Anopheles mascarensis, a Malaria Vector Endemic to Madagascar and the Comoros Archipelago: A Review
Jessy Goupeyou-Youmsi, Luciano M. Tantely, Tsarasoa M. Andrianinarivomanana, Romain Girod, Catherine Bourgouin

TL;DR
This paper reviews the role of Anopheles mascarensis, a mosquito species in Madagascar and the Comoros, as a malaria vector and explores possible sibling species.
Contribution
The paper updates the geographical distribution of Anopheles mascarensis and suggests it may consist of sibling species based on regional differences.
Findings
Anopheles mascarensis is the main malaria vector in eastern Madagascar but a secondary vector elsewhere.
Populations from the east coast differ from those in the Central Highlands, suggesting sibling species.
Molecular tools could now test the hypothesis of sibling species within An. mascarensis.
Abstract
Anopheles mascarensis (An. mascarensis; De Meillon, 1947) is a mosquito species endemic to Madagascar and the Comoros Archipelago. In the past, it was confused with Anopheles marshalli (Theobald, 1929), a continental African species that does not exist in Madagascar. Malaria transmission is highly heterogeneous in Madagascar. Principal and secondary mosquito vectors, as well as malaria parasite species, may vary from one region to another. Anopheles mascarensis has been identified as the main vector of malaria in the east and southeast of Madagascar, while it plays the role of a secondary vector in other Malagasy regions. Differences in behavior between An. mascarensis populations from the east coast and those from the Central Highlands of Madagascar suggest that An. mascarensis may be composed of sibling species. In the present review, unpublished data on the geographical distribution…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
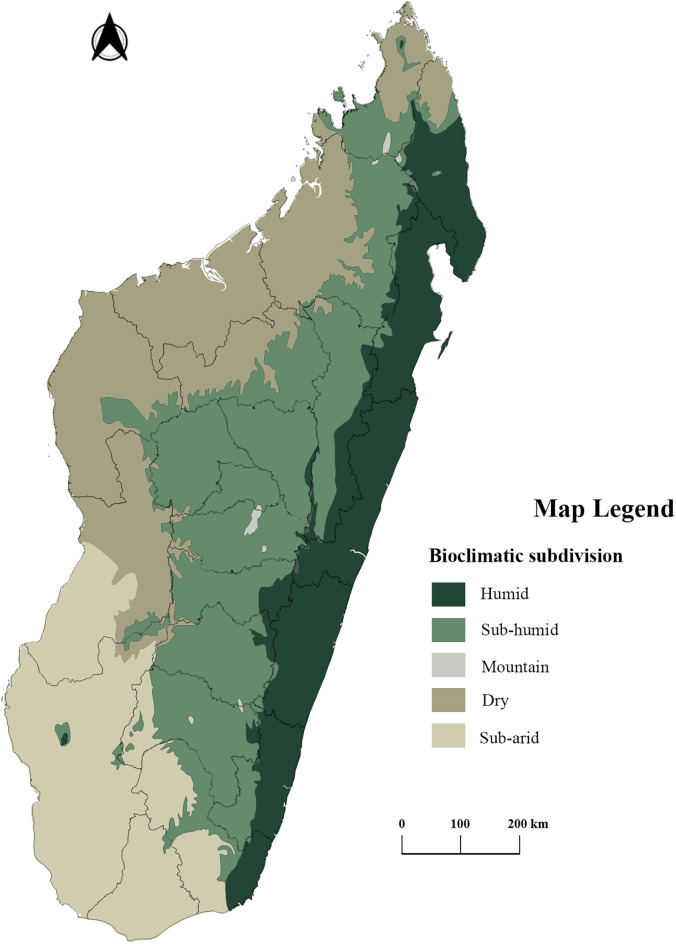 Figure 1
Figure 1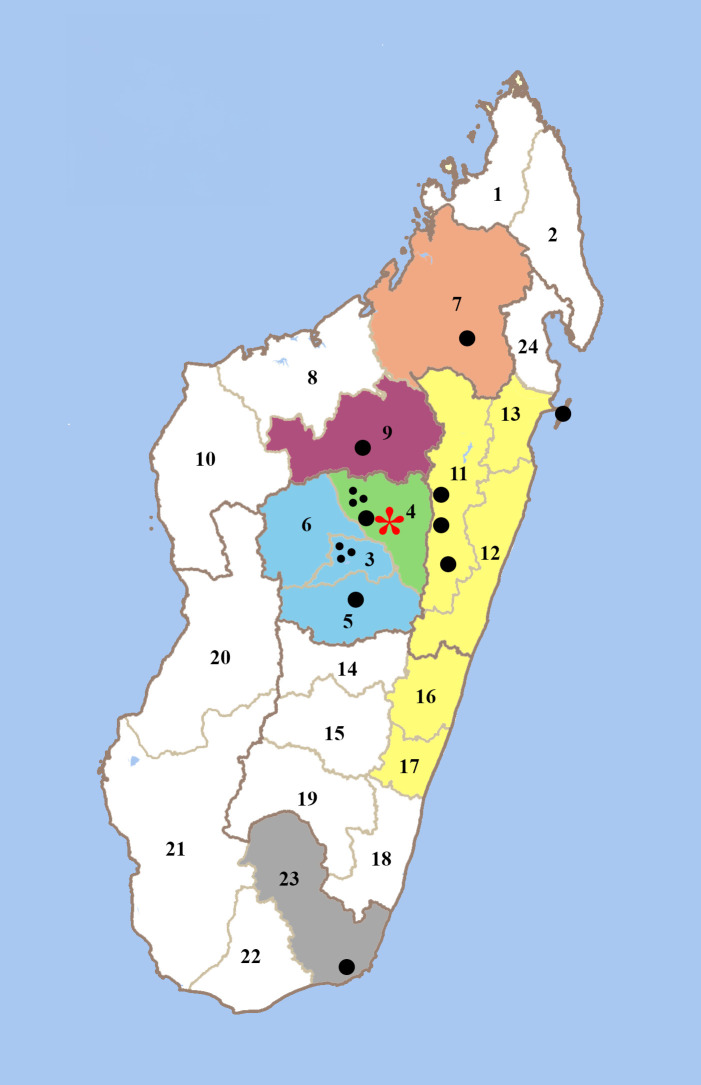 Figure 2
Figure 2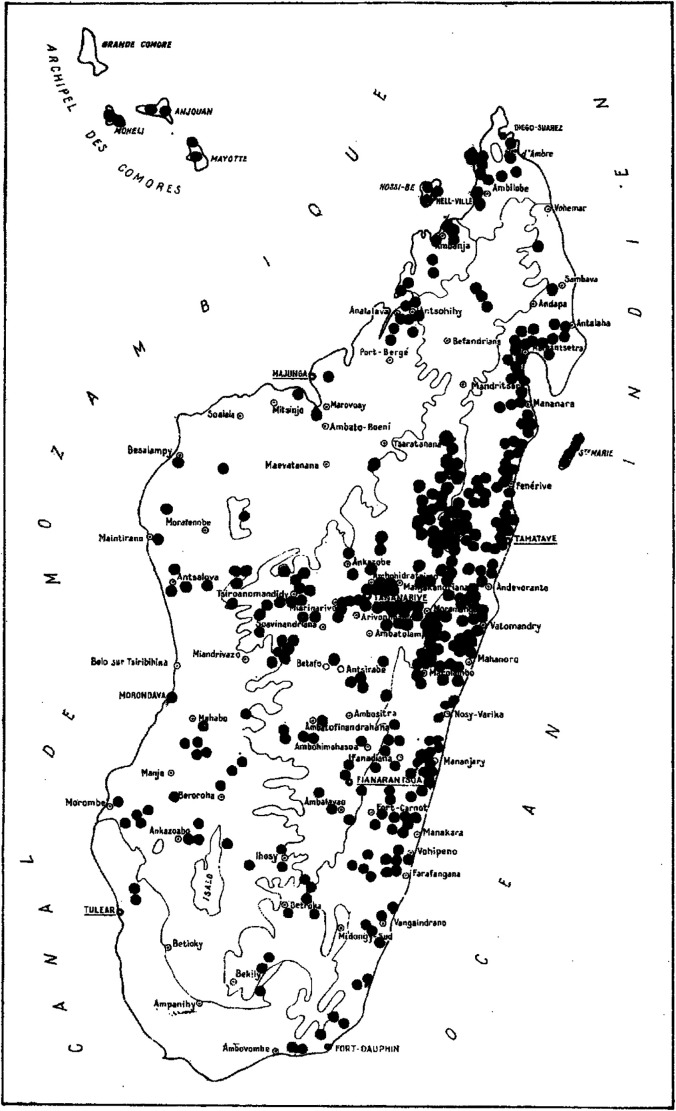 Figure 3
Figure 3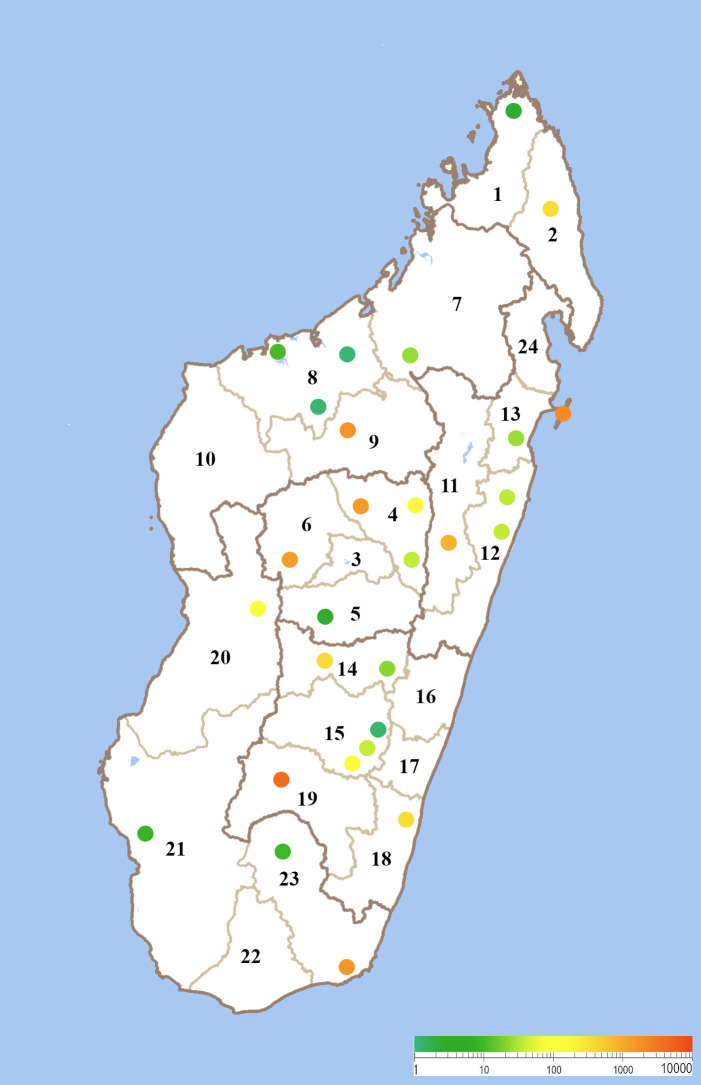 Figure 4
Figure 4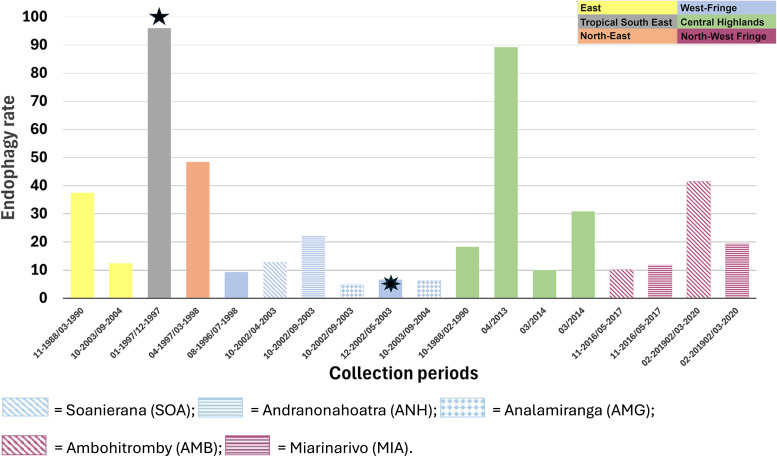 Figure 5
Figure 5| Authors | Date of Publication | Link |
|---|---|---|
| Fontenille, Lepers, et al. | 1992 |
|
| Andrianaivolambo, Domarle, et al. | 2010 |
|
| Tantely, Rakotoniaina, et al. | 2013 |
|
| Marrama, Laventure, et al. | 1999 |
