Submicroscopic Burden of Zoonotic Plasmodium knowlesi Malaria on Mursala Island and Plasmodium falciparum and Plasmodium vivax Transmission in Mainland North Sumatra, Indonesia
Inke Nadia Diniyanti Lubis, Ranti Permatasari, Lambok Siahaan, R. Andika Dwi Cahyadi, Irbah Rhea Alvieda Nainggolan, Rycha Dwi Syafutri, Monica Nadya Sinambela, Silvia Jauharah, Agatha Lestari, Minerva Theodora, Hellen Prameswari, Kim A. Piera, Bridget E. Barber

TL;DR
This study finds that zoonotic malaria caused by Plasmodium knowlesi is rare in North Sumatra but can lead to severe cases, highlighting the need for molecular testing.
Contribution
The study uses ultrasensitive PCR to detect submicroscopic zoonotic malaria in Indonesia, revealing low transmission of Plasmodium knowlesi.
Findings
7 out of 64 participants on Mursala Island had Plasmodium knowlesi infections, undetected by microscopy.
No Plasmodium knowlesi infections were found among 947 mainland participants.
Molecular diagnostics are essential for identifying submicroscopic zoonotic malaria cases.
Abstract
Accurate molecular tools are essential for estimating zoonotic malaria transmission in Southeast Asia. This study applied ultrasensitive reverse-transcriptase real-time polymerase chain reaction (PCR) to detect zoonotic malaria in febrile patients from health facilities across three mainland districts (Batubara, Tanjung Balai, and Central Tapanuli) and separately on Mursala Island, North Sumatra, Indonesia. Among 64 participants on Mursala, 7 (10.9%) adults had Plasmodium knowlesi infections (including 5 agricultural workers and 2 adults with severe WHO anemia), and 2 (3.1%) adults had Plasmodium vivax infections. All were negative by microscopy and panparasite lactate dehydrogenase rapid diagnostic tests. No P. knowlesi infections were identified among 947 participants from mainland sites; PCR detected confirmed Plasmodium species in 26%, including P. vivax (17.5%) and Plasmodium…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1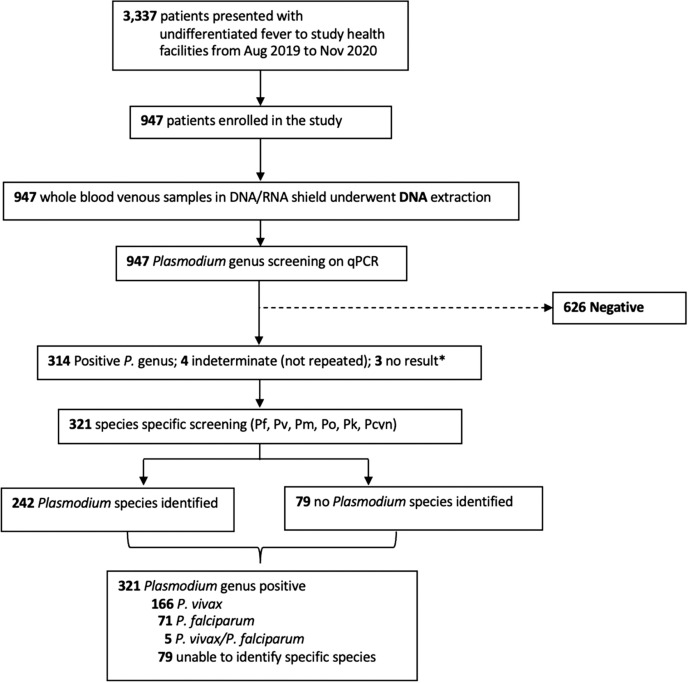 Figure 2
Figure 2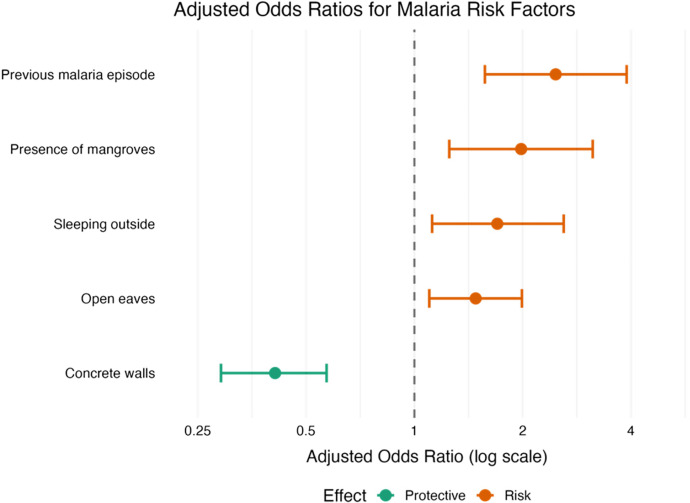 Figure 3
Figure 3| Patient Characteristics | North Sumatra—Mainland Sites | |||||
|---|---|---|---|---|---|---|
|
|
|
| Control (negative) | |||
| No. enrolled | 71 | 166 | 5 | 79 | 626 | |
| Age, median years | 16 | 16 | 18 | 15 | 18 | 0.063 |
| IQR | 12 | 13 | 14 | 17 | 26 | |
| Range | 2–55 | 4–65 | 4–30 | 2–54 | 1–80 | |
| Children (age younger than 15 years old), | 25 (35.2) | 58 (34.9) | 2 (40) | 37 (46.8) | 229 (36.6) | 0.46 |
| Male gender, | 38 (53.5) | 100 (60.2) | 4 (80) | 33 (41.8) | 321 (51.3) |
|
| Previous malaria (self-reported), | 15 (21.1) | 31 (18.7) | 2 (40) | 5 (6.3) | 40 (6.4) |
|
| Symptoms on enrolment, | ||||||
| Headache | 70 (98.6) | 147 (88.6) | 4 (80) | 62 (78.5) | 503 (80.4) |
|
| Nausea | 54 (76.1) | 127 (76.5) | 4 (80) | 51 (64.6) | 422 (67.4) | 0.069 |
| Cough | 41 (57.7) | 75 (45.2) | 2 (40) | 40 (50.6) | 324 (51.8) | 0.45 |
| Shortness of breath | 11 (15.5) | 17 (10.2) | 0 (0) | 15 (18.9) | 140 (22.4) |
|
| Vomiting | 37 (52.1) | 75 (45.2) | 2 (40) | 29 (36.7) | 276 (44.1) | 0.432 |
| Abdominal pain | 36 (50.7) | 60 (36.1) | 4 (80) | 39 (49.4) | 313 (50) |
|
| Diarrhea | 8 (11.3) | 17 (10.2) | 0 (0) | 6 (7.6) | 68 (10.9) | 0.921 |
| Bleeding | 0 (0) | 5 (3.0) | 0 (0) | 1 (1.3) | 4 (0.6) | 0.106 |
