Network Component Analysis Can Identify Potential Axenisation Strategies Circumventing Antibiotic‐Use for Phototrophic Eukaryotic Microalgae
A. Iyer, M. Monissen, Q. Teo, O. Modin, R. Halim

TL;DR
This paper proposes antibiotic-free methods for creating pure microalgal cultures using network analysis of existing techniques.
Contribution
It introduces tailored, antibiotic-free axenisation workflows for diatoms, dinoflagellates, and green algae using network component analysis.
Findings
Filtration, washing, and micropicking are effective for diatoms without antibiotics.
Micropicking, subculturing, and flow cytometry work well for dinoflagellates.
Microscopy, cell counting, and sequencing are essential for verifying culture purity.
Abstract
Axenisation of phototrophic eukaryotic microalgae has been studied for over a century, with antibiotics commonly employed to achieve axenic cultures. However, this approach often yields inconsistent outcomes and contributes to the emergence of antibiotic‐resistant microbes. A comprehensive analysis of previous reports on axenisation was necessary to identify alternate workflows tailored to each major microalgal group. Literature from scholarly databases was systematically recovered and network component analysis was performed to identify method‐clusters commonly reported for the axenisation of diatoms, dinoflagellates, and green algae. Promising workflows circumventing the use of antibiotics appeared to be filtration ↔ washing ↔ micropicking for diatoms, and micropicking ↔ subculturing ↔ flow cytometry for dinoflagellates. No clear workflow could emerge for green algae although Streak…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
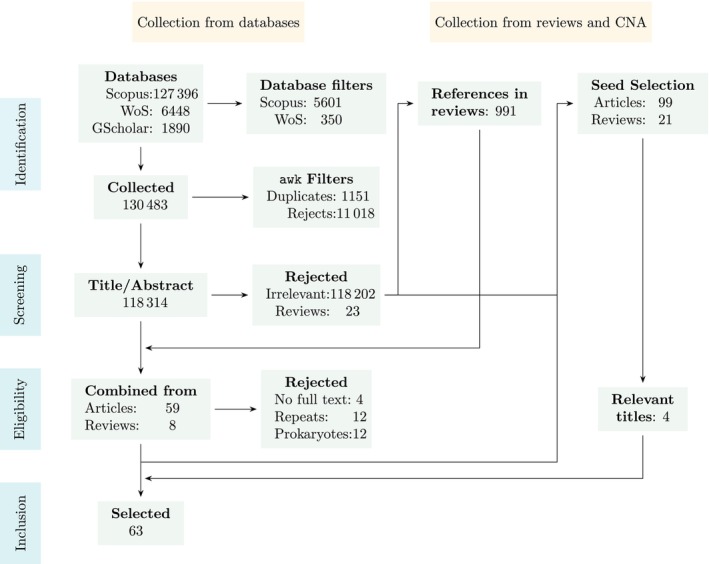 FIGURE 1
FIGURE 1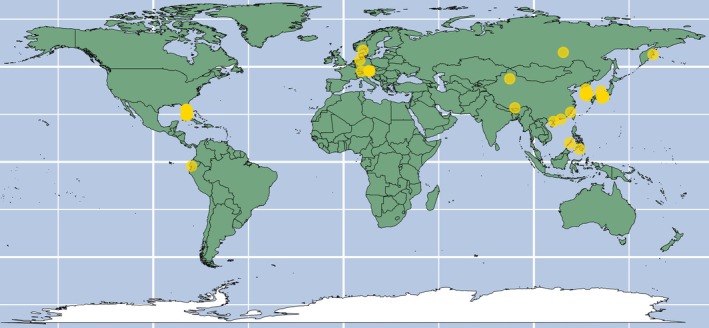 FIGURE 2
FIGURE 2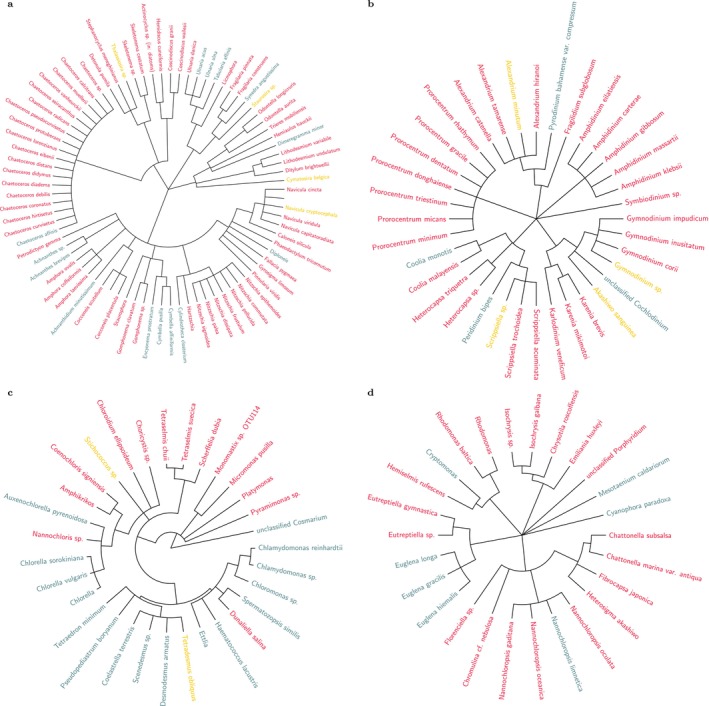 FIGURE 3
FIGURE 3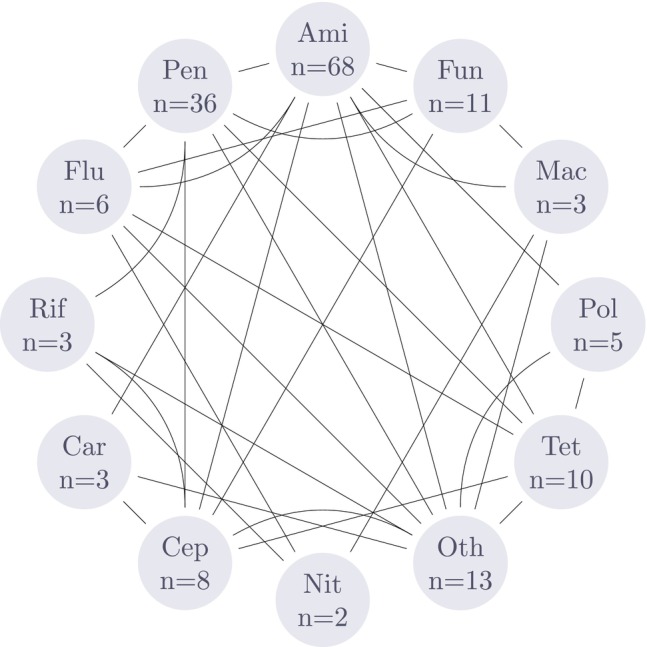 FIGURE 4
FIGURE 4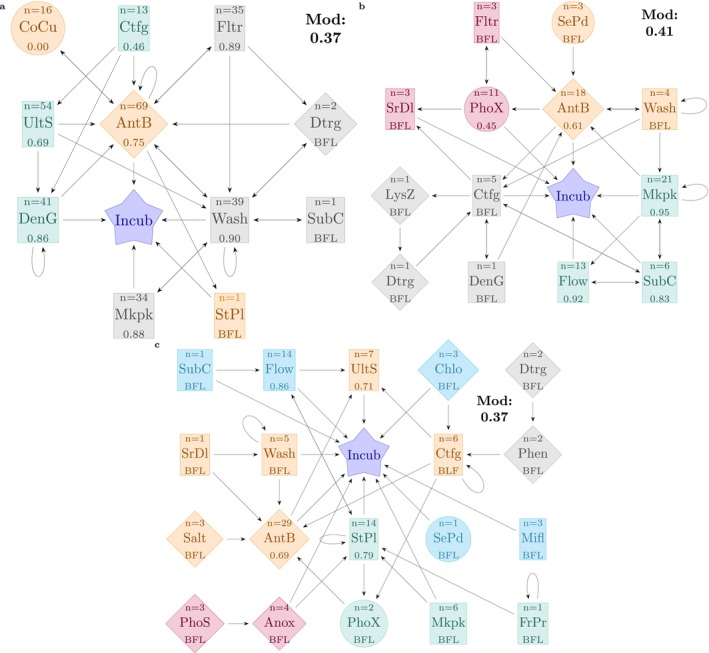 FIGURE 5
FIGURE 5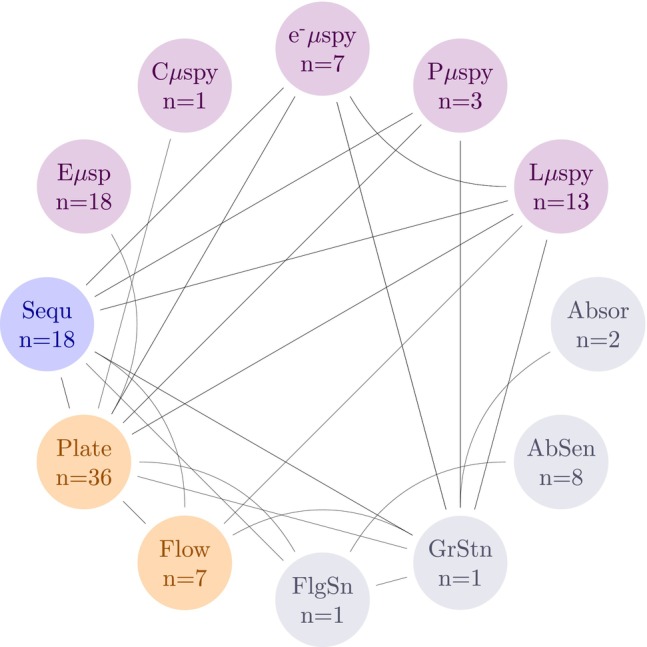 FIGURE 6
FIGURE 6| Division | Method type | Genera | Media | Country | Success |
|---|---|---|---|---|---|
| Diatoms (Pubs = 11) | Biological = 1, Chemical = 1, Physical = 8 |
| Bacillariophycean medium ( | China ( | Yes = 8, No = 2, Multi/Part = 1 |
| Dinoflagellates (Pubs = 15) | Biological = 2, Chemical = 3, Physical = 8 |
| Artificial seawater ( | China ( | Yes = 7, No = 5, Multi/Part = 3 |
| Green algae (Pubs = 33) | Biological = 2, Chemical = 6, Physical = 10 |
| Artificial seawater ( | Austria ( | Yes = 22, No = 7, Multi/Part = 3 |
| Other eukaryotes (Pubs = 22) | Biological = 5, Chemical = 24, Physical = 24 |
| Artificial seawater ( | Canada ( | Yes = 15, No = 5, Multi/Part = 2 |
| Class | Diatoms | Dinoflagellates | Green algae | Others |
|---|---|---|---|---|
| Aminoglycosides | 135 (10–600) | 160 (8–2000) | 600 (8–10,000) | 600 (8–2000) |
| Carbapenems | 98 (98–98) | 10 (10–10) | 5 (5–5) | — |
| Fluoroquinolones | 5 (5–5) | 36.5 (23–50) | 13 (10–16) | — |
| Penicillins | 250 (50–600) | 100 (50–1000) | 600 (2.5–2000) | 100 (10–2000) |
| Tetracyclines | 10 (10–10) | 12 (1–12) | 50 (5–500) | 50 (12–500) |
| Cephalosporins | — | 60 (20–100) | 400 (10–500) | 30 (30–400) |
| Macrolides | — | 6 (6–6) | 54 (8–100) | 6 (6–6) |
| Polypeptides | — | 14 (12–16) | 3.3 (3.3–5000) | 16 (3.3–30) |
| Rifamycins | — | 10 (10–10) | — | 30 (25–30) |
| Nitrofurans | — | — | 10 (10–10) | 250 (250–250) |
| Antifungal Agents | — | 5 (5–15) | 20 (0.01–400) | 15 (1–15) |
| Others | 27 (27–27) | 125 (50–200) | 35 (5–60) | 20 (2.5–68) |
| Division | Cluster | Nodes | Density | Degree | Cliques |
|---|---|---|---|---|---|
| Diatoms | Overall | Antibiotics Co Culture, Streak plating, Filtration, Surfactants, Washing, Subculturing, Micropicking, Ultrasonication, Density gradient, Centrifugation | 0.21 | 4 |
1. Ultrasonication, Antibiotics, Centrifugation, Density gradient. 2. Antibiotics, Filtration, Washing, Surfactants. |
| Cluster 1 | Streak plating, Antibiotics, and Co Culture | 0.44 | 2 |
1. Streak plating, Antibiotics. 2. Antibiotics, Co Culture. | |
| Cluster 2 | Filtration, Surfactants, Washing, Subculturing, Micropicking | 0.36 | 2 | Filtration, Surfactants, Washing | |
| Cluster 3 | Ultrasonication, Density gradient, Centrifugation | 0.44 | 2 | Ultrasonication, Density gradient, Centrifugation | |
| Dinoflagellates | Overall | Selective predation, Antibiotics, Washing, Micropicking, Subculturing, Flow Cytometry, Centrifugation, Lysozyme, Surfactants, Density gradient, Filtration, Phototaxis, Serial dilution | 0.17 | 4.5 | Subculturing, Flow cytometry, Micropicking |
| Cluster 1 | Selective predation, Antibiotics, Washing | 0.44 | 3 |
1. Selective predation, Antibiotics. 2. Antibiotics, Washing. | |
| Cluster 2 | Micropicking, Subculturing, Flow cytometry | 0.67 | 4 | Micropicking, Subculturing, Flow cytometry | |
| Cluster 3 | Centrifugation, Lysozyme, Surfactants, Density gradient | 0.31 | 2 | Centrifugation, Lysozyme, Surfactants | |
| Cluster 4 | Filtration, Phototaxis, Serial dilution | 0.33 | 2 |
1. Serial dilution, Phototaxis. 2. Phototaxis, Filtration. | |
| Green algae | Overall | Anoxy, Photosensitisation, Antibiotics, Salt solution, Ultrasonication, Washing, Centrifugation, Serial dilution, Streak plating, Phototaxis, Micropicking, French Press. | 0.11 | 2.5 | Centrifugation, Antibiotics, Phototaxis, Streak plating |
| Cluster 1 | Anoxy, Photosensitisation | 0.5 | 2 | Anoxy, Photosensitisation | |