|
| Ravoahangimalala, Rakotoarivony, et al. | 2003 |
|
| Rajaonarivelo, Le Goff, et al. | 2004 |
|
| Le Goff, Randimby, et al. | 2003 |
|
| Laganier, Randimby, et al. | 2003 |
|
| Robert, Le Goff, et al. | 2006 |
|
| Fontenille, Lepers, et al. | 1990 |
|
| Nepomichene, Tata, et al. | 2015 |
|
| Goupeyou-Youmsi, Rakotondranaivo, et al. | 2020 |
|
| Andrianinarivomanana Tsarasoa Malala | 2024 |
|
| Regions | Species/Site/Date | ma | SI% | Annual EIR | Number Tested for Plasmodium via ELISA | Plasmodium Species | References |
|---|---|---|---|---|---|---|---|
| East coast | Sainte Mari | ||||||
|
| 18.08 | 0.75 | 49.48 | 1,864 | Pf |
| |
|
| 16.48 | 1.78 | 107.4 | 9,453 | Pf, Pv, Pm, Po | ||
|
| 0.47 | 0.59 | 1.01 | 338 | Pf | ||
| Saharevo (October 2003/September 2004) | |||||||
|
| 0.74 | 0.75 | 2.03 | 268 | Pf, Pm |
| |
|
| 0.39 | 0.47 | 0.67 | 211 | Pf | ||
|
| 0.65 | 1.58 | 3.75 | 633 | Pf, Pv | ||
| Tropical southeast | Esana (January 1997/December 1997) | ||||||
|
| 7.57 | 0.89 | 24.6 | 677 | Pf |
| |
|
| 2.66 | N/A | 9 | N/A | Pf | ||
|
| 0.66 | N/A | 3 | N/A | Pf | ||
| Northeast | Mandritsara (April 1997/March 1998) | ||||||
|
| 0.83 | NT | NT | NT | – |
| |
|
| 1.34 | 2.13 | 10.41 | 47 | Pf | ||
|
| 4.48 | 1.31 | 21.42 | 842 | Pf | ||
| Western fringe | Ambohimena (August 1996/July 1998) | ||||||
|
| 0.61 | NT | NT | NT | – | ||
|
| 0.35 | 0.00 | 0.00 | 871 | – | ||
|
| 6.73 | 0.2 | 4.91 | 10,753 | Pf | ||
| Soani | |||||||
|
| 1.04 | 0.86 | 3.26 | 116 | Pf |
| |
|
| N/A | N/A | N/A | N/A | – | ||
|
| N/A | N/A | N/A | N/A | – | ||
| Andranonahoatra (October 2002/September 2003) | |||||||
|
| 0.28 | 0.00 | 0.00 | 54 | – |
| |
|
| N/A | N/A | N/A | N/A | – | ||
|
| N/A | N/A | N/A | N/A | – | ||
| Analamiranga (October 2002/September 2003) | |||||||
|
| 3.17 | 0.00 | 0.00 | 722 | – |
| |
|
| 0.72 | 0.00 | 0.00 | 672 | – | ||
|
| 9.15 | 0.17 | 5.68 | 4,056 | Pf, Pv | ||
| Analamiranga (October 2003/September 2004) | |||||||
|
| 1.91 | 0.00 | 0.00 | 590 | – |
| |
|
| 0.59 | 0.00 | 0.00 | 363 | – | ||
|
| 7.55 | 0.07 | 1.93 | 2,774 | Pf | ||
| Central highlands | Manarintsoa (October 1988/February 1990) | ||||||
|
| 1.14 | 0.00 | 0.00 | 178 | – | ||
|
| 1.67 | 0.11 | 0.67 | 2,759 | Pf, Pv, Pm | ||
|
| 0.09 | 0.47 | 0.15 | 214 | Pf | ||