| Convulsions | 10 (14.1) | 13 (7.8) | 0 (0) | 4 (5.1) | 118 (18.8) |
|
| Fever (temperature >37.5°C), | 35 (49.3) | 101 (60.8) | 2 (40) | 51 (64.6) | 293 (46.8) |
|
| Febrile days (before presentation), median (IQR, range) | 6 (4, 1–40) | 6 (4, 1–30) | 7 (7, 5–15) | 5 (3, 2–25) | 5 (4, 0–40) | |
| Microscopy result | ||||||
|
| – | 99 | 2 | 2 | 11 | – |
|
| 42 | 2 | 1 | 2 | 10 | – |
|
| 20 | 44 | 2 | 3 | 7 | – |
|
| 1 | – | – | – | – | – |
|
| – | 1 | – | – | – | – |
| Negative | 8 | 20 | – | 72 | 598 | – |
| Sensitivity, % (95% CI) | 59.2 (46.8–70.7) | 59.6 (51.8–67.2) | 60 (14.7–94.7) | – | 95.5 (93.6–97) | |
| Specificity, % (95% CI) | 98.4 (97.3–99.1) | 98.1 (96.9–98.9) | 82.2 (79.6–84.6) | 68.8 (63.5–73.9) | ||
| Parasite count | ||||||
| Positive result, | 62 (87) | 145 (87) | 5 (100) | 7 (8.9) | – | |
| Parasites/ | 3,309 (2,060–5,315) | 4,199 (3,510–5,023) | 8,010 (4,139–15,502) | 7,026 (3,330–14,822) | – | |
| Parasites/ | 4,870 | 5,009 | 9,638 | 4,525 | – | |
| IQR | 1,330–11,673 | 2,019–9,017 | 7,068–13,007 | 3,668–14,136 | – | |
| Range | 8–53,850 | 200–54,800 | 2,340–15,904 | 2,305–36,280 | – | |
| RDT result | ||||||
| No. conducted, | 51 (72) | 123 (74) | 3 (60) | 72 (91) | 441 (71) | |
| Sensitivity, % (95% CI) | 82.4 (69.1–91.6) | 81.3 (73.3–87.8) | 100 (29.2–100) | 11.1 (4.9–20.7) | – | |
| Specificity, % (95% CI) | 78.5 (75.1–81.6) | 86.0 (82.9–88.8) | 73.9 (70.5–77.2) | 71.8 (68.1–75.4) | 93.4 (90.7–95.6) | |
| Anemia | 20/43 (47) | 41/92 (45) | 3/4 (75) | 15/37 (41) | 120/398 (30) |
|
| Creatinine | 106 (92–124) | 115 (97–133) | 115 (102–155) | 97 (94–115) | 97 (87–115) | 0.003 |
| Platelet count, ×103/ | 102 (55–137)[12–425] | 98 (68–146)[11–565] | 75 (58–133)[50–266] | 227 (148–309)[11–520] | 237 (162–315)[10–758] |
|
| Thrombocytopenia (platelet count <150 × 103/ | 36/42 (86) | 69/92 (75) | 3/4 (75) | 10/37 (27) | 87/399 (22) |
|
| Severe malaria, | 0 (0) | 1 (0.6) [0–3.3] | 0 (0) | 0 (0) | – | 0.244 |
| Patient Characteristic | North Sumatra—Mursala Island | ||||
|---|---|---|---|---|---|
|
|
|
| Control | ||
| No. enrolled, | 6 (10) | 1 (2) | 1 (2) | 55 (86) | – |
| Age, years | |||||
| Median | 46 | 34 | 26 | 25 |
|
| IQR | 30–60 | – | – | 14–40 | |
| Range | 23–72 | – | – | 1–80 | |
| Children (age younger than 15 years old), | 0 (0) | 0 (0) | 0 (0) | 14 (25) | 0.321 |
| Male gender, | 3 (50) | 0 (0) | 0 (0) | 24 (44) | 0.999 |
| Previous malaria (self-reported), | 0 | 0 | 0 | 16 (29) | 0.326 |
| Symptoms on enrolment, | |||||
| Any symptom | 6 (100) | 1 (100) | 1 (100) | 50 (91) | 0.999 |
| Fever history (within 48 hours) | 2 (30) | 1 (100) | 0 | 31 (9) | |
| Headache | 6 (100) | 1 (100) | 0 | 42 (12) | |
| Nausea | 3 (50) | 0 | 1 (100) | 21 (6) | |
| Cough | 6 (100) | 0 | 0 | 35 (10) | |
| Shortness of breath | 4 (75) | 1 (100) | 1 (100) | 17 (5) | |
| Vomiting | 0 (0) | 0 | 0 | 15 (4) | |
| Abdominal pain | 5 (83) | 1 (100) | 0 | 31 (9) | |
| Fever (temperature >37.5°C) | 0 | 0 | 0 | 0 | – |
| Febrile days (before presentation), | 4.5 (2–7) [2–7] | 14 | – | 7 (2–7) [1–30] | 0.670 |
| Microscopy result—positive | 0 (0) | 0 (0) | 0 (0) | 0 (0) | – |
| RDT result (PAN-pLDH/Pf-HRP2) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | – |
| Anemia | 3/3 (100) | 0/1 (0) | 1/1 (100) | 15/41 (37) | 0.062 |
- —Australian Centre for International Agricultural Research and the Department of Foreign Affairs and Trade, Australian Government