| Cluster 2 | Antibiotics, Salt solution, Ultrasonication, Washing, Centrifugation, Serial dilution | 0.33 | 4 |
1. Serial dilution, Antibiotics, Washing. 2. Antibiotics, Centrifugation, Ultrasonication. 3. Antibiotics, Centrifugation, Washing. | |
| Cluster 3 | Streak plating, Phototaxis, Micropicking, French Press | 0.37 | 2.5 |
1. Micropicking, Streak plating. 2. Phototaxis, Streak plating. 3. Streak plating, French press. | |
| Cluster 4 | Flow Cytometry, Predation, Subculturing, Hypochlorite, Microfluidics | 0.04 | 0 | Flow cytometry, Subculturing | |
| Cluster 5 | Surfactants, Phenol | 0.25 | 1 | Surfactants, Phenol |
| Limitations | Consequence | Mitigation |
|---|---|---|
|
| Many divisions such as cyanobacteria were excluded from the study even though they are an important member of the microalgae umbrella, and are of significant scientific and industrial relevance. | The workflow presented here, particularly the division‐wise approach to study the workflows, is modular and can be extended to other organisms. Further work can build upon the existing code base ( |
|
| Intrinsic success rates of each method involved in the axenisation workflow were not calculated and consequently, the success rates of known and potential hypothetical workflows cannot be calculated. | More high‐quality experimental evidence of under‐reported methods is required to ascertain success rates of individual methods and their combinations to construct robust efficacy models of the workflow networks. |
|
| Comparisons between culture banks on important techniques, method workflows, and caveats when working with certain microalgae were missed. It remains unclear which microalgae have formed obligate symbiotic relations with bacteria and if it is possible to supplement the necessary growth factors to grow the microalgae in an artificial axenic culture. | National culture banks can be systematically identified and a questionnaire can be designed to identify workflows, methods, and important caveats when working with certain microalgae. |
- —Irish Research Council10.13039/501100002081
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlgal biology and biofuel production · Innovative Microfluidic and Catalytic Techniques Innovation · Diatoms and Algae Research
Introduction
1
Axenicity ‘describes the state of a culture in which only a single species, variety, or strain of organism is present and entirely free of all other contaminating organisms’ (Thain and Hickman 2004). Popular culture banks host an impressive collection of microalgal species, but most of the offered type samples are xenic, that is, they contain bacteria, amoebae, or fungi that grow in consort with microalgae in the natural habitat from where they are collected. Although characterising these commensal organisms provides important clues about the community where microalgae thrive, limiting the influence of confounders is crucial in experimental designs. For example, Deng et al. (2024) had to achieve and verify the axenicity of their Coscinodiscus radiatus cultures to unambiguously assess the modulatory effects of the bacteria Mameliella sp. CS4 and Marinobacter sp. CS1 on microalgal ageing and extracellular vesicle production. Robust investigation in microalgal research requires a supply of pure cultures, but despite a century of reportage on axenisation procedures no consensus or unified workflow has been described that can reliably generate pure cultures. Contaminating microbes may be present in the extracellular (e.g., as phycospheric bacteria, attached symbionts) as well as intracellular environments (e.g., as intracellular symbionts) and consequently, a harmonised protocol for axenisation becomes difficult even for the same species. For example, Brown (1982) and Su et al. (2007) employed physical methods such as centrifugation and filtration and reported contrasting axenisation outcomes for Eutreptiella sp. However, Kuo and Lin (2013) noted the presence of Alphaproteobacteria and Marinobacter in the phycoshperic as well as intracellular environments, and employed an antibiotic cocktail with lysozyme to achieve contaminant‐free cultures. Currently, the onus of achieving, maintaining and verifying axenicity of mother cultures used in experiments falls on the researchers rather than the supplier/culture collection. Consequently, interpreting the results from publications emanating from different research groups requires caution as the final outcome may have been significantly influenced by the co‐cultured microbes.
Previously, Fernandez‐Valenzuela et al. (2021) classified the methods based on their mode of action and provided a narrative summary on their application to generate axenic microalgal cultures. For example, methods involving direct physical manipulation of a cell or a group of cells such as in micropicking, filtration, centrifugation, etc. were deemed as physical methods. Physical methods tend to be labour‐intensive, require training and heavy initial investment on instrumentation. However, they tend to be reliable in their outcomes, can be automated to a large extent and are more sustainable as they do not employ the use of chemicals or generate significant effluents. Chemical methods use compounds to kill or inhibit microbial growth (e.g., antibiotics, chlorate, etc.) and have the advantage of being highly accessible and easy to use. Antibiotics are by far the most commonly used method in axenisation workflows although this