| Ankazobé (December 1989/April 1990) | |||||||
|
| N/A | 0.00 | 0.00 | 63 | – | ||
|
| 0.96 | 0.87 | 3.05 | 115 | Pf, Pv, Pm | ||
|
| 20.82 | 0.92 | 70.08 | 2,498 | Pf, Pv, Pm | ||
| Northwestern fringe | Andriba (December 1989/April 1990) | ||||||
|
| N/A | 0.00 | 0.00 | 180 | – | ||
|
| 8.47 | 0.16 | 0.13 | 611 | Pf, Pv | ||
|
| 8.83 | 1.8 | 58.01 | 276 | Pf, Pv | ||
| Andriba (November 2016/April 2017) | |||||||
|
| 0.92 | 0.00 | 0.00 | 59 | – |
| |
|
| 5.81 | 2.41 | 51.11 | 374 | Pf, Pv | ||
|
| 2.53 | 1.42 | 13.11 | 212 | Pf, Pv | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Parasitic Diseases Research and Treatment
INTRODUCTION
Anopheles mascarensis (An. mascarensis) is a mosquito species endemic to Madagascar and the Comoros Archipelago. It was described by De Meillon in 19471 on the basis of unique adult samples collected in Diego Suarez (which is actually Antsiranana), at the northern tip of Madagascar. It was named after the Mascarene Islands, which include Mauritius, La Réunion, and Rodrigues, located on the eastern side of Madagascar.2 It has long been confused with Anopheles marshalli (An. marshalli) Theobald, a continental African species that does not exist in Madagascar.3^,^4 Anopheles mascarensis belongs to the Cellia subgenus of the Anopheles genus, which includes many important malaria vectors in Africa and Asia.5^,^6 Several entomologists have reported subtle but significant morphological variations suggesting the existence of geographical variants of An. mascarensis.3 During numerous malaria surveillance campaigns over the last century, the presence of An. mascarensis was reported in several places in Madagascar, but it was never implicated as a potential malaria vector until 1992.3^,^7^,^8 In 1992, Fontenille and colleagues9^,^10 found that An. mascarensis was an important secondary vector of human malaria on Sainte Marie Island, located off the east coast of Madagascar. In 1999, Marrama et al. reported that An. mascarensis acted as the primary malaria vector in a locality near Fort Dauphin (which is actually Tolagnaro) in the tropical southeastern region of Madagascar.11
