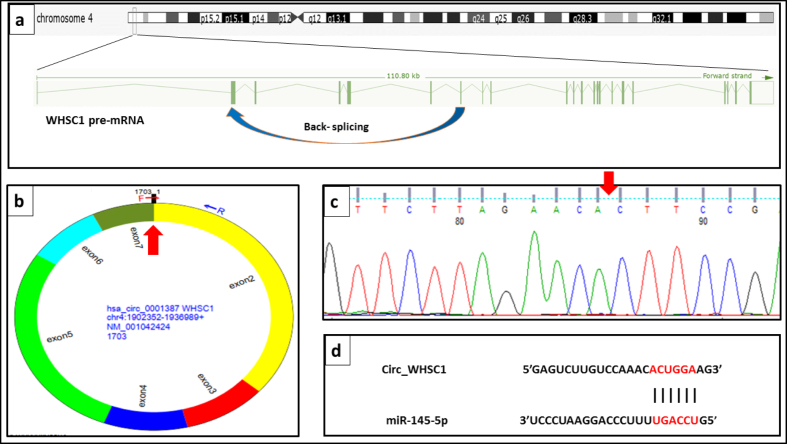 Figure 5
Figure 5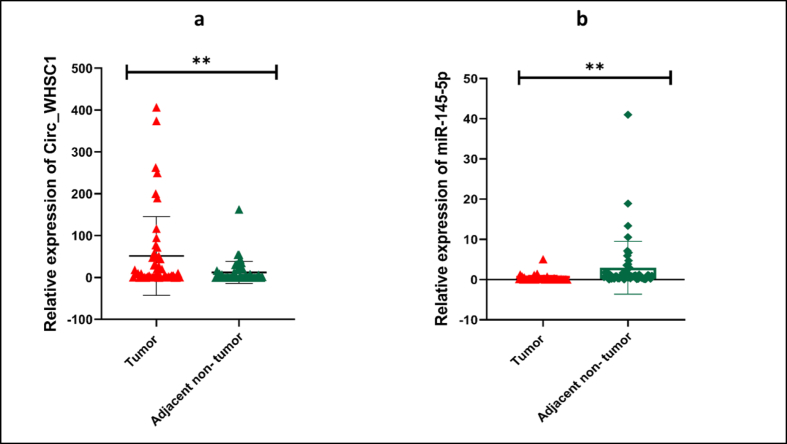 Figure 6
Figure 6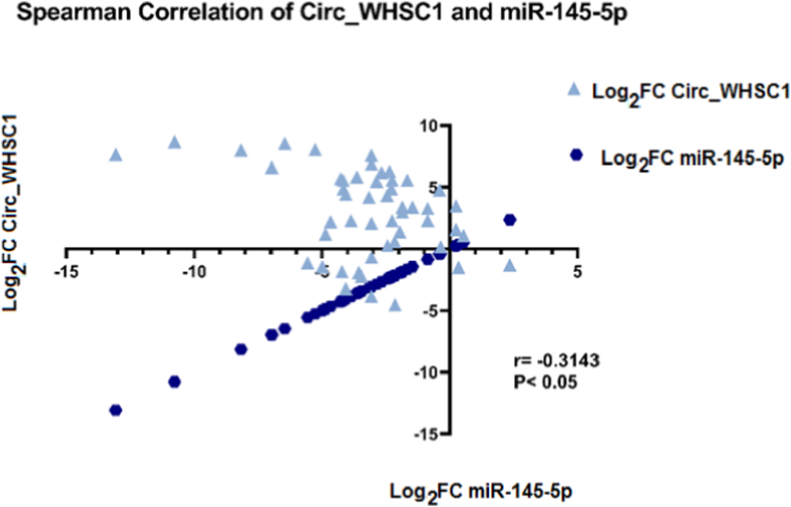 Figure 7
Figure 7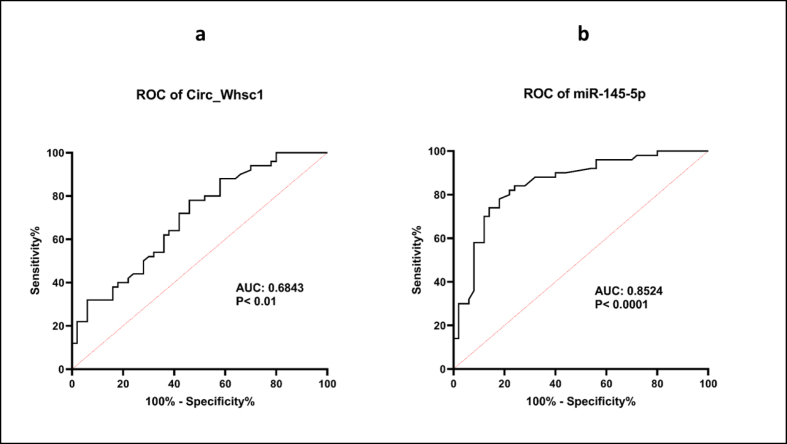 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsConnective Tissue Growth Factor Research · Circular RNAs in diseases · Hippo pathway signaling and YAP/TAZ
Introduction
1
Globally, breast cancer is the most common cancer and a leading cause of cancer-related death among women [1]. Despite advances in the therapeutic methods, breast cancer has a high incidence and death rate due to its multifactorial nature and tumor heterogeneity, which lead to tumor metastasis, recurrence, and treatment resistance [2]. Hence, identification of new molecular targets has pronounced implication to diminish breast cancer mortality and expand clinical efficacy [[3], [4], [5]].