has begun to raise problems around the emergence of antibiotic resistance (Iyer 2025). Lastly, harnessing specific physiological traits unique to the microalgal division or species to help separate them from contaminants was deemed as biological methods such as phototaxis or selective predation. These methods are often unique or highly tailored for a species, making their use rather restricted. However, biological methods often double as identity markers as well; particularly when working in multi‐algal systems. In fact, Fernandez‐Valenzuela et al. (2021) highlight the importance of a tailored approach for each species and highlighted the fragmented and inconsistent reportage of the methodology in the available literature. However, details on literature collection and analyses were not described in their review. Thus, the work presented herein attempts to systematically collect and collate all the available resources, extract the relevant information from the described methods and perform network component analysis to understand overarching trends in axenisation workflows and address the research question: ‘Which axenisation workflow(s) is/are best suited for a given microalgal division’. The secondary aim was to identify axenisation workflows circumventing the use of antibiotics.
The approach used in this work involves Network component analysis (NCA) to categorise the methods used within axenisation workflows into nodes connected in a network, and then infer causal relationships by comparing the sequence and combinations in which methods are applied. NCA is a powerful tool for visualising and identifying multiple ‘sub‐networks’ or ‘alternate pathways’ that can tailor workflows to be compatible with secondary considerations such as method success rates or access to instrumentation, etc. Furthermore, the cluster and clique identification analyses can help stratify methods based on emergent network properties. For example, Fernandez‐Valenzuela et al. (2021) manually classified and stratified methods such as centrifugation or washing as ‘intermediate’ steps and antibiotics and micropicking as ‘main’ steps in an axenisation workflow based on their mechanism and mode of action. NCA, however, is agnostic to a methods' mechanism and rather looks for clusters based on the nature of its connections to other methods in the network.
Lastly, the results from this work are contextualised in real‐world experiences and expectations by noting the comments from four culture banks on the challenges faced when attempting to generate and maintain axenic microalgal cultures.
Materials and Methods
2
Search Protocol
2.1
The search protocol used a hybrid method of structured and systematic querying using bibliometric databases, followed by citation network analyses to discover and collect publications. Search was carried out in two phases. In the first phase, the search term (algae AND (axen* OR purif*)) was queried in Web of Science, Google Scholar, and Scopus bibliometric databases and filtered based on title and abstract using an awk script. The publications were then subjected to citation network analysis using the online tool Local Citation Network (https://localcitationnetwork.github.io/).
The search terms were further expanded to ensure that all publications were truly captured in the first round. The search terms followed the module of <synonyms for microalgae> AND < microalgal divisions> AND < synonyms for axenisation> and expressed as:(Microalg* OR phytoplankton OR eukaryotic microalg*)AND(chlorophy* OR bacillariophyceae OR diatom OR haptophyt* OR crypto* OR dino* OR chrysophyt* OR eugleno* OR rhodophyt* OR prasinophyte OR coccolithophorid)AND(axen* OR purif* OR decontam* OR aseptic OR steril* OR gnotobio* OR unialg* OR monoalg* OR “bacteria‐free” OR “microbe‐free”)
Publications in all languages were included and relevant information was extracted from the text using online translation tools. The working definition of microalgae for the purposes of this review was organisms that are single‐cellular, phototrophic, eukaryotic, free‐living, and capable of asexual reproduction. The exhaustive list of parameters, exclusion criteria, and rationale is available in Table S1.
Data Items and Synthesis
2.2
The following information was extracted from the literature and is provided in a master table as a .csv file (Table S2):
- Year of publication
- Country of the research institution(s)
- Microalgal species: The species, including the culture code/accession number if provided by the author.
- Purification method(s): The method described by the publication to isolate microalgae or decrease the contaminant load. The order in which the methods are written represents the order in which they were applied in the described experimental procedure.
- Screening method(s): Methods used to verify if the method used to achieve axenicity was successful. Alternatively, this also refers to methods to quantify the contaminant load and check the degree of success of the treatment.
- Media used: The media used to grow the microalgae. In cases where the plates are used to test axenicity, the media composition of the agar plates was noted here as well.
- Screening method(s): Methods used to verify axenicity. Alternatively, this also refers to methods to quantify the contaminant load.
- Quality score: Refer to the subsection 2.3 below.
Data extraction procedures are detailed further in Supporting Information: Text, Section S1.4.