The epidemiology of malaria transmission in Madagascar is complex because of the geography of the country and its climatic stratification.12??^–^15 A topographical map of Madagascar can be found at https://commons.wikimedia.org/wiki/File:Madagascar_Topography.png. Five bioclimatic domains have been proposed by Cornet16 (Figure 1). A brief description of these bioclimatic domains can be found in Pock Tsy et al.17 Anopheles funestus (An. funestus), Anopheles gambiae s.s. (An. gambiae), and Anopheles arabiensis are the main malaria vectors in most regions of Madagascar. The tremendous malaria vector control efforts put in place after the devastating malaria epidemics that occurred between 1986 and 1988 revealed changes in malaria vector distribution, notably the reemergence of An. funestus as a major malaria vector in the Central Highlands.18??^–^21 Despite continuous malaria control efforts, malaria remains a major health problem in Madagascar, which is currently facing both climatic and societal changes that have increased poverty in areas where malaria has reestablished itself as a major threat.22^,^23 The situation has worsened after the coronavirus disease 2019 pandemic.24 In this context, whether An. mascarensis acts as a locally secondary or major vector remains an open question. Furthermore, the descriptions of geographical, morphological, and behavioral variants over the last 50 years suggest that An. mascarensis may comprise a complex of sibling species with possibly different vectorial capacities.3^,^9
{kind=link}
Madagascar’s bioclimatic domains. Adapted from Cornet 1973 (https://datasuds-geo.ird.fr/geonetwork/srv/resources/records/0839b2e5-596a-48e5-8678-1af779dffa72), with modifications performed by the Geomatic Group of the Epidemiology Unit at the Institut Pasteur de Madagascar.
In the present review, published data on the geographical distribution of An. mascarensis, its biology, and its contribution to malaria transmission were analyzed. The list of publications used is presented in Table 1, and the localization of the investigated sites is shown in Figure 2. Furthermore, information was extracted from the database of the Medical Entomology Unit at the Institut Pasteur de Madagascar, spanning the period from 1989 to 2015. The current work provides a foundation for further investigation into whether An. mascarensis is indeed a complex of species with distinct geographical distributions, behaviors, and malaria vectorial capacities.