miRNAs are regulatory non-coding transcripts that target certain mRNAs and regulate expression of the encoded proteins [6,7]. There are several biological processes that miRNAs participate in, such as cell proliferation, signal transduction, apoptosis, etc. So, it is obvious that dysregulation of miRNAs plays an imperative role in cancer pathogenesis [8]. hsa-miR-145-5p is frequently downregulated in various cancers, including breast cancer [9,10]. Established roles of this miRNA include suppression of tumor progression, induction of tumor apoptosis, and modulation of cancer stemness [[11], [12], [13]] by regulating expression of numerous genes.
CircRNAs have a key role in down regulation of miR-145-5p in cancer [[14], [15], [16]]. CircRNAs are a class of endogenous noncoding RNAs characterized by a covalently closed continuous loop, formed by back-splicing of single-stranded RNA transcripts. These transcripts perform their biological functions mainly by acting as microRNA (miRNA) sponge [17,18]. Several studies have shown the link between circRNA dysregulation and tumorigenesis [[19], [20], [21], [22]]. Recently, circRNAs represented a new group of biomarkers for diagnosis of malignancy, refining prognosis, and targeted therapy [23,24]. However, the functional roles of many circRNAs in cancer remain poorly understood [25].
Recent studies have shown that circ_WHSC1 (hsa_circ_0001387) contributes to tumor proliferation, invasion, metastasis, and drug resistance in ovarian [26], lung [27,28], endometrial [29] and nasopharyngeal [30] cancers. It has been demonstrated that circ_WHSC1 promotes ovarian cancer proliferation via targeting miR-145-5p [26]. Nevertheless, the potential roles of circ_WHSC1 in breast tumorigenesis and its relationship with miR-145-5p in breast cancer have remained unclear. So, we designed this multi-component study and employed a combination of bioinformatic analyses and laboratory validation. We integrated in silico analyses of publicly available datasets (TCGA via CancerMIRNome and ENCORI) with experimental validation on 50 paired breast tumor and adjacent normal tissue samples. Bioinformatic analyses included differential expression, survival, diagnostic receiver operating characteristic (ROC), and functional enrichment (Gene Ontology [GO]/KEGG) assessments for miR-145-5p, as well as interaction network mapping for circ_WHSC1. Clinically, circ_WHSC1 and miR-145-5p expression levels were quantified via qRT-PCR, and their diagnostic potential and correlation with clinicopathological features were statistically evaluated.
Methods and materials
2
Bioinformatics analysis on hsa-miR-145-5p expression level and overall survival outcomes of breast cancer in TCGA cohort
2.1
We analyzed the expression profile of hsa-miR-145-5p in breast cancer using the CancerMIRNome platform. This platform serves as a resource for assessment of miRNAs expression, and their diagnostic and prognostic relevance across various cancer types [31]. We utilized the related data obtained from TCGA cohort which included the expression data of 1078 breast cancer samples and 104 normal controls. We also assessed the diagnostic relevance of hsa-miR-145-5p in breast cancer using this tool. Additionally, we investigated the relationship between the expression levels of hsa-miR-145-5p and the overall survival (OS) outcomes of breast cancer, utilizing data from the CancerMIRNome database. A log-rank p-value <0.05 was considered statistically significant.