The various axenisation methods noted in the publications were classified either as Physical, Chemical, and Biological, where Physical methods involved the use of techniques such as colony picking to isolate a group of axenic cells from a Petri‐dish for further culturing, or density gradient centrifugation to separate microalgae based on relative cellular densities. Chemical methods referred to the use of antibiotics or special compounds that inhibit the growth of unwanted microbes to achieve axenic microalgal cultures. Biological methods included methods that leveraged the unique abilities of the microalgae to isolate them from bacteria and fungi, for example, the use of phototaxis to isolate microalgae (Imai and Yamaguchi 1994), or removing the carbon source and depleting the nitrogen content of the medium, to give the microalgae a competitive advantage. Another example includes the addition of specific microbes that overgrow or inhibit the existing contaminating microbes, and then subsequently removing the introduced microbe using antibiotics or through physical methods.
Evidence Quality
2.3
Using the approach described by Jadad et al. (1996), five questions were framed based upon which the experimental quality of each publication describing the axenisation procedure was scored by AI, MM and QT. The final score was the rounded average of the three scores. The questions are noted below:
- Does the publication report the source of the organism(s)?Yes: 1, No: 0 Explanation: The contaminating organism(s) may change based on the location from where the microalgae was sampled. In the case of purchase from culture collections, in‐house processing protocols, long‐term storage (such as cryopreservation), and handling may alter the composition or presence of other organisms cultured along with the microalgae.
- Are technical details of the axenisation method explained to facilitate reproducibility?Yes: 1, Incompletely: 0, No: −1 Explanation: The question pertains to whether sufficient details of the chemicals or components used in the axenisation process are provided to easily procure or purchase them, or accurately construct, assemble, or fabricate them.
- Was growth medium clearly described?Yes: 1, Referenced: 0, No (either not mentioned, or only mentioned without proper reference): −1 Explanation: The growth medium works in synergy with the purification to support growth of the microalgae which may be stressed after any purification step. Corresponding changes made to the growth media would help ensure successful regeneration of the isolated cultures. Besides, numerous variants of media have been described without labelling or classifying them as an entirely different composition mix.
- Was axenicity verified after treatment?With multiple methods: 1, With only one method: 0, No: −1 Explanation: Veracity of the experimental purification method success validates treatment efficacy.
- Were contaminating organisms identified?Yes: 1, No: 0 Explanation: A method's efficacy may change depending on the nature of the species contaminating the microalgal culture. Characterisation or the identification of the contaminating microbes would help establish relevance to the purification method employed.
Data Visualisation and Availability
2.4
Data were processed using R (4.4.0) (R Core Team 2024) and preliminary analyses were performed using the tidyverse package (Wickham et al. 2019). Phylogenetic trees from the species names were constructed using the R packages metacoder (Foster et al. 2017), ggtree (G. Yu et al. 2017), taxize (Chamberlain and Szöcs 2013), and rentrez (Winter 2017). Further network component analysis was performed using the iGraph (Csárdi et al. 2024) and tidygraph (Pendersen 2024) packages. Subsequent visualisation of the network and the phylogenic tree was performed using the TikZ (Tanau 2021) and pgfplots (Feuersänger 2023) packages in LaTeX (Lamport 1994).
For the network analysis, two main outcomes were noted: first, the success rates for each method (number of times axenisation was successfully achieved/number of times the method was used), and second, the grouping/clustering of methods in a network for each microalgal division. To calculate success rates, only those methods that were reported in more than six species were included to remove over‐representation of a single method or species within the division level analysis. Next, the clusters within each network were identified using ‘optimised modularity algorithms’ (see documentation of iGraph) and the largest clique (methods commonly used together across the publications) was then obtained.
The codes used to process the information are presented in Supplementary_codes.pdf. The awk code used to filter additional irrelevant titles is provided in Supporting Information: Text, Section S2. All data and resources necessary to reproduce the analyses are available in the OSF repository: https://osf.io/8cmk7/.
Contact With Culture Collections
2.5
The culture collections of CCAP, Bigelow, CSIRO, and NIES were contacted to enquire about the number of unique microalgal species they house, and how many of them are available only in xenic forms, how many are available as axenic as well as xenic. In addition, a subjective question was asked on the challenges they face when trying to make some strains axenic. Their answers are quoted and added to the arguments made in the Discussion section.
Results
3
Summarisation of Data
3.1
The systematic search yielded 63 publications from which data was extracted and analysed as shown in Figure 1. Only four publications lacked any DOIs or similar permanent identifiers. Among the selected publications, 34 reported only one microalgae while the other 29 reported multiple microalgal species. The search query using databases yielded 52 relevant articles, and an additional 11 articles were recovered using citation network analysis. Thus, lateral search methods could contribute to 17.7% of the total recovered relevant publications. A summary of all the information collected from 63 publications is provided in Table 1. A consolidated master table with the greater details of each study is provided in Table S2.
PRISMA‐Sc flow schema of the first search strategy and the recovered publications included herein. CNA = citation network analysis that was performed using the GitHub tool to discover additional publications. Across the search workflow, 52 relevant articles using the traditional search method were recovered, with an additional 11 using citation network analysis. Thus lateral search methods could contribute to 17.7% of the total recovered relevant publications. Gscholar = Google Scholar; WoS = Web of Science.
Across the publications, there were 24 unique axenisation methods that were broadly classified as ‘Biological’ comprising bacterial co‐culturing (CoCu), selective predation (SePd), and phototaxis (PhoX), ‘Chemical’ comprising antibiotics (AntB), lysozyme (LysZ), hypochlorite (Chlo), surfactants (Dtrg), phenol (Phen), anoxy (Anox), dye‐based photosensitisation (PhoS), and salt solution (Salt), and ‘Physical’ comprising centrifugation (Ctfg), streak plating (StPl), washing (Wash), micropicking (Mkpk), filtration (Fltr), density gradient (DenG), ultrasonication (UltS), subculturing (SubC), microfluidics (Mifl), french press (FrPr), serial dilution (SrDl), and adsorptive resin (Resn). A brief description of these methods is provided in Supporting Information: Text S1, Sections S1.1–S1.3. The six most commonly used methods grouped according to the report quality and axenicity outcome are shown in Figure S1 and the comprehensive counts of methods used for each species grouped according to axenicity outcome are provided in Table S3.
Reported Species and Sampling Locations
3.2
Across the literature, 174 unique species were identified and grouped into non‐taxonomic divisions such as diatoms, dinoflagellates, etc. and further classified as freshwater, marine, or brackish species. Microalgae purified from soil were also found to grow in freshwater and were categorised accordingly. The recorded divisions included cryptomonads (n = 4), diatoms (n = 81), dinoflagellates (n = 36), euglenoids (n = 5), golden algae (n = 1), eukaryotes (n = 10), green algae (n = 31), haptophytes (n = 4), red algae (n = 1), and green plants (n = 1). Although diatoms comprised the largest number of reported species, they were generally subjected to only 11 of the total 24 methods. Conversely, all 24 methods were applied to the green algae division. Many of the reported organisms such as Nannochloris sp., and Chlorella sp., have significant commercial value and thus the investigation of newer methods to rapidly and effectively achieve axenic cultures is incentivised
Only 19 publications documented the collection of samples from natural water sources (e.g., lakes, rivers, and coastlines) for the isolation and identification of microalgae. Furthermore, except for Aray‐Andrade et al. (2018), who sampled along the Ecuadorian coastline, most reported sample collections from natural sources were confined to the Northern Hemisphere. Another 27 publications reported their experiments using cultures from culture banks (public or private), while 17 publications provided no information on where the microalgal samples were obtained.
The geographical sites where microalgal sampling were reported are shown in Figure 2, and the phylogenetic trees of the reported species are presented in Figure 3a–d.
Geographic locations of microalgal samples collected from locations noted in 19 publications.
Phylogenetic tree of the species noted in the collected publications. Species names are coloured red for marine, sunset‐yellow for brackish, and steelblue for freshwater species. (a) Taxonomic tree of 81 diatom species identified across 11 publications. (b) Taxonomic tree of 36 dinoflagellate species identified across 15 publications. (c) Taxonomic tree of 31 green algae species identified across 33 publications. (d) Taxonomic tree of 26 microalgal algae species identified across 22 publications.