Localization of the field sites where the presence of Anopheles mascarensis was reported (1990–2024). The limits of the 24 different administrative regions are indicated, as well as their official numbered code according to https://fr.wikipedia.org/wiki/R%C3%A9gions_de_Madagascar#/. Black dots = sampled sites; blue = western fringe; gray = tropical southeast; green = Central Highlands; orange = northeast; purple = northwestern fringe; red star = Antananarivo; yellow = east coast.
RESULTS
Systematic position.
According to Alexis Grjebine, An. mascarensis was once possibly named Anopheles funesta var neireti by Blanchard in 1906 and later An. funestus var imerensis by Monier and Treillard in 1935, on the basis of abundant samples collected around Antananarivo.8^,^25 Unfortunately, none of these mosquitoes were preserved. Until the holotype description by De Meillon in 1947, An. mascarensis was long described as An. marshalli.1 Even later, it was still often confused with this species, despite the typical morphological characteristics of the cibarial armature of the adults described by De Meillon.3^,^4 The larval and pupal stages were only described in 1961.26 In this latter publication, the authors discussed the presence or absence of some characteristics of An. mascarensis adults that could call into question the classification of this species within the Neomyzomyia series, as proposed by De Meillon. The most recent classification has placed An. mascarensis within the subgenous Cellia Theobald and the Neomyzomyia series.27 Within this series, a Mascarensis group that contains the sole An. mascarensis species was proposed.28 In contrast, An. marshalli Theobald belongs to the Myzomyia series of the Cellia subgenus. It was later classified in the Marshalli complex29 within the Marshalli group.30 This latter information was assembled by Harbach.31^,^32 Additional information can be found in Irish et al.33
Geographical distribution.
In his comprehensive review on Malagasy anophelines published in 1966, Grjebine assembled a wealth of information on both the chorology and biology of An. mascarensis, and provided the first distribution map of An. mascarensis in Madagascar and the Comoros Archipelago.8 Most records were established over a 10-year period from 1952 to 1962, revealing the presence of An. mascarensis in ∼400 localities across Madagascar (Figure 3). With the exception of the most southern arid part of Madagascar, An. mascarensis was widespread in every region and at all elevations, from 1,200 m to the coastline. It was also found in Anjouan, Moheli, and Mayotte in the Comoros Archipelago. Its abundance varied from one locality to another, with a trend toward higher abundance on the east coast of Madagascar (Ivoloina, Toamasina, Sainte-Marie Island, and Mananara).3^,^7^,^8^,^10^,^26 On the west coast, it was scarce in some localities.7^,^8^,^34 In the Central Highlands, it was found in many places from north to south and from the eastern to western fringes, as well as in the vicinity of Antananarivo, the capital city of Madagascar.
Geographical distribution of Anopheles mascarensis (Grjebine, 1966). Reproduction authorized by IRD Editions.
Published records on the occurrence of An. mascarensis are almost nonexistent from 1966 to 1988. However, during arbovirus prevalence surveys in the Tsiroanomandidy Region (the western fringe of the Central Highlands) between 1984 and 1986, An. mascarensis was reported as the second most prevalent Anopheles mosquito after Anopheles maculipalpis.35 Notably, a similar survey covering the entirety of Malagasy Island from January 1982 to May 1988 revealed the presence of An. mascarensis in multiple sites across the country, confirming a higher prevalence of this mosquito species in the eastern region of Madagascar, as well as its absence or scarce occurrence in the north and south.36 Finally, after the 1980s malaria epidemics in the Central Highlands, a specific survey on malaria vectors was conducted in 1987 in three villages located near Antananarivo.18 The abundance of An. mascarensis was similar to that of the suspected malaria vectors An. gambiae and An. funestus, but far lower than that of Anopheles coustani (An. coustani) and Anopheles squamosus (An. squamosus), both of which have been recognized as potential secondary malaria vectors in Madagascar.37 For the Comoros, An. mascarensis larvae were found in Anjouan in 1974,38 and adults were found in Mayotte in 1972.39 A more recent survey (2008–2012) revealed the continued presence of An. mascarensis in Mayotte.40
To complement this information, records of An. mascarensis collated in the mosquito database of the Medical Entomology Unit at the Institut Pasteur de Madagascar were extracted. The samples correspond to entomological surveys performed from 1989 to 2015 using diverse methods, including odor-baited traps (human and animal), CDC light-traps, Muirhead–Thomson pit traps, catches in stable and outdoor animal shelters, human landing catches (HLCs), and indoor pyrethroid spraying. Only a subset of this database has been published.41??^–^44 The entomological surveys were performed in 126 localities (or villages). Anopheles mascarensis was found in 93 localities distributed across 28 districts, covering 18 of the 24 administrative regions of Madagascar (Supplemental Table 1). According to these records, 13,026 An. mascarensis were caught during these 26 years of entomological surveys (Supplemental Table 2). With the exception of the years 1989, 1993, 1994, and 2005, An. mascarensis was caught every year. A plot of the abundance of An. mascarensis by district is presented in Figure 4. However, it is important to note that some places were surveyed only once, whereas others benefited from longitudinal surveys. This map confirms the presence of An. mascarensis in areas where it was already present in the 1960s. The absence of records in some areas, when compared with the 1960s map, does not necessarily indicate a decline in its distribution over time. Indeed, most surveyed localities were included in malaria surveillance activities over the period spanning from 1989 to 2015.