- —National Health and Medical Research Council, Australia
- —Australian Centre of Research Excellence in Malaria Elimination
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Vector-borne infectious diseases · Bird parasitology and diseases
INTRODUCTION
Zoonotic malaria caused by Plasmodium knowlesi has emerged as a significant public health challenge in Southeast Asia.1^,^2 Indonesia has a sizeable population at risk of zoonotic malaria3^,^4 because of its ecological diversity and widespread overlap between the parasite’s major natural macaque hosts (Macaca fascicularis and Macaca nemestrina) and Anopheles leucosphyrus group vectors.5 Rapid agricultural expansion and land use changes have amplified the risk of zoonotic malaria by increasing interaction between humans, macaques, and mosquito vectors.6^,^7 The risk is particularly high among adults engaged in forest and agricultural activities.8^,^9 However, understanding of P. knowlesi transmission in Indonesia is limited because of being frequently misdiagnosed as nonzoonotic species using routine microscopy, with Plasmodium falciparum and Plasmodium vivax often incorrectly identified even more so than the morphologically similar Plasmodium malariae.10^,^11 Lateral flow-based rapid diagnostic tests (RDTs), designed and commonly used for detection of P. falciparum and P. vivax, have also lacked the specificity and until recently, the sensitivity required for reliable P. knowlesi identification, particularly at low parasite densities.12?^–^14 Diagnostic misclassification using available point-of-care diagnostics has previously obscured the actual burden of P. knowlesi malaria, a trend first observed in Sarawak, Malaysian Borneo, where a rise in cases of microscopy-diagnosed P. malariae ultimately led to the initial molecular identification of P. knowlesi as a dominant cause of human malaria in 2004.15
The use of accurate polymerase chain reaction (PCR) assays in research settings has highlighted the presence of P. knowlesi transmission across western Indonesia, particularly within the provinces of Sumatra10^,^11^,^16????^–^21 and Kalimantan,22???^–^26 but outside the nonendemic eastern province of Papua, where the highest burden of nonzoonotic malaria remains.27 However, there remain significant gaps in our understanding of the distribution of P. knowlesi within areas such as North Sumatra, where diverse ecological conditions are conducive to zoonotic malaria transmission3 but where many districts have not been systematically evaluated using sensitive molecular tools. Plasmodium knowlesi transmission has been reported to date in the districts of Batubara, Langkat, and Dairi and the islands of South Nias.11^,^20^,^28 Other zoonotic malaria species, such as Plasmodium cynomolgi, have not been screened for in western Indonesia, despite sharing the same macaque hosts and mosquito vectors.29 Understanding the prevalence and distribution of zoonotic Plasmodium species in regions like North Sumatra is critical for improving malaria control efforts, meeting WHO malaria elimination goals, and addressing the hidden burden of zoonotic malaria in Indonesia.30 The aim of this study was to conduct molecular epidemiological surveillance using improved ultrasensitive PCR for both zoonotic P. knowlesi and P. cynomolgi detection in high-risk areas across North Sumatra.