Antibiotics—The Most Popular Treatment
3.3
Antibiotics were by far the most preferred approach to generate axenic cultures as they are convenient to apply and the outcomes were generally rapid. Across the 63 publications, 42 reported the use of antibiotics spanning across 12 classes as shown in Table 2. In almost all cases, these antibiotics were applied as a cocktail and the combinations are shown in Figure 4. As the number of components in a cocktail increases, the range and potency to kill contaminants rose, albeit at the risk of affecting the viability of the microalgae.
Antibiotic class combinations reported in the literature. Ami = aminoglycosides; Car = carbapenems; Cep = cephalosporins; Flu = fluoroquinolones; Fun = antifungal agents; Mac = Macrolides; n = number of publications reporting the use of the antibiotic class; Nit = nitrofurans; Oth = others; Pen = penicillins; Pol = polypeptides; Rif = rifamycins; Tet = tetracyclines.
Diatoms were exposed to the fewest classes of antibiotics; predominantly Aminoglycosides and Penicillins, while dinoflagellates and green algae appeared to have been experimented upon with more classes of antibiotics. The three most commonly employed antibiotics included two aminoglycosides; namely, streptomycin (n = 21; median conc. = 100 mg L^−1^), kanamycin (n = 17; median conc. = 200 mg L^−1^) and one penicillin; namely, ampicillin (n = 16; median conc. = 100 mg L^−1^). neomycin + kanamycin (n = 7), streptomycin + penicillin‐G (n = 7) and streptomycin and neomycin (n = 6) were the most common cocktail pairs employed in the publications.
Antibiotics require dosing depending on axenisation strategy. For example, Kan and Pan (2010) and Han et al. (2016) profess acute exposure of the culture to antibiotic cocktails to achieve axenicity, while Youn and Sung‐Bum (2007) and Wang et al. (2016) employ antibiotics at moderate to low concentrations to achieve axenic cultures over multiple rounds of treatment to achieve and maintain axenicity.
Alternate Axenisation Pathways With Network Component Analyses
3.4
Workflow networks for three of the most commonly reported divisions, namely, diatoms, dinoflagellates, and green algae are provided in Figure 5a–c respectively. Workflow networks for microalgal families with low representation grouped under ‘others’ are available in Figure S2.
Clustering of the axenisation workflows. In each panel, methods belonging to the same cluster are assigned the same colour. Panel A shows the workflows incorporating the 11 methods used for 81 diatoms species in 11 publications. Panel B shows workflows incorporating 13 methods for 36 dinoflagellate species in 15 publications. Panel C shows the workflows incorporating 19 methods for 26 green algae species in 22 publications. Numbers in the network components represent success rates when number of reports is n > 6; for n ≤ 6 reports, success rates are below the filter limit (BFL) and not calculated. Node shapes denote method types: Squares for ‘Physical’, circles for ‘Biological’, and diamonds for ‘Chemical’. Nodes of the same colour indicate they belong to the same cluster. The analysis is presented at the species level. Mod value represents the modularity value (range between 0 = no clustering to 1 = perfect clustering) of the nodes in the network space.
Analyses included identifying clusters and cliques between the methods employed. ‘Clusters’ represent the collection of nodes (here, methods) with similar network characteristics such as the edge density (number of connections to other nodes), and relative position and separation from the central node—Incubation (i.e., if they are directly connected to incubation or first connect to other nodes/methods before reaching incubation; see iGraph documentation by Csárdi et al. (2024) and more general works by Clauset et al. (2004) for deeper analysis). ‘Cliques’ on the other hand, comprise adjacent nodes that are all directly interconnected (i.e., methods that are often used together in immediate succession) (Eppstein et al. 2010).
Network component analysis was applied to identify the largest clique of methods within a cluster so that methods with similar network properties could be used together. For example in Figure 5a, although filtration (Fltr), surfactants (Dtrg), washing (Wash), subculturing (SubC), and micropicking (Mkpk) clustered together (cluster 2, marked in grey) owing to similar network properties, the largest clique of methods was between Filtration ↔ Surfactants ↔ Washing. Details of the analyses and codes for Figure 5, are provided in Supporting Information: Codes (Code Chunk 6).
In the case of green algae (Figure 5a and Table 3), the overall network density was 0.11 (ratio of actual number of connections to the theoretical maximum number of connections) while in comparison, diatoms have an overall network density of 0.21. Considering that 19 different methods have been reported on green algae across 22 publications, while only 11 methods were reported on diatoms across 11 publications, it appears that authors have been more experimental with the axenisation of green algae, with a disperse reportage on a wider method collection as opposed to diatoms where the method set is relatively limited and reported more frequently across the publications.
Details of the overall network, and the clusters and cliques for diatoms, dinoflagellates, and green algae are summarised in Table 3. Identification of clusters and cliques for other eukaryotic microalgae (Figure S2) was not performed as they did not represent any specific division/grouping.
Appraising Publications and Confirming Axenicity
3.5
A point‐based appraisal mechanism was devised to assess publications describing experimental axenisation procedures for microalgae. Based on the questions noted in Section 2.3, about 7 out of 63 publications (11%) had a score < 0. The timeline of publications and the distribution of the publication quality within each time‐period is provided in Figure S3.
Poor method description (addressing Question 2) and lack of post treatment verification of axenicity (addressing Question 4) were the most common reasons for low scores. For example, across the 63 publications, six publications did not report a follow‐up verification (consequently scoring −1 for Question 2), 23 publications reported the use of only one verification method (scoring 0 for Question 2). Only 12 publications used a combination of two verification methods, while the remaining 22 employed three or more methods. A connection graph of the usage and combinations of verifications noted in literature is shown in Figure 6.
Methods reported in literature used either individually, or in combination to assess axenicity in microalgal cultures. Resolution of information is at publication level. AbSen = Antibiotic sensitivity; Absor = absorbance; Cμspy = confocal microscopy; e‐μ spy = electron microscopy; Eμspy = epifluorescence microscopy; FlgSn = flagella staining; Flow = flow cytometry; GrStn = gram staining; Lμspy = light microscopy; Pμspy = phase contrast microscopy; Plate = plate counting; Sequ = 16S/18S sequencing. Methods marked in purple are microscopy‐based methods. Methods marked in orange are cell counting methods. Methods marked in blue are sequencing methods. Methods marked in grey are physiology specific methods.
Across the publications, 13 distinct methods were reported to check axenicity of the final culture which could be grouped based on microscopy, cell/colony counts, sequencing, and physiology. Collectively, microscopy techniques that included confocal, electron, epifluorescence, phase contrast, and standard light microscopy were the most popular approach to verify axenicity (n = 42). Agar plate counting (n = 36) was the next most common technique. Sequencing (both 16S/18S) methods were used in 18 publications, and lastly methods leveraging specific aspects of the microalgal physiology such as light absorbance, flagella staining, etc. were collectively reported in 12 publications.
Discussion
4
Axenisation of microalgal cultures remains a topic of significant research interest and can become necessary or undesirable depending on the research question being pursued. For example, Li et al. (2022) profiled the bacterial consortium at each life stage of Haematococcus pluvialis and posited that the presence of bacteria enhanced biomass production and growth rate without any real threat of a culture crash, while Yu et al. (2022) contradict this by reviewing the strategies to purify the microalgae and noted the risks posed to a cultivation batch under different nutrient conditions when contamination was unchecked, and highlighted the importance of maintaining axenic and sterile cultivation conditions to achieve optimal biomass and astaxanthin production.