Functional enrichment analysis
2.2
To elucidate the functional enrichment of mentioned miRNA, which encompasses GO and KEGG pathway, we employed the pathway module available in the ENCORI database (https://rnasysu.com/encori/index.php). This database performs analyses based on the predicted target genes of the miRNA. The GO analysis was categorized into Biological Process (BP), Molecular Functions (MF), and Cellular Components (CC). The identified terms were organized based on log_10_ adjusted p-value (FDR) threshold of less than 0.01.
Collection and handling of breast tumor and non-tumor tissue samples
2.3
All specimens were obtained from the patients referred to two surgery centers, in Tehran, Iran. All samples were gathered in RNA Later reagent (Biobasic, Canada) and kept based on instructions. A total of 50 pairs of tumor and corresponding non-tumor tissue samples were examined by a pathologist. Tissues were fresh-frozen and transferred to −70 °C freezer until being used for RNA extraction. Patients experienced neither radiotherapy nor chemotherapy before sampling. The protocol was certified by the ethical review committee of the Tehran University of Medical Sciences under the ethical ID of IR.TUMS.MEDICINE.REC.1401.272, in accordance with the Helsinki Declaration. Patient agreement was attained before the samples were taken.
RNA extraction and cDNA synthesis
2.4
Total RNA was retrieved using TRIzol reagent (Invitrogen). RNA integrity and concentration were assessed using agarose gel electrophoresis and spectrophotometry, respectively. For cDNA production, the ExcelRT™ 1st Strand cDNA Synthesis Kit (SMOBIO) was applied. The specific stem-loop RT primers for miR-145-5p and snRNA U6 were added to the reaction mix and cDNA was synthesized based on kit direction. The sequence of circ_WHSC1 was attained from the Circbase database, and specific divergent primers were designed by CircPrimer 2.0 software.
Real-time PCR
2.5
Real-time PCR was performed using RealQ Plus 2x Master Mix Green low Rox (Ampliqon) via Light-Cycler96 Roche. Reactions were carried out in 10 μl volume in duplicate. Gene expressions were quantified using the 2^−ΔΔCT^ method, normalized to GAPDH (for circ_WHSC1) or U6 (for miR-145-5p). PCR products were evaluated by melting curve examination and electrophoresis on agarose gel. Back-splicing-derived circularization of circ_WHSC1 was confirmed through Sanger sequencing using divergent primers spanning the junction site (primer sequences provided in Table 1).Table 1. Primers sequences.Table 1. Gene symbolPrimer nameSequence (5′ to 3′)TmSize of amplicon (bp)Source of primerCircWHSC1ForwardCACAGTCTTCGGAAGTGTTCT57.90134This studyReverseACTGCCGAGGATTTCTGGTG60.04miR-145-5pStem- Loop RT primerGTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGGGATOur previous study [32]ForwardGGCTTAGTCCAGTTTTCCCAG58.8669ReverseGTGCAGGGTCCGAGGT58.01U6snRNAStem- Loop RT primerGTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAAATATOur previous study [32]ForwardGCTTCGGCAGCACATATACTAAAAT60.0591ReverseCGCTTCACGAATTTGCGTGTCAT63.00GAPDHForwardAAGGCTGAGAACGGGAAGCT61.4891Our previous study [32]ReverseCAGCATCGCCCCACTTGATT61.03
Statistical analysis
2.6
Statistical assessments were done by GraphPad Prism 8.4.3 and SPSS v.26. Relative expression of circ_WHSC1 and miR-145-5p was compared in breast tumors and adjacent tissues by applying the paired sample t-test. The correlation of expression of circ_WHSC1 and miR-145-5p was determined using the Spearman correlation factor. The relationship between the expression of circ_WHSC1 and clinicopathological characteristics of patients was judged using Mann-Whitney and Kruskal-Wallis tests. Furthermore, ROC curves were generated using GraphPad Prism. The p-value <0.05 was considered to describe the statistical significance in all assessments.