MATERIALS AND METHODS
Study design.
The main study design consisted of longitudinal passive malaria case detection of febrile patient presentations at selected health facilities on mainland North Sumatra, Indonesia. A separate small cross-sectional study was conducted on Mursala Island.
Study sites.
The health facility surveys on the North Sumatra mainland covered a study area of 6,933 kilometers squared (km^2^), with an estimated 1,579,000 people at risk31 (Figure 1). There were eight health facilities from across three districts, including four government primary health clinics in Batubara, two primary health clinics and a referral hospital in Central Tapanuli, and an urban referral hospital in Tanjung Balai. The estimated prevalence of malaria was taken into account when selecting the health facility locations. At the time of the study design in 2019, provincial reports indicated that Batubara accounted for 41% of malaria cases in North Sumatra, with an annual parasite index (API) of 1.1 positive case per 1,000 people tested; Asahan accounted for 10% prevalence, with an API of 0.16, and Central Tapanuli accounted for 2% prevalence, with an API of 0.06.1^,^3
Map of study sites: Batubara district (blue), Tanjung Balai district (green), and Central Tapanuli (red).
A separate cross-sectional study was performed on Mursala Island, which is also part of Central Tapanuli district; it is located 22 km off the coast and inhabited by 5,400 people, and it has a size of 80 km^2^. Community approval and awareness for the survey were obtained after meeting with village leaders and heads of households.
Study participants.
Patients at mainland study health facilities were enrolled if they met the following inclusion criteria: age of 1 year old or older, body temperature above 37.5°C or a history of fever in the last 48 hours, presented with nonspecific symptoms (suspected malaria), resided primarily in the study district within the previous 1 year, and provided informed written consent (from parents/guardians if younger than 18 years old). For Mursala Island, criteria were similar, although a history of fever was not required. Demographic, clinical, and environmental data and malaria prevention methods and activities, including macaque interaction data, were collected by trained research staff using a standardized electronic case record form (REDCap v. 10.6, Nashville, TN). Severe malaria was defined using WHO 2014 Research and Epidemiological criteria for severe disease.32
Blood sampling.
Venous whole blood (or a capillary sample in those 10 years old or younger and all participants from Mursala) was collected in ethylenediaminetetraacetic acid for standard hematology and biochemistry at local laboratories where available. Hemoglobin was measured with a HemoCue^®^ 201 machine (HemoCue AB, Ängelholm, Sweden) on Mursala Island. Malaria screening was performed using Ministry of Health-supplied routine malaria RDTs with combined pan-Plasmodium species parasite lactate dehydrogenase (PAN-pLDH) and P. falciparum-specific histidine-rich protein-2 (Pf-HRP2) targets depending on site availability and microscopic examination of blood smears stained with Giemsa. Slides and RDTs were read by local health facility officers, and slides were re-examined and quantified by an experienced microscopist blinded to the first microscopy and RDT results according to standard WHO procedures.33 A total of 300 and 100 µL of whole blood from adults and children, respectively, were placed in field-stable DNA/RNA Shield^TM^ (Zymo Research, Irvine, CA) total nucleic acid preservation media and frozen at 4°C before shipping to the Faculty of Medicine, Universitas Sumatera. Patients confirmed as malaria positive on point-of-care testing were treated according to national guidelines.34
Laboratory malaria PCR detection.
Total nucleic acids were extracted (QIAamp DNA Blood Mini Kit, Hilden, Germany) from 200 µL of the blood sample preserved in DNA/RNA Shield followed by high-capacity complementary DNA (cDNA) reverse transcription (Applied Biosystems, Waltham, MA). Amplification of cDNA was conducted using a quantitative real-time polymerase chain reaction (qRT-PCR) workflow targeting 18S ribosomal RNA genes.35 Positive Plasmodium genus results were defined as cycle threshold (Ct) values below 40 from duplicate runs, with a Ct value difference of less than three. Those positive had conventional species-specific PCR assays performed using the same cDNA for P. knowlesi,36 P. cynomolgi,37 and other human Plasmodium species (P. falciparum, P. vivax, Plasmodium ovale spp., and P. malariae38) detection. Each PCR amplification included Plasmodium species positive and negative controls and molecular weight standards.
STATISTICAL ANALYSES
All statistical analyses were performed using Stata v. 17.0 (StataCorp, College Station, TX). The χ^2^ or Fisher exact tests were used to evaluate proportional differences in binary variables, and the Student’s t-tests or Wilcoxon rank sum tests were used for pair-wise comparisons of clinical and epidemiological data. Results of microscopy and RDT assays were evaluated against reference PCR, enabling calculation of diagnostic sensitivity and specificity with exact binomial 95% CIs. The crude malaria incidence rate was calculated as the number of malaria cases per 100,000 at-risk people per year at a district catchment level. Study site district catchment populations were projected estimates from 2020 census data.39 Logistic regression models were used for univariable and multivariable epidemiological associations with individual Plasmodium species infections, with odds ratios (ORs) and 95% CIs reported.