On a wider note, there is a resurgence of interest in community interactions between microalgae and other organisms (Pushpakumara et al. 2023), either as interactions in non‐associative biological mixtures in an experimental system (Grossart 1999; Solomon et al. 2023), or indeed in symbiotic periphyton (Francoeur et al. 2003), and subsequently their emergent metabolic properties in relation to the wider ecology. Consequently, teasing out the individual contribution from a group effect, or attributing metabolic properties to an organism remains challenging (Fouilland et al. 2018). Although the publication by Dextro et al. (2024) focuses on cyanobacteria, the arguments noted in the review about the desirability (or lack thereof) of axenicity in microalgal cultures translate to phototrophic eukaryotic microalgal cultures as well, and their conclusions appear to suggest that axenic cultures are suitable for genomic, proteomic, and industrial applications, while non‐axenic cultures that preserve the natural bacterial consortium are necessary for fundamental research on understanding ecological dynamics between these microbes.
Many high quality and detailed reviews such as those by Fernandez‐Valenzuela et al. (2021), Molina‐Cárdenas et al. (2016), Vu et al. (2018), and McCracken (1969), have previously described axenisation procedures for microalgae. While these publications provide critical insights in applying the investigated methods, their approach was narrative and a top‐down perspective of the various strategies adopted as a combination of axenisation methods has not been presented thus far. The approach adopted in this study was to collect methodologies described in publications and visualise them as ‘networks’ of the methods connected to each other depending on the sequence in which they were applied that ultimately led to the final step of ‘incubation’ where the culture is presumed to be axenic. This allows for methods to cluster within the network based on key characteristics such as edge density and relative node position. Edge density refers to the number of connections a node shares with others, influencing how tightly integrated a method is within the network. Another important factor is the relative position of a method relative to ‘Incubation’ which is the final step in an axenisation workflow and the central node in the network. This position determines whether a methodology is single step (i.e., only method directly connected to Incubation) or sequential (many methods used in sequence) before generating an axenic culture.
The methods described in the publications were grouped according to their mode of action into Physical, Chemical and Biological; with each presenting their characteristic advantages and shortcomings that need to be considered when designing an axenisation workflow. Physical methods require the most initial investment in the form of equipment as well as manual skill to successfully decontaminate the culture. By themselves, physical methods such as density gradient, washing, or filtration do not generate axenic cultures but can contribute to the wider process to achieving them. Micropicking techniques require specialised skills to carefully separate microalgal cells from the contaminating organisms without damaging cells. Flowcytometry, on the other hand, can automate the precise sorting process but is generally ineffective against physical aggregates of microalgal cells and bacteria.
Chemical methods, on the other hand, are the most popular, though the use of antibiotics. But as noted before, this has led to the wider problem of generating antibiotic‐resistant bacteria/fungi, while also adversely affecting the growth of the microalgal cells. In some cases, microalgae are known to metabolise the antibiotics and thus, antibiotic application may inadvertently become ineffective (Frascaroli et al. 2024). Besides, chemical methods such as the use of detergents or oxidising agents such as chlorate can significantly hamper the viability of the culture, and incessant use can become mutagenic.
Lastly, biological methods have not been widely reported across the literature owing to the highly specific nature of their mode of action. Phototaxis is not a global microalgal ability and is mostly restricted to certain dinoflagellates and green algae. Biological methods offer a more sustainable method of contaminant removal by leveraging physiological properties of the microalgae and selectively promoting their growth at the cost of the contaminants.
Clusters and Cliques From Network Component Analysis
4.1
The network component analysis revealed variations in methodological approaches and interconnections across microalgal divisions, with key differences in network density, clustering, and reliance on specific methods. Workflows described to achieve axenic diatom cultures comprising of 11 nodes (methods) exhibited a moderate network density of 0.21 (ratio of the number of connections to the theoretical maximum number of connections possible) and a median node degree of 4 (i.e., number of connections; incoming and outgoing, for each node), and an evenly distributed number of methods in each cluster. Workflows for dinoflagellates comprising of 13 nodes had a lower network density of 0.17, despite a higher median degree of 4.5, suggesting a broader range of methods applied, but with weaker interconnections. For example, methods such as phototaxis or selective predation were used in limited combinations with other methods. The sparseness in the workflow network is demonstrated further in the methods described for green algae. With 19 nodes, the network density was only 0.11 and had a median degree of 2.5. This indicates that a large number of specialised methods were used, largely in isolation. For example, selective predation and microfluidics were used in isolation without using them in concert with other methods.
Cluster Characteristics
4.1.1
Diatoms
4.1.1.1
Two method‐specific clusters of similar cliques with similar network densities of 0.44 emerged, namely Cluster 1 comprising streak plating, antibiotics (success rate = 0.75), and co‐culture (success rate = 0.0), and Cluster 3 comprising ultrasonication (success rate = 0.69), density gradient (success rate = 0.86), and centrifugation (success rate = 0.46). Cluster 1 uses a mix of physical, chemical, and biological methods to achieve axenic cultures, while Cluster 3 comprises exclusively physical methods and consequently offers a workflow that circumvents the use of antibiotics.
Dinoflagellates
4.1.1.2
Cluster 2 comprised a clique with the highest network density of 0.67, and exclusively involved physical methods of micropicking (success rate = 0.95), subculturing (success rate = 0.83), and flow cytometry (success rate = 0.92). Other clusters of interest were Cluster 3 and Cluster 4 that used methods leveraging the specificity of enzyme action (lysozyme) and motility (phototaxis), but had low network densities and an insufficient number of publications to calculate success rates.
Green Algae
4.1.1.3
Clustering with green algae was challenging as the method diversity was high, but the connections between them were weak owing to the lack of sufficient literature. For instance, Cluster 1 comprising of only anoxy and photosensitisation had the highest network density of 0.5, but none of the methods were sufficiently represented in the literature to calculate a success rate. In comparison, Cluster 2 comprising of a wider collection of methods such as antibiotics (success rate = 0.69), salt solution, ultrasonication (success rate = 0.71), washing, centrifugation, and serial dilution, and an overall network density of 0.33, had two methods with sufficient representation to calculate success rates but involved the use of antibiotics. Streak plating (success rate = 0.79) ↔ flowcytometry (success rate = 0.86) → ultrasonication appears to be the only alternate workflow with sufficient data about their success rates circumventing antibiotic use, although each method appears in a different cluster.
Key Comparisons and Trends
4.1.1.4
Across the workflows described to achieve axenic microalgal cultures, clusters and cliques could be identified that avoided the use of antibiotics. For example, dense clusters for methods like micropicking, subculturing, and flow cytometry indicate that precision techniques could be used to achieve axenic dinoflagellate cultures. Similarly, with diatoms, filtration, washing, and micropicking or ultrasonication and density gradient have been reported to attempt axenisation. In contrast, green algae do not show any preferred approach towards achieving axenisation, although there is preliminary evidence showing the use of anoxy and photosensitisation to be effective.
Across all divisions, antibiotics emerged as a common bridging method, appearing in high‐degree nodes and multiple cliques. Mechanical methods like ultrasonication and centrifugation were consistently clustered in diatoms and green algae, but were less central for dinoflagellates, reflecting divisions‐specific adaptations to their cell structures and ecological traits. Dinoflagellates had the highest reliance on precise isolation methods, likely due to their complex morphology and their own susceptibility to antibiotics and harsh chemical methods. Diatoms showed a balance between mechanical and chemical techniques, leveraging synergies within and across clusters. Green algae utilized a blend of unique (e.g., photosensitisation) and shared methods (e.g., antibiotics), indicating a diverse and disperse network of approaches.