Results
3
Bioinformatics analysis on hsa-miR-145-5p expression level and overall survival outcomes of breast cancer in TCGA cohort
3.1
Expression analysis utilizing the CancerMIRNome database showed that hsa-miR-145-5p was significantly down-regulated in 1078 breast cancer specimens compared to 104 controls (Fig. 1; p-value = 1.7e-56).Fig. 1. An examination of levels of hsa-miR-145-5p in breast cancer tissues (n = 1078) compared to normal tissues (n = 104) was conducted utilizing the CancerMIRNome database. A p-value of less than 0.05 was deemed statistically significant.Fig. 1
The analysis of the ROC curve using the CancerMIRNome database indicated that hsa-miR-145-5p may possess significant diagnostic value for individuals diagnosed with breast cancer, as evidenced by an Area Under the Curve (AUC) of 0.97, a p-value <0.0001, and a 95 % Confidence Interval ranging from 0.95 to 0.99 (Fig. 2).Fig. 2. The diagnostic value of the hsa-miR-145-5p in breast cancer using receiver operating characteristic (ROC) curve analyses (Tumor (n = 1078) vs. Normal (n = 104)) via the CancerMIRNome database. A p-value <0.05 was regarded as significant (AUC: area under curve).Fig. 2
Overall survival (OS) analysis of hsa-miR-145-5p in breast cancer using in silico tools
3.2
As demonstrated in Fig. 3, the survival analysis by the CancerMIRNome database indicated no significant associations between lower expression of hsa-miR-145-5p and OS of breast cancer patients.Fig. 3. The overall survival curve of patients with breast cancer based on hsa-miR-145-5p expression according to the CancerMIRNome (total number of patients = 1078). Log-Rank p-value <0.05 was considered statistically significant. N: Sample count.Fig. 3
GO and KEGG pathway analyses based on in silico tools
3.3
First, we demonstrated the interactive network between miR-145-5p and its targets interacting network in breast cancer (Fig. 4).Fig. 4. Interacting network between hsa-miR-145-5p and its hypothetical targets. The blue square represents hsa-miR-145–5p and green circles indicate target genes of this miRNA (www.mirnet.ca).Fig. 4
The GO analysis unearthed that the targets of hsa-miR-145-5p were mainly enriched in Cellular Macromolecule Localization, Establishment Of Protein Localization, Positive Regulation Of Nucleobase Containing Compound Metabolic Process in BP (Table 2); in Enzyme Binding, Protein Containing Complex Binding, Ribonucleotide Binding in MF (Table 3); and in Organelle Subcompartment, Golgi Apparatus, Endoplasmic Reticulumin CC (Table 4). The KEGG pathway analyses also indicated that these genes were enriched in Endocytosis, Focal Adhesion, Regulation Of Actin Cytoskeleton pathways (Table 5).Table 2. Biological Processes (BP) terms for hsa-miR-145-5p.Table 2BP termslog_10_ (p-value)log_10_ (FDR)Cellular Macromolecule Localization−99.36547−95.60543Establishment Of Protein Localization−84.14343−80.8605Positive Regulation Of Nucleobase Containing Compound Metabolic Process−84.14396−80.68494Positive Regulation Of Biosynthetic Process−80.58294−77.42496Intracellular Transport−80.376−77.31493Regulation Of Protein Modification Process−79.54705−76.56516Regulation Of Intracellular Signal Transduction−76.46086−73.54591Positive Regulation Of Protein Metabolic Process−71.38218−68.52523Positive Regulation Of Molecular Function−71.18779−68.38199Regulation Of Phosphorus Metabolic Process−70.83962−68.07957Table 3Molecular Function (MF) terms for hsa-miR-145-5p.Table 3MF termslog_10_ (p-value)log_10_ (FDR)Enzyme Binding−104.25187−101.19496Protein Containing Complex Binding−70.88172−68.12584Ribonucleotide Binding−67.29546−64.71567Transcription Regulator Activity−61.19369−58.73885Identical Protein Binding−59.96512−57.60718Adenyl Nucleotide Binding−57.65983−55.38108Sequence Specific Dna Binding−57.1824−54.9706Rna Binding−56.74355−54.58974Cytoskeletal Protein Binding−52.24391−50.14125Dna Binding Transcription Factor