RESULTS
Mainland health facility surveys.
From August 2019 to December 2020, there were 3,377 patients with acute febrile illness presenting to the health facilities in mainland North Sumatra (Figure 2). Of these, 947 (28.1%) were enrolled, including 704 (74.3%) patients from Batubara, 136 (14.4%) patients from Tanjung Balai, and 107 (11.3%) patients from Central Tapanuli. The median age was 17 years old (interquartile range [IQR]: 12–34; range: 2–67), and 48% were female.
Enrollment flowchart for health facility surveys and Plasmodium (P.) species results. Aug = August; Nov = November; Pcyn = Plasmodium cynomolgi; Pf = Plasmodium falciparum; Pk = Plasmodium knowlesi; Pm = Plasmodium malariae; Po = Plasmodium ovale; Pv = Plasmodium vivax; qPCR = quantitative polymerase chain reaction.
Overall, 321 (33.9%; 95% CI: 30.9–37.0%) patients tested positive for malaria by PCR; 71 (7.5%) patients tested positive for P. falciparum, 166 (17.5%) patients tested positive for P. vivax, and 5 (0.5%) patients tested positive for mixed P. falciparum/P. vivax. In 79 (8.3%) patients, we were unable to determine the exact Plasmodium species infection. There were 96 febrile patients with submicroscopic infections (29.9% of total infections; 95% CI: 24.9–35.2%), including 8 patients with P. falciparum, 16 patients with P. vivax, and 72 patients with Plasmodium genus.
There were no significant differences in the median age (approximately 16 years old) or the proportion of children (approximately 35%) with P. falciparum or P. vivax infections (Table 1). The majority presented with nonspecific symptoms, most commonly headache, nausea/vomiting, cough, and abdominal pain, with a median preceding fever duration of 6 days. Anemia (using WHO age and sex hemoglobin criteria40) was observed in more than 40% of patients infected with P. falciparum, P. vivax, or undetermined Plasmodium species. Thrombocytopenia (platelet count <150 × 10^3^/µL) was present in more than 75% of patients with P. falciparum or P. vivax malaria. In contrast, anemia and thrombocytopenia were only present in 30% and 22% of the febrile nonmalaria controls, respectively (P ≤0.008). A previous history of malaria was positively associated with acute infections (OR: 2.91; 95% CI: 1.88–4.50; P <0.001).
There was a single patient meeting 2014 research and epidemiological criteria for WHO-defined severe disease (0.4% of malaria cases with an identified Plasmodium species; 95% CI: 0.01–2.3%): an adult male with vivax malaria, a hemoglobin of 6.3 g/dL, and a parasite count of 3,120/µL.
On routine microscopy, 249 positive results for malaria were reported, including 12% with P. vivax, 6% with P. falciparum, and a single P. knowlesi/P. malariae result. These findings resulted in an overall sensitivity for Plasmodium species detection against reference PCR of 68.8% (95% CI: 63.5–73.9%), with a specificity of 95.5% (95% CI: 93.6–97%) against malaria-negative controls. Plasmodium falciparum single infections were correctly identified in 42 (59.2%) of patients, with a further 21 cases (29.6%) misidentified as mixed infections. Similarly, single P. vivax infections were correctly identified in 60% of participants; however, they were misidentified as mixed P. vivax infections in another 27%. In contrast, there were only small numbers of false-positive microscopy results for P. falciparum and P. vivax single infections, leading to high diagnostic specificities of 98.4% (95% CI: 97.3–99.1%) and 98.1% (95% CI: 96.9–98.9%) for each species, respectively. The geometric mean parasite count for P. falciparum infections was low at 2,844/µL (95% CI: 2,055–3,936/µL), similar to P. vivax at 2,273/µL (95% CI: 2,487–3,187/µL).
The performance of the combined PAN-pLDH/Pf-HRP2 RDT was poor compared with reference PCR, with 155 (22.5%) positive PAN-pLDH results and 79 (11.4%) positive Pf-HRP2 results recorded. These findings resulted in sensitivity and specificity of 82.4% (95% CI: 69.1–91.6%) and 78.5% (95% CI: 75.1–81.6%), respectively, for P. falciparum detection and sensitivity and specificity of 81.3% (95% CI: 73.3–87.8%) and 86.0% (95% CI: 82.9–88.8%), respectively, for P. vivax. Sensitivity of the RDT compared with microscopy against PCR as the reference was higher overall for detection of P. falciparum (82% versus 53%; P = 0.0015) and P. vivax (81% versus 65%; P = 0.0065) single infections.
Student was the most frequently reported primary occupation among malaria cases, accounting for approximately 50% overall (Supplemental Table 1), which reflects the predominantly adolescent cohort. On univariable analysis, a history of sleeping outdoors was associated with higher malaria risk, whereas insecticide-treated bed net use and personal insect repellent were protective. Housing type also played a key role; concrete walls were protective, whereas open eaves increased risk. Proximity to mangroves or cleared forest areas was also associated with higher odds of infection. Factors that remained independently associated with malaria in the multivariable analysis are summarized in Figure 3.