Verification Methods
4.2
Across the recovered literature, six publications did not properly verify the axenicity of their final culture, while eight publications only used one method to declare axenicity. This is important as each verification method has inherent limitations that make it insufficient when used as the sole test. For example, plate counting is a well‐established technique for detecting viable bacteria, as it requires microbial growth on a solid substrate. However, its limit of detection varies depending on the bacterial species and the growth conditions, typically ranging from > 8 to 100 living cells/mL (Cappuccino and Sherman 2008). Some bacteria may fail to grow on standard media but can persist in microalgal cultures, leading to false confirmation of axenicity. On the other hand, molecular approaches such as 16S rRNA gene sequencing are highly sensitive, with detection limits as low as ~10 cells/mL (Brandt and Albertsen 2018). However, while sequencing can confirm the presence of bacterial DNA, it cannot differentiate between live bacteria and lysed cells. This means that sequencing alone may overestimate contamination levels by detecting residual DNA from non‐viable cells. Microscopy methods, such as epifluorescence or differential interference contrast microscopy, provide direct visualisation of potential contaminants but suffer inherent subjectivity in detecting the type and level of contaminants. The accuracy of this method depends on the experience of the observer and prior knowledge of expected contaminants in a given culture. Moreover, microscopy may fail to distinguish bacterial cells if they are present at very low densities or embedded within algal sheaths. Flow cytometry can be used as an alternative to identify single cells as well as differentiate viability (using the propidium iodide or 7‐aminoactinomycin D). However, the size disparity between microalgae and bacteria makes it challenging to accurately identify ‘singlets’, especially when dealing with closely associated bacteria‐microalgae aggregates. For example, diatoms with tightly bound bacteria within the silica frustule may be detected as a singlet. Additionally, while live/dead staining can be incorporated into flow cytometry protocols to assess bacterial viability, this approach may still fail to detect bacteria in intimate symbiotic relationships with microalgae.
Given these limitations, a combination of complementary testing methods is necessary to deduce axenicity with high confidence. Plate counting provides confirmation of viable bacteria, sequencing ensures highly sensitive detection, and microscopy serves as a cross‐validation tool for detecting bacteria that might be overlooked due to primer biases in PCR or aggregation with microalgae. Flow cytometry could serve as a complementary tool, particularly if live/dead staining is included, but its efficacy depends on the strength of microalgal‐bacterial associations. Based on the method combinations shown in Figure 6, a robust approach could involve using plate counting (or flow cytometry if live/dead staining is implemented) alongside sequencing and microscopy to maximise detection sensitivity while minimising false negatives.
Perspective From Microalgal Culture Collection
4.3
Culture collections are critical towards curating and maintaining microalgal cultures and were identified as a prime source to gain insight from the expertise and knowledge drawn from regular practical work. A query was made to these culture collections to enquire about their practices to achieve axenicity, and the tips and important considerations when maintaining axenicity in working culture. The number of phototrophic eukaryotic microalgae along with the number of axenic species held in four major culture banks is provided in Table S4. Upon enquiry, personnel from CSIRO stated that axenicity was achieved through the use of antibiotics, while NIES exclusively used washing and micro‐picking and did not use antibiotics. The respondent from Bigelow did not mention any specific method but acknowledged that the process can be labour intensive. All the culture collection respondents emphasised the use of good sterile lab techniques to ensure axenicity, with the respondent from Bigelow noting that.‘Maintaining axenic cultures is not difficult if proper working conditions are maintained. We do all work in a flow hood and practice sterile technique’.
The emphasis of the importance of considering the role of commensal/phycospheric microbes was also universally echoed, with the respondent from CCAP stating,‘…however we don't try to make other axenic (unless necessary for a particular project) as many strains don't like to grow well as axenic cultures’.
and from CSIRO,‘We find many do not grow if axenic, possibly due to the need for vitamins etc. produced by bacteria. Streptomycins and G‐penicillin etc can be used—some species will not tolerate them but most will’.
The respondent from NIES stated,‘One of the challenges to maintain axenic strains is that not a few axenic strains grow poorly. This is likely due to the removal of benefit from coexisting bacteria, for example’. [sic]
It is unclear how many of the microalgae in the culture collection are obligate symbionts, and if there has been a systematic attempt to achieve and maintain axenic cultures. The responses from the culture collection institutes suggest that there is currently insufficient interest in the researchers requesting the cultures to generate and maintain axenicity; likely ascribed to a common understanding that beneficial and stable symbiosis between microalgae and phychospheric bacteria is needed to achieve high biomass productivity. The lack of interest can be due to the heavy initial investment and time/labor required for axenisation, lack of knowledge of the media composition to facilitate cultivation of microalgae without the commensal bacteria, the lack of need for high purity cultures for the translational, or the more application‐based research that researchers are generally interested.
Intracellular Symbionts
4.4
Intracellular microbes pose a unique challenge when attempting to generate axenic cultures. Firstly, standard physical methods such as filtration, colony‐picking, etc. remain ineffective as the microbe is often shielded by the microalgal cell wall. For example, Maire et al. (2021) used 3‐D fluorescence in situ hybridisation (FISH) and electron microscopy to demonstrate the presence of phycospheric as well as intracellular bacteria in cultured Symbiodiniaceae cells (endosymbiotic dinoflagellates) making antibiotic treatments less effective. Furthermore, intracellular symbionts can be passed to daughter algal cells during mitosis and therefore persist through cell transfers. For example, N_2_‐fixing cyanobacterial endosymbionts are vertically transmitted in rhopalodiacean diatoms (e.g., Rhopalodia gibba and Epithemia turgida ) during cell division (Prechtl et al. 2004; Nakayama et al. 2014). The cyanobacterial cells have undergone genome reduction and become obligate symbionts making axenisation in such cases impossible. Lastly, many eukaryotic microalgae are cobalamin auxotrophs and associate with bacteria to obtain the necessary nutrients (Croft et al. 2005). Such intricate associations with intracellular microbes make any axenisation treatment, such as the antibiotic‐free workflow being championed in this review, very challenging.
Potential Research Gaps
4.5
Across the collected literature, many points and inconsistencies in the reportage were noted that were of significant importance when trying to study and assess axenisation procedures.
Ambiguous Glossary
4.5.1
Among the collected literature (including reviews), there is significant ambiguity in the use of terms that essentially allude to various aspects of an organism rather than its culture. Terms such as ‘culture’, ‘species’, ‘strain’ and ‘clonal’ have been used interchangeably across the literature although they mean very different things, Belcher and Swale (1988). Even within the context of the collated research, the term species refers to the unique taxonomic identity of the organism being investigated. The term strain refers to a subpopulation of a species that has a property (or set of properties) of interest; for example, the strain IMET1 of Nannochloropsis oceanica is capable of higher lipid production although it is taxonomically identical to other populations of N. oceanica (Ma et al. 2014). Clonal refers to the population that has been derived from a single microalgal cell through asexual reproduction, sharing identical genetic copies and expression. These three terms therefore refer to the investigated organism itself, but not the conditions in which it is grown. The term ‘culture’ refers to the enclosed system in which the organism is grown. Belcher and Swale (1988) explains that,A culture, as applied to algae, refers to the complex of culture medium, the organism(s) growing in the medium, and the vessel enclosing these.
Unfortunately, referral to the culture itself remains inconsistent and synecdochical, with terms such as “unialgal”, “single‐strain”, and “single‐culture” used synonymously to imply axenicity. These terms may refer to the fact that only one algal species is grown in the culture, but does not necessarily imply the absence of non‐algal entities (e.g., bacteria). Thus, inconsistent usage of terms across the discipline contributes to the uncertainty and inaccuracy in the interpretation of the textual reports.
Lack of Attention to the Medium
4.5.2
The growth medium conditions are inherently different from the natural conditions from where the microalgae samples were collected and consequently, the growth dynamics of the microalgae and the contaminating microbes are expected to change. This is particularly true when employing nutrient‐rich heterogenous medium such as Lysogeny Broth which provides a carbon source, and tends to favour bacterial growth over microalgae. Conversely, carbon‐deficient medium such as the f medium relies on the ability of the microalgae to use photosynthesis to assimilate inorganic carbon and achieve dominance over heterotrophic bacteria. The corresponding increase in metabolic stress when using chemical methods such as hypochlorite or antibiotics during axenisation process, however, could significantly affect microalgal cell viability and decrease the chances of successful subculturing. This can be demonstrated using the example of Euglena gracilis , where, in all three reported cases, high antibiotic concentrations were used to achieve axenicity. However, the two studies that successfully established axenic cultures; namely by Gumińska et al. (2018) and Pappas and Hoffman (1952) employed Cramer‐Myers's medium and Hall's medium respectively. Both these media recipes include a carbon source to allow microalgal cells to tap into their mixotrophic pathways. In contrast, Jones et al. (1973), who used artificial seawater, found that the microalgae became unviable before the bacteria were affected. Antibiotics caused chloroplast bleaching in Euglena, thereby impairing their ability to fix carbon. The supplementation of a carbon source, as provided by the media used in Gumińska et al. (2018) and Pappas and Hoffman (1952) may have mitigated the antibiotics' deleterious effects.