Activity−46.43723−44.38033Table 4Cellular Component (CC) terms for hsa-miR-145-5p.Table 4CC termslog_10_ (p-value)log_10_ (FDR)Organelle Subcompartment−69.32104−66.45475Golgi Apparatus−67.84869−65.28343Endoplasmic Reticulum−60.08455−57.69538Endosome−50.38498−48.12075Neuron Projection−46.2645−44.09718Vacuole−44.29713−42.27594Anchoring Junction−44.30292−42.21479Chromosome−44.06045−42.09725Plasma Membrane Region−42.76317−40.85113Vesicle Membrane−40.68108−38.81479Table 5KEGG terms for hsa-miR-145-5p.Table 5KEGG termslog_10_ (p-value)log_10_ (FDR)Endocytosis−21.82506−19.62916Focal Adhesion−19.40604−17.51117Regulation Of Actin Cytoskeleton−18.41674−16.69797Mapk Signaling Pathway−17.72381−16.12997Pathways In Cancer−17.13598−15.63905Axon Guidance−14.10341−12.68566Insulin Signaling Pathway−11.63633−10.28553Neurotrophin Signaling Pathway−11.46096−10.16815P53 Signaling Pathway−11.27545−10.03379Lysosome−9.97121−8.77531
Expression analysis of circ_WHSC1 and miR-145-5p in clinical specimens
3.4
Sanger sequencing was employed for confirming the back splice junction resulted from joining of exon 2 and exon 7 of WHSC2 gene as host gene of circ_WHSC1. Fig. 5 shows a schematic representation of circ_WHSC1 biogenesis (Fig. 5a), divergent primer designed for amplification of circ_WHSC1 (Fig. 5b), verification of the back-splice junction site of circ_WHSC1 (Fig. 5c), and interaction site of circ_WHSC1 and miR-145-5p (Fig. 5d).Fig. 5a. schematic representation of circ_WHSC1 biogenesis b. circ_WHSC1 divergent primer illustrated by circprimer 2.0. c. the back-splice junction site of circ_WHSC1 was verified by Sanger sequencing d. Interaction site of circ_WHSC1 and miR-145-5p (circinteractome.nia.nih.gov).Fig. 5
Expression levels of circ_WHSC1 and miR-145-5p in clinical samples were normalized to GAPDH and U6, respectively. We found significant up-regulation of circ_WHSC1 in breast tumors compared with corresponding neighboring normal tissues (4.17-fold change, P = 0.0026) (Fig. 6a). Additionally, significant reduction of miR-145-5p (9.11-fold downregulation, P = 0.0068) level was detected in breast tumors compared with non-tumor tissues (Fig. 6b). Detailed statistics of expression ratios are shown in Table 6.Fig. 6. Scatter plots with bar showing relative expression of circ_WHSC1 (a) and miR-145-5p (b) in breast cancer tissues (n = 50) relative to normal neighboring tissue (n = 50) (Mean and standard deviation values are shown, ∗∗: p-value <0.01).Fig. 6. Table 6Detailed statistics of expression ratio of circ_WHSC1 and miR-145-5p in breast cancer vs. corresponding neighboring normal tissues (Paired t-test results are shown).Table 6. Parameterscirc_WHSC1**miR-145-5pP value0.00260.0068One- or two-tailed P value?Two-tailedTwo-tailedt, dft = 3.178, df = 49t = 2.825, df = 49Number of pairs5050R squared (partial eta squared)0.17090.1400Average expression of tumor samples group (T)51.468074120.321544Average expression of non-tumor samples group (N)12.321318842.93108Fold change4.1771 (T/N)9.1156 (N/T)
Correlations between expressions of circ_WHSC1 and miR-145-5p
3.5
Correlation analyses between expression quantities of circ_WHSC1 and miR-145-5p were achieved by Spearman correlation coefficient. As demonstrated in Fig. 7, a weak negative correlation was perceived between levels of circ_WHSC1 and miR-145-5p (r = −0.3143, 95 % CI = −0.5509 to −0.03094 and p-value = 0.0262).Fig. 7. Point graph in XY axes showing Spearman correlation of miR-145-5p and circ_WHSC1 expression (Number of tumor samples = 50, r = −0.3143, p-value <0.05).Fig. 7
The implication of circ_WHSC1 as a marker
3.6