Forest plot of adjusted odds ratios for epidemiological associations with malaria acquisition risk.
Mursala Island cross-sectional survey.
In a small cross-sectional survey conducted on Mursala Island in September 2019, 64 individuals were enrolled (Table 2). The median age was 27 years old (IQR: 15.5–40; range: 1–80), and 58% were female. Most participants (92%) were symptomatic, including 35% reporting a history of fever. Polymerase chain reaction detected Plasmodium spp. infections in nine individuals (14.1%; 95% CI: 6.6–25%), comprising six with P. knowlesi, one with P. vivax, one with mixed P. knowlesi/P. vivax, and one Plasmodium genus-positive case with undetermined species. No infections were detected in patients younger than 18 years of age. None of the Plasmodium species-infected individuals reported a prior malaria diagnosis.
Overall, seven patients (10.9%; 95% CI: 4.7–21.9%) were infected with P. knowlesi, with a higher median age (36 years old; IQR: 26–60) compared with malaria-negative or non-P. knowlesi cases (17 years old; P = 0.037). Females accounted for 57% of P. knowlesi cases. Common symptoms among P. knowlesi cases included headache, cough, and abdominal pain, although only 29% reported fever in the preceding 48 hours. The median reported duration of fever was 4.5 days (IQR: 2–7) for P. knowlesi and 14 days for P. vivax. Anemia, defined by WHO age- and sex-specific thresholds,40 was present in all P. knowlesi-infected individuals tested. Two adults with P. knowlesi infections met WHO criteria for severe malaria,32 with hemoglobin levels of 4.4 and 5.1 g/dL, respectively. Compared with nonmalaria controls, P. knowlesi cases had significantly lower median hemoglobin levels (7.1 versus 13 g/dL; P = 0.004). No other severe malaria features were observed. However, further assessment of respiratory distress or organ dysfunction (e.g., acute kidney injury, hyperbilirubinemia, hypoglycemia, and metabolic acidosis) was limited by the lack of appropriate clinical and laboratory facilities, respectively. None of the P. knowlesi or P. vivax infections were detected by microscopy or the combined PAN-pLDH/Pf-HRP2 RDT.
In Mursala, although the small survey size limits meaningful comparisons, five of the seven individuals infected with P. knowlesi had occupations related to agriculture (four farmers and one plantation worker) (Supplemental Table 2). Recent forest exposure (>4 hours) and proximity to rubber plantations were more common among P. knowlesi cases (86% each) than controls (46% and 43%, respectively; P = 0.076 for both comparisons). However, forest activities related to wood collection were associated with higher risk of P. knowlesi acquisition (OR: 16; 95% CI: 1.8–144; P = 0.013). Bed net use the night before enrollment was common in both groups (86% of P. knowlesi cases versus 64% of controls). Only one P. knowlesi-infected case reported sleeping outside in the previous 2 weeks. Regular awareness of monkey presence was high among cases (86%), although not significantly different from controls. No household structural or environmental factors, including wall type, elevation, or open eaves, were significantly associated with infection.
DISCUSSION
This study aimed to further characterize the distribution of zoonotic P. knowlesi infections in North Sumatra, Indonesia using accurate molecular tools. Key findings include the presence of previously unreported P. knowlesi submicroscopic symptomatic infections on the remote rural island of Mursala. Additionally, results highlight the significant ongoing burden of nonzoonotic malaria (34%) detected by PCR among acute febrile illness presentations in densely populated mainland coastal areas. North Sumatra public health reporting estimated that P. falciparum caused 15% of clinical malaria and that P. vivax caused 85% of clinical malaria in 2019 (Provincial Health Office 2019, unpublished data). In our study, the predominance of P. vivax infections underscores the ongoing contribution of this species to the malaria burden despite national elimination efforts, with a doubling of malaria cases reported in North Sumatra since the coronavirus disease 2019 (COVID-19) pandemic in 2020.27 Although Mursala Island had a lower overall malaria prevalence, submicroscopic infections were predominantly caused by zoonotic P. knowlesi transmission. No other zoonotic Plasmodium species, such as P. cynomolgi, were identified.