Distribution map of Anopheles mascarensis collected from 1989 to 2015. Data were extracted from the mosquito database of the Medical Entomology Unit at the Institut Pasteur de Madagascar. The limits of the 24 different administrative regions are indicated. Colored dots correspond to the 28 sampled districts (Supplemental Table 2), and abundance is indicated according to the colored scale.
Biology.
Breeding sites.
Anopheles mascarensis usually breeds in renewable watercourses, particularly in transplanted or uncropped rice fields.8^,^45 The larvae dwell in both forest streams and water bodies in deforested areas. They also breed in irrigation canals and streams.8^,^26^,^46 In Anjouan (Comoros), they were found in watercourses38 and in marshy areas, such as in Mayotte.40 Anopheles mascarensis larvae can tolerate streams in which the current reaches 40 cm per second; however, they are primarily found in still water ponds.8 In sunny rice fields, they look for shaded areas provided by shrubs, trees, or any upright vegetation. Regarding association with other malaria vectors, An. mascarensis larvae are commonly found alongside An. gambiae s.l. and An. coustani larvae, but rarely with An. funestus larvae.8
Trophic and resting behavior.
In his seminal work, which was conducted in several regions of Madagascar, Grjebine concluded that An. mascarensis is a polyphagous and predominantly zoophagic species that feeds on humans when cattle are rare.8 He reported that anthropophagy can vary from 0% to 85%, with geographic variations possibly linked to agricultural and cultural practices. The published reports covering the period from 1988 to 2024 (Table 1) provide limited information on the trophic behavior of An. mascarensis, based on blood meal analysis. Only three publications revealed data indicating an anthropophagic rate that varies from 0% to 7.9%.41^,^42^,^47 Most mosquitoes feed on bovines. The HLC information from the same set of publications and from the Institut Pasteur de Madagascar database covering the years from 1989 to 2015 confirms Grjebine’s report,8 which reveals that An. mascarensis is exophagic, preferentially feeding outside (Supplemental Tables 2 and 3, Figure 5), with two exceptions. First, the populations collected in Fort Dauphin were highly endophagic (rates of 87.87–96.04%). As hypothesized by Marrama et al.,11 the endophagic behavior of An. mascarensis in Fort Dauphin was likely due to constant high winds blowing in that area. The other population with a high endophagic rate (89.29%) was collected in the Central Highlands (Figure 5, Supplemental Table 3). However, the sample size was limited (n = 28), and the endophagic rate of mosquitoes from neighboring villages was less than 31% (Figure 5, Supplemental Table 3). Regarding the resting behavior of An. mascarensis, both unpublished (Supplemental Table 2) and published data (Supplemental Table 4) confirm that the surveyed populations are preferentially exophilic, in agreement with Grjebine’s report.8 The sample size of the single record revealing a high endophilic rate (83.33%) was quite low (n = 6), with no additional information on the climatic conditions at the time of the survey (i.e., high wind).48 Globally, An. mascarensis has a tendency for exophagy (feeding outside) and exophily (resting outside).
Endophagic rate of Anopheles mascarensis in different regions of Madagascar. Within the western and northwestern fringe regions, villages that were sampled during different periods are labeled. At the site with the eight-pointed star, human landing catches were performed, along with captures using Mbita traps, corresponding to the pooled data from three villages (SOA, ANH, and AMG; Supplemental Table 3). Five-pointed star = the weather was highly windy, possibly affecting the outdoor collection; eight-pointed star = the lowest value reported was less than 6%.
Insecticide resistance.
Possibly because of its exophilic and exophagic behavior, as well as its known zoophily, An. mascarensis was not included in systematic insecticide resistance assays until recently. Past records have revealed the high frequency of nulliparous females after insecticide treatments, suggesting that in the 1960s, this mosquito species was fully susceptible to insecticides.8 To the best of the authors’ knowledge, a single recent report addressed An. mascarensis’ susceptibility status to insecticides.49 From indoor-collected adults at four sites in the east and southeast of Madagascar, it can be concluded that in those areas, An. mascarensis is fully susceptible to permethrin, deltamethrin, and pirimiphos-methyl, despite small sample sizes (18 < n <65). Similar results were obtained in the president malaria initative Africa indoor residual spraying project.50
Vector role of Anopheles mascarensis.
Malaria.