Even the manner in which a method is applied in tandem with the medium can affect outcomes. For example, if anoxy is achieved through flushing the medium with sterile CO_2_ (as described by Ma et al. (2017)), it can begin to alter the pH of the culture medium. In such instances, consideration to the buffering capacity, for example through the use of TAP medium has a significantly higher buffering capacity owing to the presence of phosphate buffer salts compared to BBM, and correspondingly, the effectiveness of the applied technique can vary significantly.
Lack of Identification of Co‐Growers
4.5.3
Across the 63 publications, only three supplied sufficient information about the contaminating microbes. This is particularly relevant in cases where chemical methods such as phenol or antibiotics were used to achieve axenicity, as the effectiveness of the applied method is contingent on the relative susceptibility of the contaminating microbe to the treatment in comparison to the microalgae. In other words, if the contaminating/co‐growing microbes are more resilient to the applied treatment in comparison to the microalgae, subsequent efforts to reproduce the outcome may fail even if the described procedure is diligently followed. This is particularly relevant in the case of diatoms, as demonstrated by Grossart (1999) that even within the same culture different micro‐communities of microalgae and bacteria can exist.
Lack of Diversity in Reportage
4.5.4
A wider collection of literature helps ensure a strong representation of techniques, especially across different lab settings. The 81 diatom species noted in Figure 3a–d were obtained from just 11 publications in which 11 methods were described to achieve axenic cultures. Likewise for dinoflagellates, the 36 species were noted in 15 publications and described 13 methods that were used to achieve axenic cultures, while for green algae, 31 species subjected to 19 different methods were described in 32 publications. Finally, in the matter of microalgal divisions represented in the literature, cryptomonads, eugalenoids, red algae, and so on, that were grouped as ‘other eukaryotes’ in Figure 3D, were poorly represented in the collected literature and indicate a potential direction for further research work. A more focused approach towards testing novel methods that is subsequently corroborated by other worker groups/laboratories would significantly increase confidence in the success rates noted for each method. Moreover, since each method (or its combination) has its intrinsic false positive rates, using the same workflow over multiple species, even within the same Division, can affect subsequent efforts to reproduce the results. More work is also required on systematic sampling, identification and purification of microalgae from natural sources. Although axenisation research has been carried out in many countries (Table 1), on‐field sampling was described in only 19 publications (Figure 2). Type‐sampling, archiving and preservation of microalgae from natural water sources has become increasingly relevant as their local populations have become increasingly sparse and the chance of obtaining rarer species has become more precarious owing to the effects of climate change (Gimenez Papiol 2017).
Limitations of This Study
4.6
The work presented here attempted to systematically collect and curate relevant publications, and extract information based upon which cogent axenisation workflows and strategies could be developed. However, there are aspects of this work that require careful interpretation and require further research to furnish missing pieces of data. The limitations of the present study are provided in Table 4.
Limitations of the NCA Approach
4.6.1
Despite the merits of NCA as a powerful abstraction tool for visualising novel ‘pathways’ (axenisation workflows) and being computationally easier to process and study complex processes, it is heavily dependent on the quantity and quality of the data used to construct the networks. Insufficient data or skewed data can generate weak clusters which begin to reflect in the mod value and the edge densities (see the graph outputs of Supporting Information: Code chunks 8, 14 and 20) that can result in weak clusters or malformed cliques. Within the context of this work, a weak edge‐density was interpreted as a lack of data and scope for further work. The networks thus arrived at in Figure 5 serve as a representation of the limited research being invested in only a small group of methods with insufficient cross‐laboratory corroboration.
Further, nuances involved in the implementation of each technique are also lost to the network analysis. For example, the duration of antibiotic incubation, or the optimal flow‐rate used in cell sorting, are all homogenised into a singular representative node, and NCAs are poorly suited to represent specific condition optimisation. Consequently, a chosen workflow may be well suited to eliminate phycospheric bacteria, but may remain ineffective against intracellular bacteria.
Conclusion
5
Promising workflows circumventing the use of antibiotics appear to be filtration ↔ washing ↔ micropicking for diatoms, and micropicking ↔ subculturing ↔ flow cytometry for dinoflagellates. No clear workflow could emerge for green algae although Streak plating ↔ Flowcytometry → Ultrasonication could provisionally be considered. More high quality reports with other methods and microalgal groups are required to estimate success rates and suggest alternate workflow strategies to achieve axenic eukaryotic microalgal cultures. A combination of at least three tests is recommended to verify successful axenisation that could include cell counting (using growth colonies on agar or flow cytometry), sequencing (16S and/or 18S), and microscopy (epifluorescence, light, confocal, etc.)
Author Contributions
A. Iyer: conceptualization, investigation, methodology, software, data curation, formal analysis, supervision, writing – original draft. M. Monissen: methodology, investigation, formal analysis. Q. Teo: formal analysis, methodology, validation, review and editing. O. Modin: funding acquisition, writing – review and editing. R. Halim: writing – review and editing, project administration, resources, supervision, conceptualization, funding acquisition.
Funding
This work was supported by Irish Research Council (GOIPD/2023/1240).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1: Supporting Information
Table S2: Master table with information from 63 publications collected and summarised.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aray‐Andrade, M. M. , M. I. Uyaguari‐Diaz , and J. R. Bermúdez . 2018. “Short‐Term Deleterious Effects of Standard Isolation and Cultivation Methods on New Tropical Freshwater Microalgae Strains.” Peer J 6, no. July: e 5143. 10.7717/peerj.5143.30038855 PMC 6054863 · doi ↗ · pubmed ↗
- 2Belcher, H. , and E. Swale . 1988. Culturing Algae: A Guide for Schools and Colleges. Institute of Terrestrial Ecology.
- 3Brandt, J. , and M. Albertsen . 2018. “Investigation of Detection Limits and the Influence of DNA Extraction and Primer Choice on the Observed Microbial Communities in Drinking Water Samples Using 16S r RNA Gene Amplicon Sequencing.” Frontiers in Microbiology 9, no. September: 2140. 10.3389/fmicb.2018.02140.30245681 PMC 6137089 · doi ↗ · pubmed ↗
- 4Brown, L. M. 1982. “Production of Axenic Cultures of Algae by an Osmotic Method.” Phycologia 21, no. 3: 408–410. 10.2216/i 0031-8884-21-3-408.1. · doi ↗
- 5Cappuccino, J. G. , and N. Sherman . 2008. Microbiology: A Laboratory Manual. 8th ed. Pearson/Benjamin Cummings.
- 6Chamberlain, S. A. , and E. Szöcs . 2013. “Taxize: Taxonomic Search and Retrieval in R.” F 1000 Research 2, no. October: 191. 10.12688/f 1000 research.2-191.v 2.24555091 PMC 3901538 · doi ↗ · pubmed ↗
- 7Clauset, A. , M. E. J. Newman , and C. Moore . 2004. “Finding Community Structure in Very Large Networks.” Physical Review E 70, no. 6: 066111. 10.1103/Phys Rev E.70.066111.15697438 · doi ↗ · pubmed ↗
- 8Croft, M. T. , A. D. Lawrence , E. Raux‐Deery , M. J. Warren , and A. G. Smith . 2005. “Algae Acquire Vitamin B 12 Through a Symbiotic Relationship With Bacteria.” Nature 438: 90–93.16267554 10.1038/nature 04056 · doi ↗ · pubmed ↗