Although P. knowlesi infections have been previously reported in North Sumatra,11^,^20 none were detected during this study among participants from densely populated coastal mainland sites located away from higher-risk agricultural or forest areas. Modeled transmission suitability for P. knowlesi in the coastal districts of Batubara and Central Tapanuli is relatively low, with median values of 0.12 (IQR: 0.05–0.38) and 0.35 (IQR: 0.13–0.55), respectively, compared with 0.61 (IQR: 0.27–0.82) in the more interior region of South Tapanuli (R. J. Tobin, L. E. Harrison, M. K. Tully, I. N. D. Lubis, R. Noviyanti, N. M. Anstey, G. S. Rajahram, M. J. Grigg, J. A. Flegg, D. J. Price, and F. M. Shearer, unpublished data).3 This finding likely reflects distinct epidemiological patterns, similar to those in Malaysian Borneo, where heterogeneous P. knowlesi transmission is concentrated in rural areas undergoing land use change.3^,^41^,^42 Unlike Malaysia, however, there is limited understanding about the distribution and behavior of suspected An. leucosphyrus group mosquito vectors across Sumatra43^,^44 or the variation in P. knowlesi infection prevalence in local macaque populations,21^,^44^,^45 both of which are key determinants of spatial zoonotic transmission patterns.3^,^5^,^46 However, the demographic profile of P. knowlesi-infected individuals—exclusively adults—suggests higher exposure among those engaged in agricultural activities where macaque hosts are present. Both males and females had P. knowlesi infections detected, with similar self-reported exposure patterns aligning with findings from other areas of previously reported zoonotic malaria transmission in Malaysia9 and elsewhere in western Indonesia.16 In contrast, P. falciparum and P. vivax infections in this study predominantly affected younger adolescents, consistent with well-established demographic patterns in other endemic regions approaching elimination of these nonzoonotic species.47
The majority of zoonotic and nonzoonotic malaria cases presented with nonspecific clinical symptoms, highlighting the ongoing disease burden of malaria and critical need for vigilant point-of-care diagnostic testing. Laboratory findings of moderate anemia and thrombocytopenia were more common in malaria-positive patients, which may aid clinical suspicion for diagnostic screening. The small number of patients with P. knowlesi infections from Mursala all exhibited anemia, despite extremely low-level submicroscopic parasitemia, including two cases that met WHO criteria for severe malarial anemia (hemoglobin <7g/dL).32 Although chronic causes of anemia cannot be excluded—particularly given the high baseline prevalence in malaria-negative individuals—P. knowlesi-associated anemia may also result from splenic retention of less-deformable uninfected erythrocytes and/or bystander destruction of uninfected erythrocytes or dyserythropoiesis,48^,^49 potentially exacerbating underlying vulnerability. Evidence from Malaysia similarly reports anemia in uncomplicated P. knowlesi infections, although generally less prevalent and severe than in P. falciparum and P. vivax malaria.50^,^51 The low parasitemia levels in Mursala cases could reflect partial immunity from repeated P. knowlesi exposure, crossprotection from prior P. vivax infections, or reduced erythrocyte invasion because of parasite or host genetic factors, including hemoglobinopathies.52 Parasite counts of >15,000/µL have been linked to a >16-fold risk of severe P. knowlesi malaria in Malaysian Borneo, often with clinical complications, such as jaundice, respiratory distress, hypotension, and acute kidney injury.50^,^51 These manifestations were not observed either because of the low parasite counts or because of the limited laboratory testing available in this study.
The threat of emerging P. knowlesi transmission highlights the ongoing importance of access to robust diagnostic capabilities and antimalarial treatment in provinces approaching elimination of nonzoonotic Plasmodium species. Indonesia’s national malaria control program currently relies on passive surveillance using microscopy or PAN-pLDH/Pf-HRP2-based RDTs.53 In our study, microscopy demonstrated only moderate sensitivity for detecting P. falciparum (59.2%) and P. vivax (62.0%) monoinfections, with notable difficulty distinguishing mixed infections. This raises concerns around misclassification of P. vivax, which may result in a lack of primaquine administration for radical liver cure. The PAN-pLDH/Pf-HRP2 RDT showed strong performance in detecting P. falciparum and P. vivax, reinforcing its utility in field settings. However, our molecular diagnostics identified a high proportion of submicroscopic (30%) and Plasmodium genus of undetermined species (25%) in mainland sites, likely because of the enhanced sensitivity of the reverse transcriptase real-time PCR workflow with a lower limit of detection in the Plasmodium genus compared with species-specific assays.35 However, we cannot exclude the possibility of false-positive results arising from sample inhibitors, degraded RNA, or other assay-related factors. The qRT-PCR assay applied in this study has been shown to detect both zoonotic and nonzoonotic malaria at up to 10,000-fold-higher threshold limits, providing valuable insights into low-level zoonotic malaria transmission dynamics and supporting malaria elimination efforts in Southeast Asia. Integrating a reverse transcription step into conventional PCR assays has also been demonstrated to significantly improve sensitivity and may represent a more robust approach for complementary molecular surveillance, including in asymptomatic populations at risk.35 In contrast, conventional point-of-care tests failed to detect the low-density P. knowlesi infections found in patients on Mursala Island, despite their symptomatic presentation. This aligns with previous studies showing poor performance of PAN-pLDH-based RDT for P. knowlesi,13^,^14 although the sensitivity of more recent RDTs has improved for parasite counts greater than 200/µL.12 These limitations emphasize the critical need to integrate molecular tools into surveillance systems in areas approaching malaria elimination, especially where P. knowlesi is suspected.30
Plasmodium knowlesi infection has been associated with environmental and occupational risk factors, including forest and agricultural work (particularly among farmers and plantation workers), sleeping outside, and household characteristics such as open eaves or gaps in walls.9 In our study, although the majority of P. knowlesi cases (71%) were engaged in agricultural activities, this occupation alone did not significantly increase infection risk. Recent forest exposure, house proximity to rubber plantations, and wood collection in forested areas were commonly identified for the small number of P. knowlesi infections in Mursala Island, although we were underpowered to determine if these were also statistically significant associations in this setting. It is likely that these trends indicate a zoonotic transmission cycle involving forest-dwelling vectors and macaque reservoirs, contrasting with the coastal semiurban environments of our mainland study sites where P. knowlesi infections were not detected. Furthermore, although malaria prevention measures, such as bed net use and personal insect repellent use, may continue to provide benefits for preventing nonzoonotic malaria, their use did not provide significant protection against P. knowlesi infection in our study. This finding emphasizes the need for tailored prevention strategies targeting high-risk groups engaged in forest-related activities to effectively mitigate zoonotic malaria transmission risk.