In Madagascar, it has long been believed that malaria transmission is only due to An. funestus and An. gambiae sensu lato. Intensive entomological surveys conducted until the 1960s did not reveal any An. mascarensis mosquitoes bearing sporozoites in dissected salivary glands, either on the east coast (4,350 mosquitoes) or the west coast (959 mosquitoes).7 In contrast, because of their abundance, An. coustani and An. squamosus were suspected to act as local malaria vectors in areas with residual malaria, where An. gambiae and An. funestus were barely detected.37 On the basis of the abundance criterion, Fontenille et al.10 suspected that An. mascarensis could be an important malaria vector in the Sainte Marie island, where malaria prevalence reached 85% in children aged 10–15 years old. Thanks to the development of the ELISA technique for detecting Plasmodium circum-sporozoite protein in preserved mosquito heads and thoraces,51 they showed that An. mascarensis was indeed a vector of human Plasmodium with the detection of Plasmodium falciparum (P. falciparum) in 14 mosquitoes among 1,864 tested, corresponding to a mean sporozoitic index (SI%) of 0.75. During that survey, infection of An. gambiae s.l. by any of the four human Plasmodium species present in Madagascar was high, with 165 infected mosquitoes among 9,453 tested (mean SI% value of 1.78).10 In the following years, the systematic detection of Plasmodium sporozoites in An. mascarensis, as determined via ELISA or polymerase chain reaction (PCR), was performed during entomological surveillance campaigns and malaria epidemiology studies. This led to the identification of evidence suggesting that in the tropical southeast of Madagascar, An. mascarensis was the main malaria vector, even if An. gambiae s.l. and An. funestus were present.11 Indeed, An. mascarensis was responsible for two-thirds of the infective bites.
A comprehensive summary of the published data from Table 1 and Figure 2 is presented in Table 2, which includes any instances in which the three species, An. mascarensis, An. gambiae s.l., and An. funestus, were collected at the same site and period via HLC and subsequently tested for Plasmodium sporozoites. From these published data and for ease of comparison, an annual entomologic inoculation rate (EIR) was calculated, although transmission does not occur year-round at every site and data were not collected over the same time period. These collated data confirm the important contribution of An. mascarensis to malaria transmission in the eastern part of Madagascar, likely due to the abundance of this species. Notably, Le Goff et al.52 reported the detection of 11 P. falciparum-positive An. mascarensis mosquitoes out of a total of 2,463 specimens (SI%: 0.45) in the east and tropical south, according to surveys performed between 1997 and 2003. In contrast, they reported only one P. falciparum-positive An. mascarensis mosquito out of a total of 2,214 specimens (SI%: 0.045) in the western fringe of Madagascar, over the same time frame. In the Central Highlands and northwestern fringe, no Plasmodium sporozoites were detected in An. mascarensis; however, both An. gambiae s.l. and An. funestus were found to be infected with Plasmodium (Table 2).9^,^53
Whether abundance, vectorial competence, or both contribute to the role of An. mascarensis as a malaria vector is an unanswered question. To address the contribution of abundance, the proportion of An. mascarensis was compared among the three major Malagasy vectors, An. mascarensis, An. gambiae s.l., and An. funestus, across different investigated regions in Madagascar. To accomplish this aim, mosquito numbers captured by HLC were only extracted from published records. The relative abundance of An. mascarensis varies greatly geographically (Supplemental Figure 1, Supplemental Table 5). It is predominant in the tropical southeast, representing roughly 50% of malaria vectors in the east and Central Highlands. Therefore, in those regions, its high abundance is likely responsible for its role as a malaria vector. In all other regions where its relative abundance is low, An. mascarensis was not found to bear Plasmodium sporozoites, except in Soanierana. In this locality, despite its low abundance, An. mascarensis co-occurring with An. gambiae s.l. can maintain a weak malaria transmission in the western fringe region in the absence of the main vector, An. funestus.52 In conclusion, the currently available dataset suggests that abundance is the main driver of malaria transmission by An. mascarensis.
Filariasis.
Although Grjebine mentioned that An. mascarensis was found with metacyclic microfilariae on the east coast,8 likely caused by Wuchereria bancrofti (W. bancrofti), this information could not be traced to an accessible publication. However, Brunhes and collaborators54^,^55 were able to experimentally infect two female An. mascarensis mosquitoes fed on the blood of a patient infected with W. bancrofti.
Arboviruses.
In a systematic search and inventory for arboviruses circulating in Madagascar, An. mascarensis was found to be naturally infected by the Ngari virus in the Anjiro Region, which is located 72 km east of Antananarivo, on two occasions.36 Originally isolated from Senegalese Aedes simpsoni males in 1979, the Ngari virus (Bunyaviridae family, Orthobunyavirus genus) was responsible for hemorrhagic fever outbreaks in humans, from whom it was first isolated in 1993.56 Until recently, An. mascarensis was not implicated as a putative vector for other arboviruses. However, in 2021, after an outbreak of Rift Valley fever virus (RVFV) on the east coast of Madagascar, high numbers of captured An. mascarensis were found to be harboring RVFV RNA via PCR testing in 9 of 121 pools of roughly 10 females each.57 Determining the actual role of An. mascarensis in RVFV transmission requires additional investigation through virus isolation from wild mosquitoes and experimental infections in the laboratory.
Anopheles mascarensis: a potential complex of sibling species.