ROC curve was plotted to appraise the diagnostic biomarker power of mentioned genes. We used gene expression values in tumors and adjacent non-tumor tissues for producing ROC curves. This assessment discovered that circ_WHSC1 has a weak power to distinguish between tumor tissue and adjacent normal tissues (AUC = 0.683, Standard error = 0.05301, 95 % CI = 0.5795 to 0.7873, p-value = 0.0016) with 71 % specificity and 70 % sensitivity (Fig. 8a). Moreover, ROC curve was generated for miR-145-5p and demonstrated that miR-145-5p has moderate power in discriminating between these two sets of samples (AUC = 0.8524, Standard error = 0.0386, 95 % CI = 0.7767 to 0.9281, 88 % specificity and 88 % sensitivity, p-value <0.0001) (Fig. 8b).Fig. 8. Receiver operating characteristic (ROC) curve of a. circ_WHSC1 (P < 0.01) and b. miR-145-5p (P < 0.0001) in breast cancer patients (Tumor (n = 50) vs. Normal (n = 50), AUC: area under curve).Fig. 8
Correlations of circ_WHSC1 f levels and clinicopathological characteristics
3.7
The clinical information of patients such as age, tumor dimension, grade, hormone receptors and TNM stage exhibited in Table 7. Correlation examinations indicated that up-regulation of circ_WHSC1 in breast tumors was linked with lymph node invasion (p-value = 0.005), HER2 negativity (p-value = 0.031) and positive family history (p-value = 0.012).Table 7. Correlation of circWHSC1 expression and clinicopathological data (N = 50).Table 7. SubclassNumber of patients (%)Median of Circ_Whsc1 levelInterquartile Range (IQR)Mean of Circ_Whsc1 levelSDp-valueAge≤45154.45.573.043.500.84>45352.975.42.853.55Tumor diameter<2 cm70.576.992.023.490.12–5 cm383.424.883.473.3>5 cm50.276.53−0.163.42Invasion to lymphyes294.874.504.103.410.005no211.375.101.252.98Grade1133.476.953.553.930.632312.975.402.583.44363.325.223.173.23Stage1110.576.82.293.580.332323.324.153.163.37364.996.23.813.5641ERP413.374.893.163.40.18N91.166.821.263.9PRP373.375.22.963.440.7N122.667.082.513.89HER2P7−0.683.770.112.990.031N433.474.773.363.40KI67<16193.477.553.313.880.4716 or >16313.275.202.663.3Family historypresent135.574.884.993.000.012absent372.275.872.173.40
Discussion
4
Breast cancer is the most common malignancy among women [1]. This malignancy is associated with dysregulation of several circRNAs [33]. Most of these molecules exert their role in breast cancer progression through sponging miRNAs [33]. Thus, exploring the role of circRNAs/miRNAs axes in this cancer is important in identification of biomarkers.
Circ_WHSC1 is originated from exonic back-splicing of WHSC1 gene, a histone methyl transferase that is controlled by EZH2 and has imperative functions in ovarian carcinogenesis [34]. The impact of circ_WHSC1 in the carcinogenesis has been evaluated in several tissues [35]. Moreover, circ_WHSC1 has been found to stimulate ovarian tumorigenesis through adsorbing miR-145 and another miRNA, namely miR-1182 [26]. We evaluated expression level of this circRNA and miR-145 in breast cancer samples. Similar to what reported in ovarian cancer, we found significant up-regulation of circ_WHSC1 and down-regulation of miR-145-5p in breast tumors compared with corresponding adjacent normal tissues. A weak negative correlation was detected between levels of circ_WHSC1 and miR-145-5p, suggesting the similar effect of this circRNA on expression of mentioned miRNA in breast and ovarian malignancies. An earlier study has shown that circ_WHSC1 accelerates malignancy and glycolysis of triple negative breast cancer cells through regulation of miR-212-5p/AKT3 axis [36]. Another study has revealed the importance of circ_WHSC1/miR-195-5p in the regulation of FASN/AMPK/mTOR axis and promotion of breast carcinogenesis [37]. Thus, the oncogenic effects of circ_WHSC1 in breast cancer are exerted through different routes. We also assessed TCGA data of miR-145-5p in breast cancer and demonstrated similar results with the findings of current study. However, this data did not show correlation between expression level of miR-145-5p and OS of patients. The lack of correlation with patient survival in TCGA could be due to molecular subtype distribution or sample size. Thus, further assessments in larger cohorts of patients are needed.