Our study has several limitations. In the mainland study sites, there was an overrepresentation of young individuals, with only 14% of participants being adult men older than 30 years old; additionally, only 3% of all participants reported either agricultural occupations or recent forest exposure. This might have introduced a degree of selection bias by insufficiently capturing high-risk populations, particularly adult forest and agricultural workers who are most vulnerable to zoonotic malaria.9 The study's design focus on symptomatic cases presenting to health facilities potentially underestimates the true infection burden by missing mild and asymptomatic cases that did not seek medical attention.54 Additionally, the COVID-19 pandemic interrupted both the study timeline and routine malaria activities, potentially affecting health care-seeking behavior and contributing to increased malaria burden during the study period. On Mursala Island, the cross-sectional nature of the surveys and the relatively small number of P. knowlesi cases limit our ability to draw definitive conclusions about risk factors and transmission patterns. Furthermore, the lack of laboratory facilities for comprehensive assessments of WHO severity criteria prevented thorough evaluation of key parameters of severe malaria, particularly in P. knowlesi cases.
CONCLUSION
Our study highlights the complex malaria epidemiology in North Sumatra and the ongoing challenges to Indonesia’s elimination goals. Despite control efforts, nonzoonotic malaria—particularly P. vivax—remains a major cause of febrile illness on the mainland. In contrast, P. knowlesi transmission on Mursala Island reflects distinct zoonotic dynamics linked to forest-related activities, such as rubber plantation proximity, wood collection, and recent forest travel. These findings underscore the need to strengthen molecular diagnostic capacity and adopt improved sampling strategies in future surveillance to better understand and mitigate the growing threat of zoonotic malaria in Indonesia.
Supplemental Materials
10.4269/ajtmh.25-0493Supplemental Materials
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization, 2024. World Malaria Report 2024. Available at: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2024. Accessed November 7, 2025.
- 2Cox-Singh J Singh B, 2008. Knowlesi malaria: Newly emergent and of public health importance? Trends Parasitol 24: 406–410.18678527 10.1016/j.pt.2008.06.001PMC 2843823 · doi ↗ · pubmed ↗
- 3Tobin RJ, , 2024. Updating estimates of Plasmodium knowlesi malaria risk in response to changing land use patterns across Southeast Asia. P Lo S Negl Trop Dis 18: e 0011570.38252650 10.1371/journal.pntd.0011570 PMC 10833542 · doi ↗ · pubmed ↗
- 4Lempang MEP Dewayanti FK Syahrani L Permana DH Malaka R Asih PBS Syafruddin D, 2022. Primate malaria: An emerging challenge of zoonotic malaria in Indonesia. One Health 14: 100389.35686151 10.1016/j.onehlt.2022.100389 PMC 9171520 · doi ↗ · pubmed ↗
- 5Moyes CL, , 2016. Predicting the geographical distributions of the macaque hosts and mosquito vectors of Plasmodium knowlesi malaria in forested and non-forested areas. Parasit Vectors 9: 242.27125995 10.1186/s 13071-016-1527-0PMC 4850754 · doi ↗ · pubmed ↗
- 6Fornace KM, , 2019. Local human movement patterns and land use impact exposure to zoonotic malaria in Malaysian Borneo. e Life 8: e 47602.31638575 10.7554/e Life.47602 PMC 6814363 · doi ↗ · pubmed ↗
- 7Brock PM Fornace KM Grigg MJ Anstey NM William T Cox J Drakeley CJ Ferguson HM Kao RR, 2019. Predictive analysis across spatial scales links zoonotic malaria to deforestation. Proc Biol Sci 286: 20182351.30963872 10.1098/rspb.2018.2351 PMC 6367187 · doi ↗ · pubmed ↗
- 8Fornace KM, , 2018. Exposure and infection to Plasmodium knowlesi in case study communities in Northern Sabah, Malaysia and Palawan, The Philippines. Plos Negl Trop Dis 12: e 0006432.29902171 10.1371/journal.pntd.0006432 PMC 6001952 · doi ↗ · pubmed ↗