In his careful work, in which he aimed to demonstrate that An. mascarensis is sometimes confused with An. marshalli, Chauvet established biometric diagnosis parameters.3 These biometric characteristics led him to demonstrate that populations of An. mascarensis from the east coast (Ivoloina) differ from those collected near Antananarivo in the Central Highlands. These characteristics included the color of the palps (light versus dark), wing length, and banding pattern. Already at that time (1962), indicating these observed geographical variations, Chauvet wrote that An. mascarensis was possibly a species in full evolution. Based on the seminal work of M. Coluzzi on the An. gambiae s.l. mosquito, Chauvet’s observations were indeed suggestive that An. mascarensis could be a complex of species. Later on, Fontenille and Campbell9 provided additional behavioral evidence that An. mascarensis populations from the east coast differed from those in the Central Highlands: one population is anthropophilic, whereas the other is zoophilic. They suggested that An. mascarensis is a complex of sibling species with different vectorial capacities. Furthermore, Le Goff et al.52 reported considerable differences between the vector role and biology of An. mascarensis populations from the west and those from the east coast. However, a study on the genetic polymorphism of An. mascarensis from Madagascar, conducted using random amplified polymorphic DNA PCR testing, revealed that there was no genetic differentiation between the populations from the coast and those from the Central Highlands.58 With access to novel technologies such as next generation sequencing, it would be wise to reassess the genetic makeup of different populations of An. mascarensis to more precisely document the possible existence of sibling species within the An. mascarensis species.
Miscellaneous information.
In this section, three sets of published data not discussed in the previous sections are reported. First, an odor-baited entry trap investigation conducted in three villages located in the western, eastern, and southern fringes of Madagascar revealed that An. mascarensis is preferentially attracted to calves.59 Secondly, Zohdy and collaborators conducted a study in six villages located near the Ranomafana National Park in the southeastern humid tropical forest, with the aim of testing the efficacy of a human microbiota-derived attractant in CDC light traps to capture Anopheles mosquitoes. The comparative assay (with or without the attractant) revealed that the attractant has no effect on Anopheles trapping, including An. mascarensis. Their study indicates that more An. mascarensis were trapped in agricultural sites rather than in villages or forest sites.60^,^61 Surprisingly, in another study performed in the same region 3 years later using CDC light traps, no An. mascarensis were captured.62 Thirdly, an innovative strategy was developed by Tedrow et al.,63 who combined a modified barrier screen for outdoor Anopheles capture and a multiplex molecular assay capable of identifying mosquito species, blood meal origin, and Plasmodium sporozoites. In their study conducted in the Tsiroanomandidy District in December 2017 and April 2018, an equal proportion of An. mascarensis had fed on humans in December and in April (35%), whereas the proportion that had fed on cows shifted from 35% in December to 50% in April. Two individual An. mascarensis mosquitoes were found to be infected with P. falciparum and both P. falciparum and Plasmodium vivax, respectively.
CONCLUSION
The data analyzed in this review support the theory that An. mascarensis is a complex of sibling species, given its abundance relative to An. gambiae s.l. and An. funestus, which share similar biotopes, endophagic behavior, and vectorial capacity (estimated as annual EIR). Supporting the observations and data reported by both Chauvet and Fontenille and Campbell, the collated data highlight a notable difference between populations from the east coast and those from the Central Highlands of Madagascar. Testing the vectorial competence of each population against a reference strain of P. falciparum could provide an additional phenotypic tool. With advances in molecular tools, testing the hypothesis that An. mascarensis is a complex of sibling species is clearly within reach. For instance, using a set of nuclear and mitochondrial markers, such as 18S-internal transcribed spacer and cytochrome oxidase subunit I, could aid in further classification. The recent publication of the first genome of An. mascarensis will undoubtedly provide additional information and tools (April 2025; https://www.ncbi.nlm.nih.gov/bioproject/1197942). Furthermore, it would also be interesting to test this hypothesis with An. mascarensis populations from the Comoros Archipelago.
Supplemental Materials
10.4269/ajtmh.24-0697Supplemental Materials
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1De Meillon B, 1947. New records and species of biting insects from the Ethiopian region. J Entomol Soc South Afr 10: 110–124.20266389 · pubmed ↗
- 2Kitzmiller JB, 1982. Anopheline Names: Their Derivations and Histories. Annapolis, MD: Entomological Society of America.
- 3Chauvet G, 1962. Variabilite geographique chez les femelles d’Anopheles mascarensis de Meillon, 1947. Absence d’A. marshalli Theobald, 1929 a Madagascar. Bulletin de La Société de Pathologie Exotique 55: 1145–1156.
- 4De Meillon B, 1947. The Anophelini of the Ethiopian Geographical Region. Johannesburg, South Africa: South African Institute of Medical Research, 272.
- 5Sinka M, , 2012. A global map of dominant malaria vectors. Parasit Vectors 5: 69.22475528 10.1186/1756-3305-5-69PMC 3349467 · doi ↗ · pubmed ↗
- 6Pondeville E Puchot N Lang M Cherrier F Schaffner F Dauphin-Villemant C Bischoff E Bourgouin C, 2019. Evolution of sexually-transferred steroids and mating-induced phenotypes in Anopheles mosquitoes. Sci Rep 9: 4669.30874601 10.1038/s 41598-019-41094-4PMC 6420574 · doi ↗ · pubmed ↗
- 7Chauvet G Coz J Gruchet H Grjebine A Lumaret R, 1964. [Contribution to the biological study of malaria vectors in Madagascar. Results of 5 years’ study (1958–1962)]. Med Trop (Mars) 24: 29–44.14142039 · pubmed ↗
- 8Grjebine A, 1966. Insectes Diptères Culicidae Anophelinae. Faune de Madagascar. Paris, France: ORSTOM CNRS, 489.