Based on the in silico assays, this miRNA was found to be linked with a number of pathways, including those related with endocytosis, focal adhesion, regulation of actin cytoskeleton, as well as MAPK and P53 signaling pathways. Meanwhile, in silico analyses suggested interaction between circ_WHSC1 and a number of RNA binding proteins that are involved in the carcinogenesis. Among these proteins is HUR, a protein that is involved in mRNA transport from nuclear compartment to cytoplasm, and regulation of stability and translation of mRNAs. Dysregulation of HUR has been found to be associated with cancer [38]. AGO2 protein has also been reported to participate in transcriptional activation in breast cancer cells, and stimulate cell growth through induction of expression of the progesterone receptor [39]. Thus, the circ_WHSC1/miR-145-5p axis can be involved in breast carcinogenesis through different mechanisms. However, it is worth emphasizing that these are computational predictions and not experimentally confirmed. Such interactions should be validated through immunoprecipitation (RIP) or crosslinking immunoprecipitation (CLIP).
ROC curve analysis revealed weak performance of circ_WHSC1 as a biomarker for breast cancer. However, the data provided by CancerMIRNome database revealed excellent performance of miR-145-5p as a diagnostic marker in breast cancer. Thus, while miR-145-5p has stronger diagnostic performance, circ_WHSC1 has potential use as part of a biomarker panel, rather than as a standalone marker.
Up-regulation of circ_WHSC1 in breast tumors was linked with lymph node invasion, HER2 negativity and positive family history. Since circ_WHSC1 enhances cell viability, migratory potential and invasion in ovarian tumors [26], it is not surprising that its up-regulation in breast cancer is associated with lymph node metastasis. Moreover, studies in breast cancer specimens and animal models support similar roles for circ_WHSC1 in this type of cancer [36,37]. Finally, this finding is supported by the involvement of circ_WHSC1 in several cancer-related pathways, its interaction with RNA binding proteins and its transcriptional activity role. Overall, our findings suggest that the circ_WHSC1/miR-145-5p axis is a potential molecular pathway contributing to breast carcinogenesis and may serve as both a diagnostic biomarker and a therapeutic target, pending functional and clinical validation.
To confirm the regulatory relationship between circ_WHSC1 and miR-145-5p, functional assays such as circ_WHSC1 knockdown or overexpression experiments followed by miR-145-5p quantification are recommended. Luciferase reporter assays using the predicted binding sites would help validate direct interactions. Besides, to definitively validate the circular nature of circ_WHSC1, future experiments should include RNase R digestion assays. This treatment selectively degrades linear RNAs while preserving circular transcripts, providing stronger evidence of circularity beyond divergent primer PCR and Sanger sequencing. Additionally, elucidation of downstream targets requires further experimental steps. In fact, although we observed a significant downregulation of miR-145-5p, the downstream mRNA targets of this miRNA in breast cancer require further exploration. Transcriptomic profiling following miR-145-5p restoration could identify key downstream genes involved in tumor progression.
The interaction of circ_WHSC1 with RNA-binding proteins such as AGO2, HUR, FMRP, and FUS, as predicted in silico, suggests a broader role in post-transcriptional regulation. RNA immunoprecipitation (RIP) or crosslinking immunoprecipitation (CLIP) assays could confirm these interactions experimentally. Future studies using in vivo models, such as xenograft or genetically engineered mouse models, would be valuable in confirming the oncogenic role of circ_WHSC1 and its impact on tumor growth, metastasis, and therapeutic response.
Although we demonstrated associations between circ_WHSC1/miR-145-5p levels and some clinicopathological features, larger multicenter studies are necessary to validate these biomarkers across diverse patient populations and breast cancer subtypes. Finally, given the oncogenic features of circ_WHSC1 and tumor-suppressive role of miR-145-5p, therapeutic strategies such as antisense oligonucleotides (ASOs) against circ_WHSC1 or miRNA mimics for miR-145-5p restoration may be promising and warrant preclinical investigation.
In fact, the regulatory effect of circ_WHSC1 on miR-145-5p requires experimental support via knockdown/overexpression and luciferase assays to demonstrate direct interaction and functional consequence. This data might facilitate design of novel therapeutics that targets this axis. Moreover, a biomarker panel consisted of circ_WHSC1 on miR-145-5p can be designed for diagnostic purposes in breast cancer. Finally, although circRNAs and miRNAs hold significant potential in the context of personalized cancer therapy due to their expression-based specificity, the patient cohort selected in this study presents demographic heterogeneity. Given that expression data in epigenetic research are highly sensitive and influenced by environmental factors, we state this point as another limitation of our study. While we used patient-matched adjacent normal breast tissue as the control—a standard practice that rigorously controls for inter-individual genetic and environmental variability—it is recognized that such tissue may not represent a truly “healthy” state. Adjacent normal tissue can be influenced by the local tumor microenvironment or exhibit a “field effect,” potentially harboring subtle molecular alterations. Although the use of tissue from completely healthy donors could theoretically circumvent this, obtaining such samples is ethically and practically challenging, as it requires invasive procedures without clinical benefit for the donor. Therefore, our internal control design, while optimal for identifying tumor-specific dysregulation, may slightly underestimate the magnitude of differential expression between cancerous and completely normal breast epithelium. Future studies incorporating larger, multi-center cohorts and, where feasible, non-diseased reference tissues (e.g., from reduction mammoplasties with confirmed pathological normality) would help to further contextualize these findings.
Conclusions
5
This study suggests the evidence of circ_WHSC1/miR-145-5p interaction in breast cancer. This axis is a potential molecular axis contributing to the pathogenesis of breast cancer. Additional research is needed to validate the functional role of this axis in breast cancer and design novel therapeutic options that target this axis.
Ethics approval and consent to participant
All procedures were in accordance with the ethical standards of the institutional research committee and with the 1964 Helsinki declaration and its later amendments. Informed consent forms were obtained from all study participants. The study protocol was approved by the ethical committee of Tehran University of Medical Sciences (IR.TUMS.MEDICINE.REC.1401.272).
Consent of publication
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Authors’ contributions
AS was the supervisor of the study. AH, NN, and RO provided the clinical samples and related information. LS and MAh analyzed the data. MAb conducted the experiments. SG-F wrote the manuscript. All authors approved the final draft.
Funding
This study was supported by 10.13039/501100004484Tehran University of Medical Sciences.
Declaration of competing interest
Authors declare no conflict of interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Siegel R.L.Kratzer T.B.Giaquinto A.N.Sung H.Jemal A.Cancer statistics 75202510 Ca 202510.3322/caac.21871 PMC 1174521539817679 · doi ↗ · pubmed ↗
- 2Nolan E.Lindeman G.J.Visvader J.E.Deciphering breast cancer: from biology to the clinic Cell 18620231708172810.1016/j.cell.2023.01.04036931265 · doi ↗ · pubmed ↗
- 3Tang L.Jiang B.Zhu H.Gao T.Zhou Y.Gong F.He R.Xie L.Li Y.The biogenesis and functions of circ RN As and their roles in breast cancer Front. Oncol.11202160598810.3389/fonc.2021.605988 PMC 794767233718157 · doi ↗ · pubmed ↗
- 4Chen G.Li Y.Cao J.Dai Y.Cong Y.Qiao G.Construction and analysis of a survival-associated competing endogenous RNA network in breast cancer Front. Surg.92022102119510.3389/fsurg.2022.1021195 PMC 985274536684328 · doi ↗ · pubmed ↗
- 5Mao S.Cheng Y.Huang Y.Xiong H.Gong C.Comprehensive analysis of the exosomal circ RNA-mi RNA-m RNA network in breast cancer J. Gene Med.252023 e 350010.1002/jgm.350036942488 · doi ↗ · pubmed ↗
- 6Chakkaravarthi K.Ramesh R.Palaniyandi T.Baskar G.Viswanathan S.Wahab M.R.A.Surendran H.Ravi M.Sivaji A.Prospectives of Mirna gene signaling pathway in triple-negative breast cancer Pathol. Res. Pract.248202315465810.1016/j.prp.2023.15465837421840 · doi ↗ · pubmed ↗
- 7Davey M.G.Abbas R.Kerin E.P.Casey M.C.Mc Guire A.Waldron R.M.Heneghan H.M.Newell J.Mc Dermott A.M.Keane M.M.Lowery A.J.Miller N.Kerin M.J.Circulating micro RN As can predict chemotherapy-induced toxicities in patients being treated for primary breast cancer Breast Cancer Res. Treat.2022023738110.1007/s 10549-023-07033-837540289 PMC 10504160 · doi ↗ · pubmed ↗
- 8Hajibabaei S.Sotoodehnejadnematalahi F.Nafissi N.Zeinali S.Azizi M.Aberrant promoter hypermethylation of mi R-335 and mi R-145 is involved in breast cancer PD-L 1 overexpression Sci. Rep.132023100310.1038/s 41598-023-27415-836653507 PMC 9849328 · doi ↗ · pubmed ↗